
Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target

 ,
,  ,
,  ,
, 

Abstract
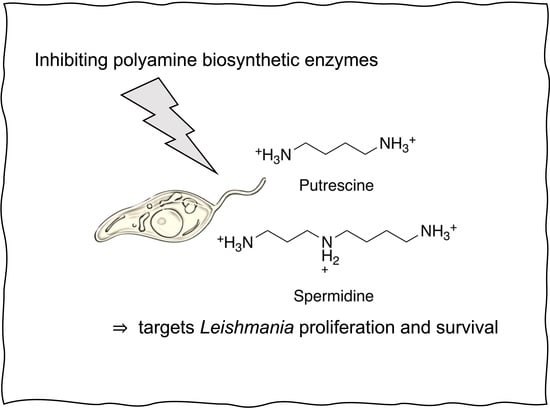
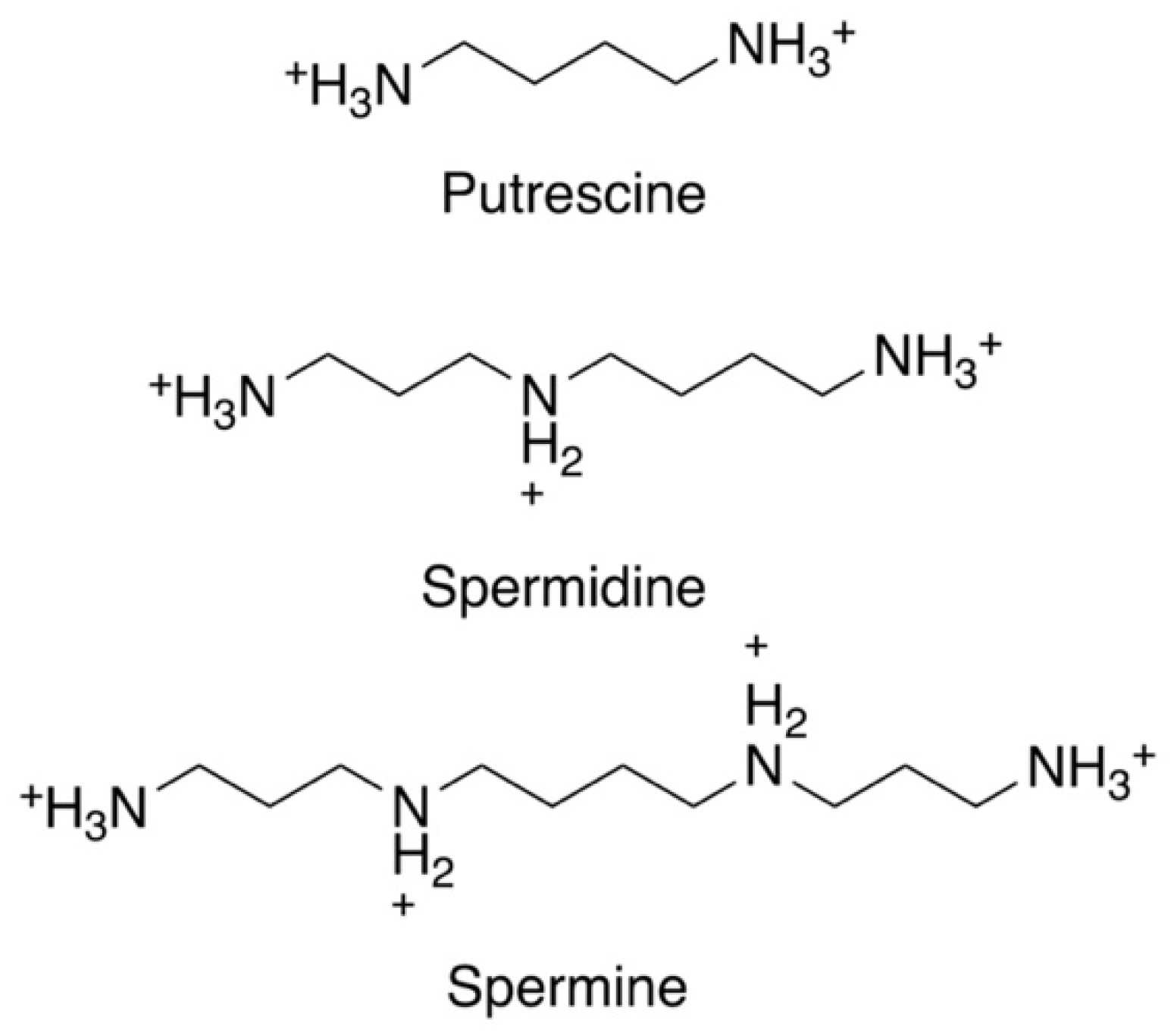
:
1. Introduction to Leishmania
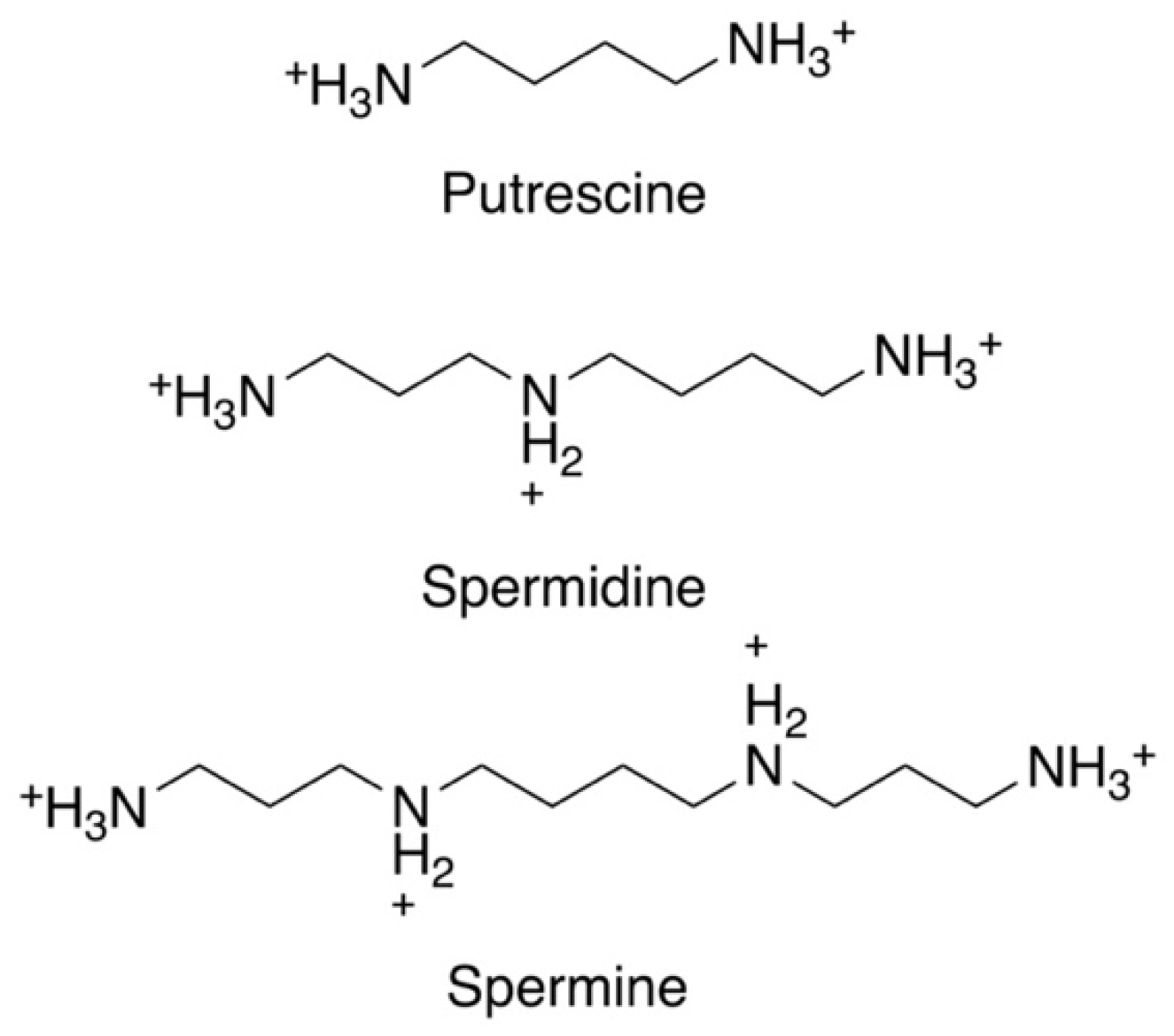
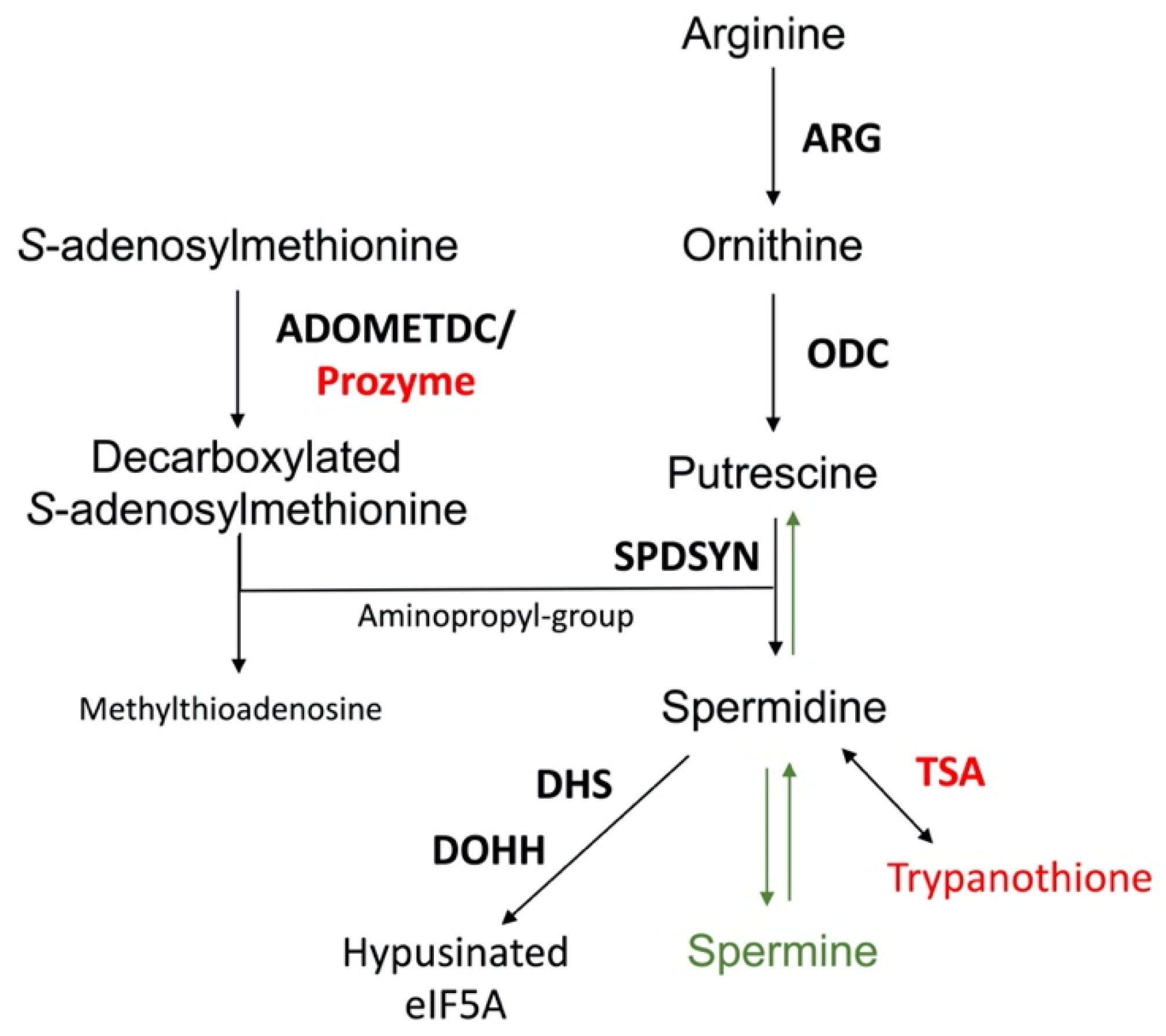
2. Significance of Polyamines
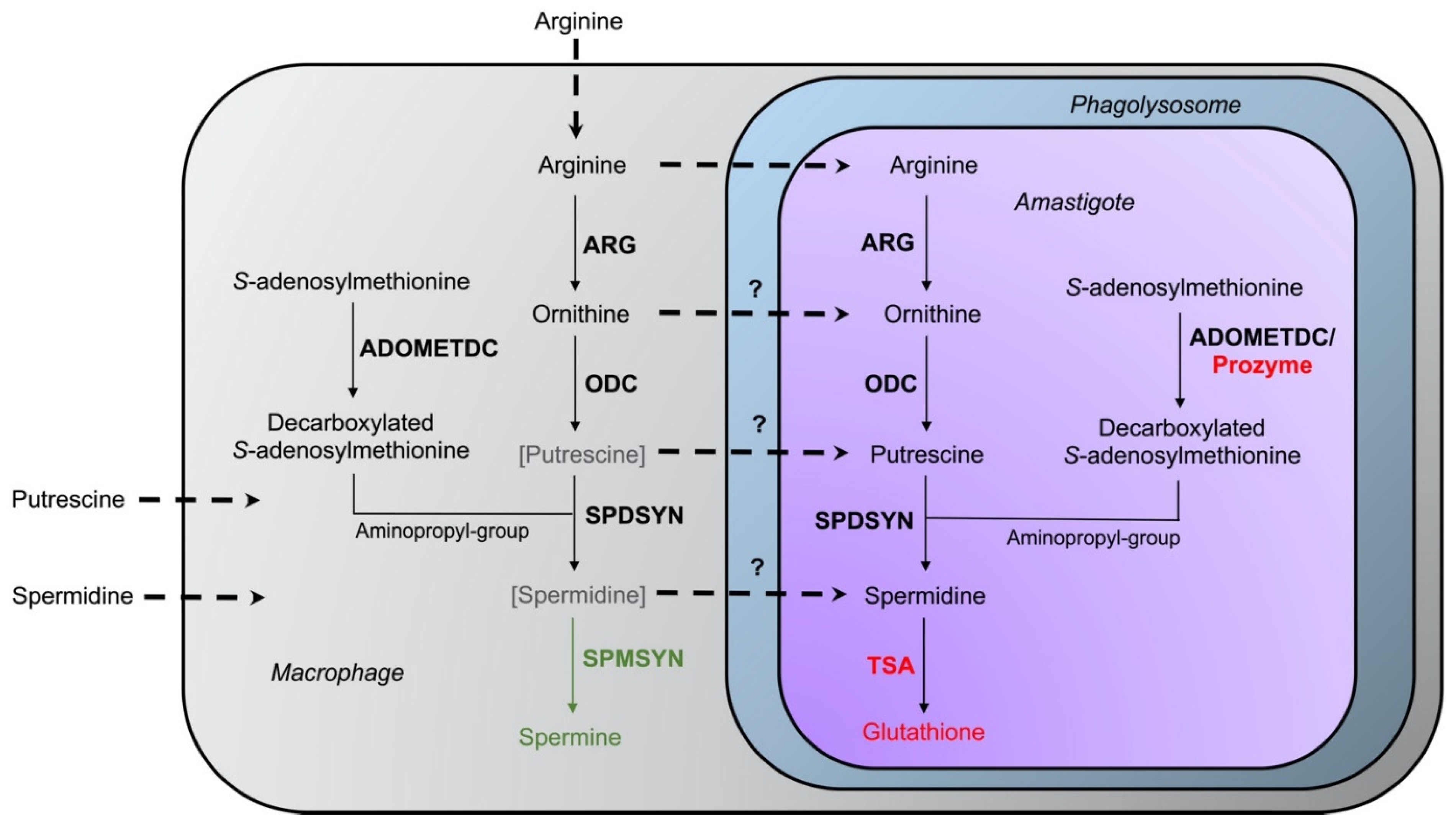
3. Relevance of Polyamines for Host Parasite Interactions
4. Ornithine Decarboxylase (ODC)
4.1. Enzyme Structure and Function
4.2. ODC Gene Deletion Studies
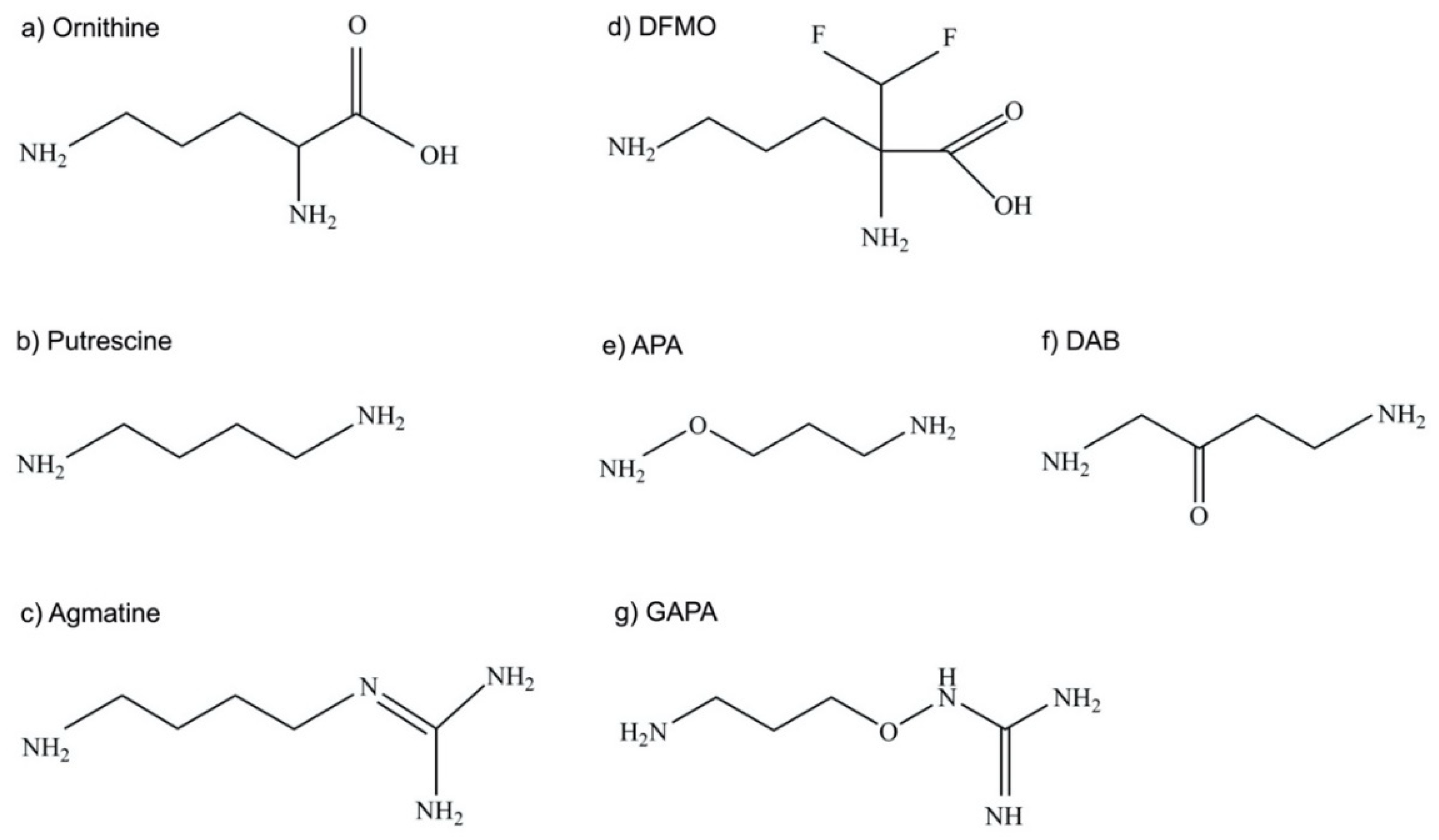
4.3. Efficacy of the Ornithine Analog DFMO as ODC Inhibitor
4.4. Putrescine and Agmatine Analogs as ODC Inhibitors
4.5. Spermine Analogs as ODC Inhibitors
4.6. Other Inhibitors of ODC
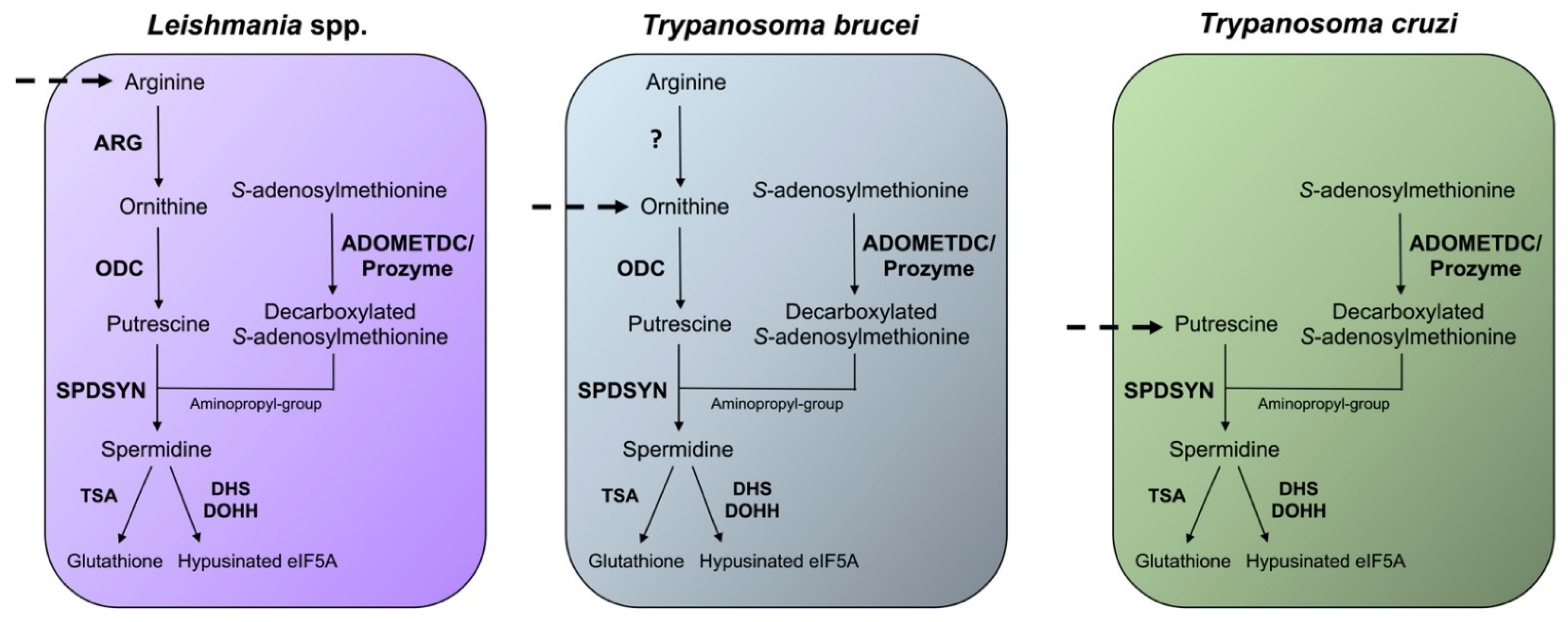
5. S-adenosylmethionine Decarboxylase (ADOMETDC)
5.1. Enzyme Structure and Function
5.2. ADOMETDC Gene Deletion Studies
5.3. Inhibitors of the Leishmanial ADOMETDC
6. Spermidine Synthase (SPDSYN)
6.1. Enzyme Structure and Function
6.2. SPDSYN Gene Deletion Studies
6.3. Inhibitors of SPDSYN
7. Polyamine Analogs
8. Polyamine Transport
8.1. Arginine Transport
8.2. Ornithine Transport
8.3. Putrescine Transport
8.4. Spermidine Transport
8.5. S-adenosylmethionine Transport
9. Drug Resistance and Polyamines
10. Discussion
11. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADOMETDC | S-adenosylmethionine decarboxylase |
| APA | 3-aminooxy-1-aminopropane |
| BFPT | N-[1] benzofuro [3, 2-d] pyrimidin-4-yl-d-tryptophane |
| DAB | 1,4-diamino-2-butanone |
| DFMO | D, L-α-difluoromethylornithine |
| DHC | dihydrocitrinone |
| GAPA | gamma-guanidinooxy propylamine or 1-guanidinooxy-3-aminopropane |
| MGBG | methylglyoxal bis(guanylhydrazone) |
| ODC | ornithine decarboxylase |
| SPDSYN | spermidine synthase |
| SPMSYN | spermine synthase |
| TSA | trypanothione synthetase/amidase |
| ARG | arginase |
References
- World Health Organization. Leishmaniasis; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Grifferty, G.; Shirley, H.; McGloin, J.; Kahn, J.; Orriols, A.; Wamai, R. Vulnerabilities to and the Socioeconomic and Psychosocial Impacts of the Leishmaniases: A Review. Res. Rep. Trop. Med. 2021, 12, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Yactayo, S.; Bern, C. Leishmaniasis and poverty. Trends Parasitol. 2006, 22, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J. The rise of leishmaniasis in the twenty-first century. Trans. R. Soc. Trop. Med. Hyg. 2018, 112, 421–422. [Google Scholar] [CrossRef] [Green Version]
- Bogdan, C. Macrophages as host, effector and immunoregulatory cells in leishmaniasis: Impact of tissue micro-environment and metabolism. Cytokine X 2020, 2, 100041. [Google Scholar] [CrossRef]
- Meira, C.D.S.; Gedamu, L. Protective or Detrimental? Understanding the Role of Host Immunity in Leishmaniasis. Microorganisms 2019, 7, 695. [Google Scholar] [CrossRef] [Green Version]
- Kupani, M.; Pandey, R.K.; Mehrotra, S. Neutrophils and Visceral Leishmaniasis: Impact on innate immune response and cross-talks with macrophages and dendritic cells. J. Cell. Physiol. 2020, 236, 2255–2267. [Google Scholar] [CrossRef]
- Tsigankov, P.; Gherardini, P.F.; Helmer-Citterich, M.; Späth, G.F.; Zilberstein, D. Phosphoproteomic Analysis of Differentiating Leishmania Parasites Reveals a Unique Stage-Specific Phosphorylation Motif. J. Proteome Res. 2013, 12, 3405–3412. [Google Scholar] [CrossRef]
- Zilberstein, D. Lysosome Sensing Is a Key Mechanism in Leishmania Intracellular Development. Front. Microbiol. 2021, 12, 864. [Google Scholar] [CrossRef]
- Zilberstein, D.; Shapira, M. The role of pH and temperature in the development of Leishmania parasites. Annu. Rev. Microbiol. 1994, 48, 449–470. [Google Scholar] [CrossRef]
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martinez, A.F.; Newman, S.; Ramanan, P.; Suarez, J.A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Curr. Trop. Med. Rep. 2021, 8, 121–132. [Google Scholar] [CrossRef]
- Sasidharan, S.; Saudagar, P. Leishmaniasis: Where are we and where are we heading? Parasitol. Res. 2021, 120, 1541–1554. [Google Scholar] [CrossRef]
- Velez, R.; Gállego, M. Commercially approved vaccines for canine leishmaniosis: A review of available data on their safety and efficacy. Trop. Med. Int. Health 2020, 25, 540–557. [Google Scholar] [CrossRef] [Green Version]
- Kumari, D.; Perveen, S.; Sharma, R.; Singh, K. Advancement in leishmaniasis diagnosis and therapeutics: An update. Eur. J. Pharmacol. 2021, 910, 174436. [Google Scholar] [CrossRef]
- Berman, J. Amphotericin B Formulations and Other Drugs for Visceral Leishmaniasis. Am. J. Trop. Med. Hyg. 2015, 92, 471–473. [Google Scholar] [CrossRef] [Green Version]
- Rakotomanga, M.; Blanc, S.; Gaudin, K.; Chaminade, P.; Loiseau, P.M. Miltefosine Affects Lipid Metabolism in Leishmania donovani Promastigotes. Antimicrob. Agents Chemother. 2007, 51, 1425–1430. [Google Scholar] [CrossRef] [Green Version]
- Dorlo, T.P.C.; Balasegaram, M.; Beijnen, J.H.; De Vries, P.J. Miltefosine: A review of its pharmacology and therapeutic efficacy in the treatment of leishmaniasis. J. Antimicrob. Chemother. 2012, 67, 2576–2597. [Google Scholar] [CrossRef]
- Le Pape, P. Development of new antileishmanial drugs—Current knowledge and future prospects. J. Enzym. Inhib. Med. Chem. 2008, 23, 708–718. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Bogdan, C. Mechanisms and consequences of persistence of intracellular pathogens: Leishmaniasis as an example. Cell. Microbiol. 2008, 10, 1221–1234. [Google Scholar] [CrossRef]
- Conceição-Silva, F.; Leite-Silva, J.; Morgado, F. The Binomial Parasite-Host Immunity in the Healing Process and in Reactivation of Human Tegumentary Leishmaniasis. Front. Microbiol. 2018, 9, 1308. [Google Scholar] [CrossRef]
- Gedda, M.R.; Singh, B.; Kumar, D.; Singh, A.K.; Madhukar, P.; Upadhyay, S.; Singh, O.P.; Sundar, S. Post kala-azar dermal leishmaniasis: A threat to elimination program. PLOS Neglected Trop. Dis. 2020, 14, e0008221. [Google Scholar] [CrossRef]
- Mendonça, M.G.; De Brito, M.E.F.; Rodrigues, E.H.G.; Bandeira, V.; Jardim, M.L.; Abath, F.G.C. Persistence of Leishmania Parasites in Scars after Clinical Cure of American Cutaneous Leishmaniasis: Is There a Sterile Cure? J. Infect. Dis. 2004, 189, 1018–1023. [Google Scholar] [CrossRef] [Green Version]
- Barrett, M.P.; Kyle, D.E.; Sibley, L.D.; Radke, J.B.; Tarleton, R.L. Protozoan persister-like cells and drug treatment failure. Nat. Rev. Genet. 2019, 17, 607–620. [Google Scholar] [CrossRef]
- Elmahallawy, E.K.; Alkhaldi, A.A.; Saleh, A.A. Host immune response against leishmaniasis and parasite persistence strategies: A review and assessment of recent research. Biomed. Pharmacother. 2021, 139, 111671. [Google Scholar] [CrossRef] [PubMed]
- Mandell, M.A.; Beverley, S.M. Continual renewal and replication of persistent Leishmania major parasites in concomitantly immune hosts. Proc. Natl. Acad. Sci. USA 2017, 114, E801–E810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colotti, G.; Ilari, A. Polyamine metabolism in Leishmania: From arginine to trypanothione. Amino Acids 2010, 40, 269–285. [Google Scholar] [CrossRef]
- Heby, O.; Persson, L.; Rentala, M. Targeting the polyamine biosynthetic enzymes: A promising approach to therapy of African sleeping sickness, Chagas’ disease, and leishmaniasis. Amino Acids 2007, 33, 359–366. [Google Scholar] [CrossRef]
- Ilari, A.; Fiorillo, A.; Baiocco, P.; Poser, E.; Angiulli, G.; Colotti, G. Targeting polyamine metabolism for finding new drugs against leishmaniasis: A review. Mini-Rev. Med. Chem. 2015, 15, 243–252. [Google Scholar] [CrossRef]
- Ilari, A.; Fiorillo, A.; Genovese, I.; Colotti, G. Polyamine-trypanothione pathway: An update. Futur. Med. Chem. 2017, 9, 61–77. [Google Scholar] [CrossRef]
- Phillips, M.A. Polyamines in protozoan pathogens. J. Biol. Chem. 2018, 293, 18746–18756. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S. Parasite Polyamines as Pharmaceutical Targets. Curr. Pharm. Des. 2017, 23, 3325–3341. [Google Scholar] [CrossRef]
- Babokhov, P.; Sanyaolu, A.O.; Oyibo, W.A.; Fagbenro-Beyioku, A.F.; Iriemenam, N.C. A current analysis of chemotherapy strategies for the treatment of human African trypanosomiasis. Pathog. Glob. Health 2013, 107, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Ebikeme, C. The Death and Life of the Resurrection Drug. PLOS Neglected Trop. Dis. 2014, 8, e2910. [Google Scholar] [CrossRef] [Green Version]
- Eperon, G.; Balasegaram, M.; Potet, J.; Mowbray, C.; Valverde, O.; Chappuis, F. Treatment options for second-stage gambiense human African trypanosomiasis. Expert Rev. Anti-Infect. Ther. 2014, 12, 1407–1417. [Google Scholar] [CrossRef] [Green Version]
- Logiudice, N.; Le, L.; Abuan, I.; Leizorek, Y.; Roberts, S.C. Alpha-Difluoromethylornithine, an Irreversible Inhibitor of Polyamine Biosynthesis, as a Therapeutic Strategy against Hyperproliferative and Infectious Diseases. Med. Sci. 2018, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Casero, R.A., Jr.; Stewart, T.M.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Damiani, E.; Wallace, H.M. Polyamines and Cancer. Methods Mol. Biol. 2018, 1694, 469–488. [Google Scholar] [CrossRef]
- Gerner, E.W.; Bruckheimer, E.; Cohen, A. Cancer pharmacoprevention: Targeting polyamine metabolism to manage risk factors for colon cancer. J. Biol. Chem. 2018, 293, 18770–18778. [Google Scholar] [CrossRef] [Green Version]
- McNamara, K.M.; Gobert, A.P.; Wilson, K.T. The role of polyamines in gastric cancer. Oncogene 2021, 40, 4399–4412. [Google Scholar] [CrossRef]
- Nakanishi, S.; Cleveland, J. Polyamine Homeostasis in Development and Disease. Med. Sci. 2021, 9, 28. [Google Scholar] [CrossRef]
- Madeo, F.; Bauer, M.A.; Carmona-Gutierrez, D.; Kroemer, G. Spermidine: A physiological autophagy inducer acting as an anti-aging vitamin in humans? Autophagy 2019, 15, 165–168. [Google Scholar] [CrossRef]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359, eaan2788. [Google Scholar] [CrossRef] [Green Version]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Sagar, N.; Tarafdar, S.; Agarwal, S.; Tarafdar, A.; Sharma, S. Polyamines: Functions, Metabolism, and Role in Human Disease Management. Med. Sci. 2021, 9, 44. [Google Scholar] [CrossRef]
- Casero, R.; Stewart, T.; Devereux, W.; Hacker, A.; Wang, Y.; Smith, R.; Woster, P.; Woster, P. The role of polyamine catabolism in anti-tumour drug response. Biochem. Soc. Trans. 2003, 31, 361–365. [Google Scholar] [CrossRef]
- Stewart, T.M.; Dunston, T.T.; Woster, P.M.; Casero, R.A., Jr. Polyamine catabolism and oxidative damage. J. Biol. Chem. 2018, 293, 18736–18745. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, K.; Barone, S.; Soleimani, M. Polyamine Catabolism in Acute Kidney Injury. Int. J. Mol. Sci. 2019, 20, 4790. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, K.; Kashiwagi, K. The functional role of polyamines in eukaryotic cells. Int. J. Biochem. Cell Biol. 2018, 107, 104–115. [Google Scholar] [CrossRef]
- Lightfoot, H.L.; Hall, J. Endogenous polyamine function—the RNA perspective. Nucleic Acids Res. 2014, 42, 11275–11290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, J.I.; Laranjeira-Silva, M.F.; Muxel, S.M.; Floeter-Winter, L.M. The impact of arginase activity on virulence factors of Leishmania amazonensis. Curr. Opin. Microbiol. 2019, 52, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Bisceglia, J.; Mollo, M.C.; Gruber, N.; Orelli, L.R. Polyamines and Related Nitrogen Compounds in the Chemotherapy of Neglected Diseases Caused by Kinetoplastids. Curr. Top. Med. Chem. 2018, 18, 321–368. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.; Stamper, B.; Elbarbry, F.; Nguyen, V.; Lopez, S.; Kawasaki, Y.; Poormohamadian, R.; Roberts, S. Natural Products That Target the Arginase in Leishmania Parasites Hold Therapeutic Promise. Microorganisms 2021, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Willert, E.; Phillips, M.A. Regulation and function of polyamines in African trypanosomes. Trends Parasitol. 2012, 28, 66–72. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. 2010, 42, 39–51. [Google Scholar] [CrossRef]
- Jiang, Y.; Roberts, S.C.; Jardim, A.; Carter, N.S.; Shih, S.; Ariyanayagam, M.; Fairlamb, A.H.; Ullman, B. Ornithine Decarboxylase Gene Deletion Mutants of Leishmania donovani. J. Biol. Chem. 1999, 274, 3781–3788. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Emmett, K.; McCANN, P.P.; Sjoerdsma, A.; Ullman, B. Effects of DL-alpha-difluoromethylornithine on Leishmania donovani promastigotes. J. Protozool. 1986, 33, 518–521. [Google Scholar] [CrossRef]
- Singh, S.; Mukherjee, A.; Khomutov, A.R.; Persson, L.; Heby, O.; Chatterjee, M.; Madhubala, R. Antileishmanial Effect of 3-Aminooxy-1-Aminopropane Is Due to Polyamine Depletion. Antimicrob. Agents Chemother. 2007, 51, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E.; Kameji, T.; Shirahata, A.; Stanley, B.; Madhubala, R.; Pajunen, A. Regulation of mammalian S-Adenosylmethionine decarboxylase. Adv. Enzym. Regul. 1988, 27, 31–39. [Google Scholar] [CrossRef]
- Wilkinson, S.R.; Kelly, J.M. Trypanocidal drugs: Mechanisms, resistance and new targets. Expert Rev. Mol. Med. 2009, 11, e31. [Google Scholar] [CrossRef]
- Yerlikaya, A.; Stanley, B. S-Adenosylmethionine Decarboxylase Degradation by the 26 S Proteasome Is Accelerated by Substrate-mediated Transamination. J. Biol. Chem. 2004, 279, 12469–12478. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.A.; Coffino, P.; Wang, C.C. Cloning and sequencing of the ornithine decarboxylase gene from Trypanosoma brucei. Implications for enzyme turnover and selective difluoromethylornithine inhibition. J. Biol. Chem. 1987, 262, 8721–8727. [Google Scholar] [CrossRef]
- Battista, T.; Colotti, G.; Ilari, A.; Fiorillo, A. Targeting Trypanothione Reductase, a Key Enzyme in the Redox Trypanosomatid Metabolism, to Develop New Drugs against Leishmaniasis and Trypanosomiases. Molecules 2020, 25, 1924. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Garg, G.; Ali, V. Current Therapeutics, Their Problems and Thiol Metabolism as Potential Drug Targets in Leishmaniasis. Curr. Drug Metab. 2016, 17, 897–919. [Google Scholar] [CrossRef]
- Afanador, G.A.; Tomchick, D.; Phillips, M.A.; Afanador, G.A.; Tomchick, D.; Phillips, M.A.; Afanador, G.A.; Tomchick, D.; Phillips, M.A.; Afanador, G.A.; et al. Trypanosomatid Deoxyhypusine Synthase Activity Is Dependent on Shared Active-Site Complementation between Pseudoenzyme Paralogs. Structure 2018, 26, 1499–1512. [Google Scholar] [CrossRef] [Green Version]
- Chawla, B.; Jhingran, A.; Singh, S.; Tyagi, N.; Park, M.H.; Srinivasan, N.; Roberts, S.C.; Madhubala, R. Identification and Characterization of a Novel Deoxyhypusine Synthase in Leishmania donovani. J. Biol. Chem. 2010, 285, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, S.; Jones, D.C.; Wyllie, S.; Fairlamb, A.H.; Phillips, M.A. Allosteric Activation of Trypanosomatid Deoxyhypusine Synthase by a Catalytically Dead Paralog. J. Biol. Chem. 2013, 288, 15256–15267. [Google Scholar] [CrossRef] [Green Version]
- Boitz, J.M.; Gilroy, C.A.; Olenyik, T.D.; Paradis, D.; Perdeh, J.; Dearman, K.; Davis, M.J.; Yates, P.A.; Li, Y.; Riscoe, M.K.; et al. Arginase Is Essential for Survival of Leishmania donovani Promastigotes but Not Intracellular Amastigotes. Infect. Immun. 2017, 85, e00554-16. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.F.L.; Zampieri, R.A.; Muxel, S.M.; Beverley, S.M.; Floeter-Winter, L.M. Leishmania amazonensis Arginase Compartmentalization in the Glycosome Is Important for Parasite Infectivity. PLoS ONE 2012, 7, e34022. [Google Scholar] [CrossRef] [Green Version]
- Reguera, R.M.; Balaña-Fouce, R.; Showalter, M.; Hickerson, S.; Beverley, S.M. Leishmania major lacking arginase (ARG) are auxotrophic for polyamines but retain infectivity to susceptible BALB/c mice. Mol. Biochem. Parasitol. 2009, 165, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.C.; Tancer, M.J.; Polinsky, M.R.; Gibson, K.M.; Heby, O.; Ullman, B. Arginase Plays a Pivotal Role in Polyamine Precursor Metabolism in Leishmania. J. Biol. Chem. 2004, 279, 23668–23678. [Google Scholar] [CrossRef] [Green Version]
- Hai, Y.; Kerkhoven, E.J.; Barrett, M.P.; Christianson, D.W. Crystal Structure of an Arginase-like Protein from Trypanosoma brucei That Evolved without a Binuclear Manganese Cluster. Biochemistry 2014, 54, 458–471. [Google Scholar] [CrossRef] [Green Version]
- Vincent, I.M.; Creek, D.J.; Burgess, K.; Woods, D.J.; Burchmore, R.J.S.; Barrett, M.P. Untargeted Metabolomics Reveals a Lack Of Synergy between Nifurtimox and Eflornithine against Trypanosoma brucei. PLOS Negl. Trop. Dis. 2012, 6, e1618. [Google Scholar] [CrossRef] [Green Version]
- Sakata, K.; Kashiwagi, K.; Sharmin, S.; Ueda, S.; Irie, Y.; Murotani, N.; Igarashi, K. Increase in putrescine, amine oxidase, and acrolein in plasma of renal failure patients. Biochem. Biophys. Res. Commun. 2003, 305, 143–149. [Google Scholar] [CrossRef]
- Carrillo, C.; Cejas, S.; Cortés, M.; Ceriani, C.; Huber, A.; González, N.S.; Algranati, I.D. Sensitivity of Trypanosomatid Protozoa to DFMO and Metabolic Turnover of Ornithine Decarboxylase. Biochem. Biophys. Res. Commun. 2000, 279, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Amano, Y.; Namatame, I.; Tateishi, Y.; Honboh, K.; Tanabe, E.; Niimi, T.; Sakashita, H. Structural insights into the novel inhibition mechanism of Trypanosoma cruzi spermidine synthase. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1879–1889. [Google Scholar] [CrossRef]
- Persson, K.; Åslund, L.; Grahn, B.; Hanke, J.; Heby, O. Trypanosoma cruzi has not lost its S-adenosylmethionine decarboxylase: Characterization of the gene and the encoded enzyme. Biochem. J. 1998, 333, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Muxel, S.M.; Aoki, J.I.; Fernandes, J.C.R.; Laranjeira-Silva, M.F.; Zampieri, R.A.; Acuña, S.M.; Müller, K.E.; Vanderlinde, R.H.; Floeter-Winter, L.M.; Muxel, S.M.; et al. Arginine and Polyamines Fate in Leishmania Infection. Front. Microbiol. 2018, 8, 2682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessenda, G.; Da Silva, J.S. Arginase and its mechanisms in Leishmania persistence. Parasite Immunol. 2020, 42, e12722. [Google Scholar] [CrossRef] [PubMed]
- Leroux, A.E.; Krauth-Siegel, R.L. Thiol redox biology of trypanosomatids and potential targets for chemotherapy. Mol. Biochem. Parasitol. 2016, 206, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Piñeyro, M.D.; Arias, D.; Parodi-Talice, A.; Guerrero, S.; Robello, C. Trypanothione Metabolism as Drug Target for Trypanosomatids. Curr. Pharm. Des. 2021, 27, 1834–1846. [Google Scholar] [CrossRef]
- Saccoliti, F.; Di Santo, R.; Costi, R. Recent Advancement in the Search of Innovative Antiprotozoal Agents Targeting Trypanothione Metabolism. ChemMedChem 2020, 15, 2420–2435. [Google Scholar] [CrossRef]
- McConville, M.J.; DE Souza, D.; Saunders, E.; Likic, V.A.; Naderer, T. Living in a phagolysosome; metabolism of Leishmania amastigotes. Trends Parasitol. 2007, 23, 368–375. [Google Scholar] [CrossRef]
- McConville, M.J.; Naderer, T. Metabolic Pathways Required for the Intracellular Survival of Leishmania. Annu. Rev. Microbiol. 2011, 65, 543–561. [Google Scholar] [CrossRef]
- McConville, M.J.; Saunders, E.C.; Kloehn, J.; Dagley, M.J. Leishmania carbon metabolism in the macrophage phagolysosome- feast or famine? F1000Research 2015, 4, 938. [Google Scholar] [CrossRef]
- Naderer, T.; McConville, M.J. The Leishmania-macrophage interaction: A metabolic perspective. Cell. Microbiol. 2007, 10, 301–308. [Google Scholar] [CrossRef]
- Saunders, E.C.; Sernee, M.F.; Ralton, J.E.; McConville, M.J. Metabolic stringent response in intracellular stages of Leishmania. Curr. Opin. Microbiol. 2021, 63, 126–132. [Google Scholar] [CrossRef]
- Ferreira, C.; Estaquier, J.; Silvestre, R. Immune-metabolic interactions between Leishmania and macrophage host. Curr. Opin. Microbiol. 2021, 63, 231–237. [Google Scholar] [CrossRef]
- Kima, P.E. The amastigote forms of Leishmania are experts at exploiting host cell processes to establish infection and persist. Int. J. Parasitol. 2007, 37, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Goldman-Pinkovich, A.; Kannan, S.; Nitzan-Koren, R.; Puri, M.; Pawar, H.; Bar-Avraham, Y.; McDonald, J.; Sur, A.; Zhang, W.-W.; Matlashewski, G.; et al. Sensing Host Arginine Is Essential for Leishmania Parasites’ Intracellular Development. mBio 2020, 11, e02023-20. [Google Scholar] [CrossRef]
- Pawar, H.; Puri, M.; Weinberger, R.F.; Madhubala, R.; Zilberstein, D. The arginine sensing and transport binding sites are distinct in the human pathogen Leishmania. PLOS Negl. Trop. Dis. 2019, 13, e0007304. [Google Scholar] [CrossRef] [Green Version]
- Zilberstein, D.; Myler, P.J. Arginine sensing in intracellular parasitism of Leishmania. Curr. Opin. Microbiol. 2021, 64, 41–46. [Google Scholar] [CrossRef]
- Balaña-Fouce, R.; Calvo-Álvarez, E.; Álvarez-Velilla, R.; Prada, C.F.; Pérez-Pertejo, Y.; Reguera, R.M. Role of trypanosomatid’s arginase in polyamine biosynthesis and pathogenesis. Mol. Biochem. Parasitol. 2012, 181, 85–93. [Google Scholar] [CrossRef]
- Wanasen, N.; Soong, L. L-arginine metabolism and its impact on host immunity against Leishmania infection. Immunol. Res. 2007, 41, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Badirzadeh, A.; Taheri, T.; Taslimi, Y.; Abdossamadi, Z.; Heidari-Kharaji, M.; Gholami, E.; Sedaghat, B.; Niyyati, M.; Rafati, S. Arginase activity in pathogenic and non-pathogenic species of Leishmania parasites. PLOS Negl. Trop. Dis. 2017, 11, e0005774. [Google Scholar] [CrossRef] [Green Version]
- Iniesta, V.; Carcelén, J.; Molano, I.; Peixoto, P.M.V.; Redondo, E.; Parra, P.; Mangas, M.; Monroy, I.; Campo, M.L.; Nieto, C.G.; et al. Arginase I Induction during Leishmania major Infection Mediates the Development of Disease. Infect. Immun. 2005, 73, 6085–6090. [Google Scholar] [CrossRef] [Green Version]
- Iniesta, V.; Gomez-Nieto, L.C.; Molano, I.; Mohedano, A.; Carcelen, J.; Miron, C.; Alonso, C.; Corraliza, I. Arginase I induction in macrophages, triggered by Th2-type cytokines, supports the growth of intracellular Leishmania parasites. Parasite Immunol. 2002, 24, 113–118. [Google Scholar] [CrossRef]
- Kropf, P.; Fuentes, J.M.; Fähnrich, E.; Arpa, L.; Herath, S.; Weber, V.; Soler, G.; Celada, A.; Modolell, M.; Müller, I. Arginase and polyamine synthesis are key factors in the regulation of experimental leishmaniasis in vivo. FASEB J. 2005, 19, 1000–1002. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, H.; Sadeghipour, P.; Taslimi, Y.; Habibzadeh, S.; Zali, F.; Zahedifard, F.; Rahmati, J.; Kamyab, K.; Ghandi, N.; Zamanian, A.; et al. Comparing acute and chronic human cutaneous leishmaniasis caused byLeishmania major and Leishmania tropica focusing on arginase activity. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 2118–2121. [Google Scholar] [CrossRef] [PubMed]
- Müller, I.; Hailu, A.; Choi, B.-S.; Abebe, T.; Fuentes, J.M.; Munder, M.; Modolell, M.; Kropf, P. Age-Related Alteration of Arginase Activity Impacts on Severity of Leishmaniasis. PLOS Negl. Trop. Dis. 2008, 2, e235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins-Rodríguez, A.A.; Pérez-Torres, A.; Escalona-Montaño, A.R.; Gutiérrez-Kobeh, L. Differential Regulation of l -Arginine Metabolism through Arginase 1 during Infection with Leishmania mexicana Isolates Obtained from Patients with Localized and Diffuse Cutaneous Leishmaniasis. Infect. Immun. 2020, 88, e00963-19. [Google Scholar] [CrossRef]
- Choi, B.-S.; Martinez-Falero, I.C.; Corset, C.; Munder, M.; Modolell, M.; Muller, I.; Kropf, P. Differential impact of L-arginine deprivation on the activation and effector functions of T cells and macrophages. J. Leukoc. Biol. 2008, 85, 268–277. [Google Scholar] [CrossRef]
- Modolell, M.; Choi, B.-S.; Ryan, R.O.; Hancock, M.; Titus, R.G.; Abebe, T.; Hailu, A.; Müller, I.; Rogers, M.E.; Bangham, C.; et al. Local Suppression of T Cell Responses by Arginase-Induced L-Arginine Depletion in Nonhealing Leishmaniasis. PLOS Negl. Trop. Dis. 2009, 3, e480. [Google Scholar] [CrossRef] [Green Version]
- Munder, M.; Choi, B.-S.; Rogers, M.; Kropf, P. L -Arginine deprivation impairs Leishmania major -specific T-cell responses. Eur. J. Immunol. 2009, 39, 2161–2172. [Google Scholar] [CrossRef] [Green Version]
- Kropf, P.; Herath, S.; Weber, V.; Modolell, M.; Muller, I. Factors influencing Leishmania major infection in IL-4-deficient BALB/c mice. Parasite Immunol. 2003, 25, 439–447. [Google Scholar] [CrossRef]
- Boitz, J.M.; Yates, P.A.; Kline, C.; Gaur, U.; Wilson, M.E.; Ullman, B.; Roberts, S.C. Leishmania donovani Ornithine Decarboxylase Is Indispensable for Parasite Survival in the Mammalian Host. Infect. Immun. 2009, 77, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Gilroy, C.; Olenyik, T.; Roberts, S.C.; Ullman, B. Spermidine Synthase Is Required for Virulence of Leishmania donovani. Infect. Immun. 2011, 79, 2764–2769. [Google Scholar] [CrossRef] [Green Version]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in aging and disease. Aging 2011, 3, 716–732. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef]
- Olenyik, T.; Gilroy, C.; Ullman, B. Oral putrescine restores virulence of ornithine decarboxylase-deficient Leishmania donovani in mice. Mol. Biochem. Parasitol. 2011, 176, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Gaur, U.; Roberts, S.C.; Dalvi, R.P.; Corraliza, I.; Ullman, B.; Wilson, M. An Effect of Parasite-Encoded Arginase on the Outcome of Murine Cutaneous Leishmaniasis. J. Immunol. 2007, 179, 8446–8453. [Google Scholar] [CrossRef]
- Hanson, S.; Adelman, J.; Ullman, B. Amplification and molecular cloning of the ornithine decarboxylase gene of Leishmania donovani. J. Biol. Chem. 1992, 267, 2350–2359. [Google Scholar] [CrossRef]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The eukaryotic pathogen, vector and host bioinformatics resource center. Nucleic Acids Res. 2021, 50, D898–D911. [Google Scholar] [CrossRef]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. TriTrypDB: A functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2009, 38, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Singh, S.; Dubey, V.K. Novel Inhibitors of Ornithine Decarboxylase of Leishmania Parasite (LdODC): The Parasite Resists LdODC Inhibition by Overexpression of Spermidine Synthase. Chem. Biol. Drug Des. 2015, 87, 352–360. [Google Scholar] [CrossRef]
- Pandey, R.K.; Prajapati, P.; Goyal, S.; Grover, A.; Prajapati, V. Molecular Modeling and Virtual Screening Approach to Discover Potential Antileishmanial Inhibitors Against Ornithine Decarboxylase. Comb. Chem. High Throughput Screen. 2016, 19, 813–823. [Google Scholar] [CrossRef]
- Dufe, V.T.; Ingner, D.; Heby, O.; Khomutov, A.R.; Persson, L.; Al-Karadaghi, S. A structural insight into the inhibition of human and Leishmania donovani ornithine decarboxylases by 1-amino-oxy-3-aminopropane. Biochem. J. 2007, 405, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Osterman, A.; Grishin, N.V.; Kinch, L.; Phillips, M.A. Formation of Functional Cross-Species Heterodimers of Ornithine Decarboxylase. Biochemistry 1994, 33, 13662–13667. [Google Scholar] [CrossRef]
- Osterman, A.L.; Kinch, L.N.; Grishin, N.V.; Phillips, M.A. Acidic Residues Important for Substrate Binding and Cofactor Reactivity in Eukaryotic Ornithine Decarboxylase Identified by Alanine Scanning Mutagenesis. J. Biol. Chem. 1995, 270, 11797–11802. [Google Scholar] [CrossRef] [Green Version]
- Almrud, J.J.; Oliveira, M.A.; Kern, A.D.; Grishin, N.V.; Phillips, M.A.; Hackert, M.L. Crystal structure of human ornithine decarboxylase at 2.1 å resolution: Structural insights to antizyme binding. J. Mol. Biol. 2000, 295, 7–16. [Google Scholar] [CrossRef]
- Grishin, N.V.; Osterman, A.L.; Brooks, H.B.; Phillips, M.A.; Goldsmith, E.J. X-ray Structure of Ornithine Decarboxylase from Trypanosoma brucei: The Native Structure and the Structure in Complex with α-Difluoromethylornithine. Biochemistry 1999, 38, 15174–15184. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.K.; Brooks, H.B.; Osterman, A.L.; Goldsmith, E.J.; Phillips, M.A. Altering the Reaction Specificity of Eukaryotic Ornithine Decarboxylase. Biochemistry 2000, 39, 11247–11257. [Google Scholar] [CrossRef] [PubMed]
- Kern, A.D.; Oliveira, M.A.; Coffino, P.; Hackert, M.L. Structure of mammalian ornithine decarboxylase at 1.6 Å resolution: Stereochemical implications of PLP-dependent amino acid decarboxylases. Structure 1999, 7, 567–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwami, K.; Wang, J.Y.; Jain, R.; McCormack, S.; Johnson, L.R. Intestinal ornithine decarboxylase: Half-life and regulation by putrescine. Am. J. Physiol. Liver Physiol. 1990, 258, G308–G315. [Google Scholar] [CrossRef]
- Ghoda, L.; Phillips, M.A.; Bass, K.E.; Wang, C.C.; Coffino, P. Trypanosome ornithine decarboxylase is stable because it lacks sequences found in the carboxyl terminus of the mouse enzyme which target the latter for intracellular degradation. J. Biol. Chem. 1990, 265, 11823–11826. [Google Scholar] [CrossRef] [PubMed]
- Perdeh, J.; Berioso, B.; Love, Q.; Logiudice, N.; Le, T.L.; Harrelson, J.P.; Roberts, S.C. Critical functions of the polyamine putrescine for proliferation and viability of Leishmania donovani parasites. Amino Acids 2019, 52, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, G.A.; Lianos, G.D.; Ragos, V.; Galani, V.; Kyritsis, A.P. Difluoromethylornithine in cancer: New advances. Futur. Oncol. 2017, 13, 809–819. [Google Scholar] [CrossRef]
- Bachmann, A.S.; Geerts, D. Polyamine synthesis as a target of MYC oncogenes. J. Biol. Chem. 2018, 293, 18757–18769. [Google Scholar] [CrossRef] [Green Version]
- Bassiri, H.; Benavides, A.; Haber, M.; Gilmour, S.K.; Norris, M.D.; Hogarty, M.D. Translational development of difluoromethylornithine (DFMO) for the treatment of neuroblastoma. Transl. Pediatr. 2015, 4, 226–238. [Google Scholar] [CrossRef]
- Somani, R.R.; Kandpile, P.S. Ornithine Decarboxylase Inhibition: A Strategy to Combat Various Diseases. Mini Rev. Med. Chem. 2018, 18, 1008–1021. [Google Scholar] [CrossRef]
- Burri, C.; Brun, R. Eflornithine for the treatment of human African trypanosomiasis. Parasitol. Res. 2003, 90 (Suppl. 1), S49–S52. [Google Scholar] [CrossRef]
- Reguera, R.M.; Fouce, R.B.; Cubría, J.; Bujidos, M.A.; Ordóñez, D. Fluorinated analogues of L-ornithine are powerful inhibitors of ornithine decarboxylase and cell growth of Leishmania infantum promastigotes. Life Sci. 1994, 56, 223–230. [Google Scholar] [CrossRef]
- Sánchez, C.P.; Mucci, J.; González, N.S.; Ochoa, A.; Zakin, M.M.; Algranati, I.D. α-Difluoromethylornithine-resistant cell lines obtained after one-step selection of Leishmania mexicana promastigote cultures. Biochem. J. 1997, 324, 847–853. [Google Scholar] [CrossRef]
- Roberts, S.C.; Jiang, Y.; Gasteier, J.; Frydman, B.; Marton, L.J.; Heby, O.; Ullman, B. Leishmania donovani Polyamine Biosynthetic Enzyme Overproducers as Tools To Investigate the Mode of Action of Cytotoxic Polyamine Analogs. Antimicrob. Agents Chemother. 2007, 51, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Hanson, S.; Beverley, S.M.; Wagner, W.; Ullman, B. Unstable amplification of two extrachromosomal elements in alpha-difluoromethylornithine-resistant Leishmania donovani. Mol. Cell. Biol. 1992, 12, 5499–5507. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Kapoor, P.; Madhubala, R. Characterization of alpha-difluoromethylornithine resistant Leishmania donovani and its susceptibility to other inhibitors of the polyamine biosynthetic pathway. Pharmacol. Res. 1996, 34, 43–46. [Google Scholar] [CrossRef]
- Keithly, J.S.; Fairlamb, A.H. Inhibition of Leishmania Species by α-Difluoromethylornithine. Leishmaniasis 1989, 171, 749–756. [Google Scholar] [CrossRef]
- Gradoni, L.; Iorio, M.A.; Gramiccia, M.; Orsini, S. In vivo effect of eflornithine (DFMO) and some related compounds on Leishmania infantum preliminary communication. Farmaco 1989, 44, 1157–1166. [Google Scholar]
- Mukhopadhyay, R.; Madhubala, R. Effect of a Bis(Benzyl)Polyamine Analogue, and DL-α-difluoromethylornithine on Parasite Suppression and Cellular Polyamine Levels in Golden Hamster During Leishmania Donovani Infection. Pharmacol. Res. 1993, 28, 359–366. [Google Scholar] [CrossRef]
- Khomutov, M.A.; Mandal, S.; Weisell, J.; Saxena, N.; Simonian, A.R.; Vepsalainen, J.; Madhubala, R.; Kochetkov, S.N. Novel convenient synthesis of biologically active esters of hydroxylamine. Amino Acids 2009, 38, 509–517. [Google Scholar] [CrossRef]
- Singh, S.; Jhingran, A.; Sharma, A.; Simonian, A.R.; Soininen, P.; Vepsalainen, J.; Khomutov, A.R.; Madhubala, R. Novel agmatine analogue, γ-guanidinooxypropylamine (GAPA) efficiently inhibits proliferation of Leishmania donovani by depletion of intracellular polyamine levels. Biochem. Biophys. Res. Commun. 2008, 375, 168–172. [Google Scholar] [CrossRef]
- Vannier-Santos, M.A.; Menezes, D.; Oliveira, M.F.; de Mello, F.G. The putrescine analogue 1,4-diamino-2-butanone affects polyamine synthesis, transport, ultrastructure and intracellular survival in Leishmania amazonensis. Microbiology 2008, 154, 3104–3111. [Google Scholar] [CrossRef] [Green Version]
- Menezes, D.; Valentim, C.; Oliveira, M.F.; Vannier-Santos, M.A. Putrescine analogue cytotoxicity against Trypanosoma cruzi. Parasitol. Res. 2005, 98, 99–105. [Google Scholar] [CrossRef]
- Baumann, R.J.; Hanson, W.L.; McCann, P.P.; Sjoerdsma, A.; Bitonti, A.J. Suppression of both antimony-susceptible and antimony-resistant Leishmania donovani by a bis(benzyl)polyamine analog. Antimicrob. Agents Chemother. 1990, 34, 722–727. [Google Scholar] [CrossRef] [Green Version]
- Baumann, R.J.; McCann, P.P.; Bitonti, A.J. Suppression of Leishmania donovani by oral administration of a bis(benzyl)polyamine analog. Antimicrob. Agents Chemother. 1991, 35, 1403–1407. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, R. Antileishmanial effect of a potent s-adenosylmethionine decarboxylase inhibitor: CGP 40215A. Pharmacol. Res. 1996, 33, 67–70. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Madhubala, R. Effects of Bis(benzyl)polyamine Analogs on Leishmania donovani Promastigotes. Exp. Parasitol. 1995, 81, 39–46. [Google Scholar] [CrossRef]
- Chakraborty, D.; Saravanan, P.; Patra, S.; Dubey, V.K. Studies on ornithine decarboxylase of Leishmania donovani: Structure modeling and inhibitor docking. Med. Chem. Res. 2012, 22, 466–478. [Google Scholar] [CrossRef]
- Grover, A.; Katiyar, S.; Singh, S.K.; Dubey, V.; Sundar, D. A leishmaniasis study: Structure-based screening and molecular dynamics mechanistic analysis for discovering potent inhibitors of spermidine synthase. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2012, 1824, 1476–1483. [Google Scholar] [CrossRef]
- Herrera-Acevedo, C.; Dos Santos Maia, M.; Cavalcanti, E.; Coy-Barrera, E.; Scotti, L.; Scotti, M.T. Selection of antileishmanial sesquiterpene lactones from SistematX database using a combined ligand-/structure-based virtual screening approach. Mol. Divers 2020, 25, 2411–2427. [Google Scholar] [CrossRef]
- Hazra, S.; Ghosh, S.; Das Sarma, M.; Sharma, S.; Das, M.; Saudagar, P.; Prajapati, V.K.; Dubey, V.K.; Sundar, S.; Hazra, B. Evaluation of a diospyrin derivative as antileishmanial agent and potential modulator of ornithine decarboxylase of Leishmania donovani. Exp. Parasitol. 2013, 135, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Hazra, B.; Mittra, B.; Das, A.; Majumder, H.K. Diospyrin, A Bisnaphthoquinone: A Novel Inhibitor of Type I DNA Topoisomerase of Leishmania donovani. Mol. Pharmacol. 1998, 54, 994–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, P.; Majee, S.B.; Ghosh, S.; Hazra, B. Apoptosis-like death in Leishmania donovani promastigotes induced by diospyrin and its ethanolamine derivative. Int. J. Antimicrob. Agents 2009, 34, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.C.; Scott, J.; Gasteier, J.E.; Jiang, Y.; Brooks, B.; Jardim, A.; Carter, N.S.; Heby, O.; Ullman, B. S-Adenosylmethionine Decarboxylase from Leishmania donovani. J. Biol. Chem. 2002, 277, 5902–5909. [Google Scholar] [CrossRef] [Green Version]
- Taladriz, S.; Ramiro, M.J.; Hanke, T.; Larraga, V. S-Adenosylmethionine decarboxylase from Leishmania infantum promastigotes: Molecular cloning and differential expression. Parasitol. Res. 2002, 88, 421–426. [Google Scholar] [CrossRef]
- Willert, E.K.; Phillips, M.A. Regulated Expression of an Essential Allosteric Activator of Polyamine Biosynthesis in African Trypanosomes. PLoS Pathog. 2008, 4, e1000183. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Nguyen, S.; Kim, S.H.; Volkov, O.A.; Tu, B.P.; Phillips, M.A. Product feedback regulation implicated in translational control of the Trypanosoma brucei S-adenosylmethionine decarboxylase regulatory subunit prozyme. Mol. Microbiol. 2013, 88, 846–861. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Agnihotri, P.; Pratap, J.V. Characterization of a Novel Putative S-Adenosylmethionine Decarboxylase-Like Protein from Leishmania donovani. PLoS ONE 2013, 8, e65912. [Google Scholar] [CrossRef] [Green Version]
- Willert, E.K.; Fitzpatrick, R.; Phillips, M.A. Allosteric regulation of an essential trypanosome polyamine biosynthetic enzyme by a catalytically dead homolog. Proc. Natl. Acad. Sci. USA 2007, 104, 8275–8280. [Google Scholar] [CrossRef] [Green Version]
- Bale, S.; Ealick, S.E. Structural biology of S-adenosylmethionine decarboxylase. Amino Acids 2009, 38, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. S-Adenosylmethionine decarboxylase. Essays Biochem. 2009, 46, 25–46. [Google Scholar] [CrossRef] [Green Version]
- Stanley, B.; Shantz, L.; Pegg, A. Expression of mammalian S-adenosylmethionine decarboxylase in Escherichia coli. Determination of sites for putrescine activation of activity and processing. J. Biol. Chem. 1994, 269, 7901–7907. [Google Scholar] [CrossRef]
- Pajunen, A.; Crozat, A.; Janne, O.A.; Ihalainen, R.; Laitinen, P.H.; Stanley, B.; Madhubala, R.; Pegg, A.E. Structure and regulation of mammalian S-adenosylmethionine decarboxylase. J. Biol. Chem. 1988, 263, 17040–17049. [Google Scholar] [CrossRef]
- Stanley, B.; Pegg, A. Amino acid residues necessary for putrescine stimulation of human S-adenosylmethionine decarboxylase proenzyme processing and catalytic activity. J. Biol. Chem. 1991, 266, 18502–18506. [Google Scholar] [CrossRef]
- Clyne, T.; Kinch, L.N.; Phillips, M.A. Putrescine Activation of Trypanosoma cruzi S-Adenosylmethionine Decarboxylase. Biochemistry 2002, 41, 13207–13216. [Google Scholar] [CrossRef]
- Persson, L. Ornithine decarboxylase and S-adenosylmethionine decarboxylase in trypanosomatids. Biochem. Soc. Trans. 2007, 35, 314–317. [Google Scholar] [CrossRef] [Green Version]
- Willert, E.K.; Phillips, M.A. Cross-species activation of trypanosome S-adenosylmethionine decarboxylase by the regulatory subunit prozyme. Mol. Biochem. Parasitol. 2009, 168, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.M.; Volkov, O.A.; Leija, C.; Lemoff, A.; Phillips, M.A. A dual regulatory circuit consisting of S-adenosylmethionine decarboxylase protein and its reaction product controls expression of the paralogous activator prozyme in Trypanosoma brucei. PLOS Pathog. 2018, 14, e1007404. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; McCloskey, D.E.; Phillips, M.A. RNA Interference-Mediated Silencing of Ornithine Decarboxylase and Spermidine Synthase Genes in Trypanosoma brucei Provides Insight into Regulation of Polyamine Biosynthesis. Eukaryot. Cell 2009, 8, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Agnihotri, P.; Srivastava, V.K.; Pratap, J.V. Novel protein–protein interaction between spermidine synthase and S-adenosylmethionine decarboxylase from Leishmania donovani. Biochem. Biophys. Res. Commun. 2015, 456, 637–642. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Madhubala, R. Antileishmanial activity of berenil and methylglyoxal bis (guanylhydrazone) and its correlation with S-adenosylmethionine decarboxylase and polyamines. Int. J. Biochem. Cell Biol. 1995, 27, 55–59. [Google Scholar] [CrossRef]
- Brockway, A.J.; Volkov, O.A.; Cosner, C.C.; MacMillan, K.S.; Wring, S.A.; Richardson, T.E.; Peel, M.; Phillips, M.A.; De Brabander, J.K. Synthesis and evaluation of analogs of 5′-(((Z)-4-amino-2-butenyl)methylamino)-5′-deoxyadenosine (MDL 73811, or AbeAdo)—An inhibitor of S-adenosylmethionine decarboxylase with antitrypanosomal activity. Bioorganic Med. Chem. 2017, 25, 5433–5440. [Google Scholar] [CrossRef]
- Volkov, O.A.; Brockway, A.J.; Wring, S.A.; Peel, M.; Chen, Z.; Phillips, M.A.; De Brabander, J.K. Species-Selective Pyrimidineamine Inhibitors of Trypanosoma brucei S-Adenosylmethionine Decarboxylase. J. Med. Chem. 2018, 61, 1182–1203. [Google Scholar] [CrossRef]
- Roberts, S.C.; Jiang, Y.; Jardim, A.; Carter, N.S.; Heby, O.; Ullman, B. Genetic analysis of spermidine synthase from Leishmania donovani. Mol. Biochem. Parasitol. 2001, 115, 217–226. [Google Scholar] [CrossRef]
- Dufe, V.T.; Qiu, W.; Müller, I.B.; Hui, R.; Walter, R.D.; Al-Karadaghi, S. Crystal Structure of Plasmodium falciparum Spermidine Synthase in Complex with the Substrate Decarboxylated S-adenosylmethionine and the Potent Inhibitors 4MCHA and AdoDATO. J. Mol. Biol. 2007, 373, 167–177. [Google Scholar] [CrossRef]
- Singh, S.; Sarma, S.; Katiyar, S.P.; Das, M.; Bhardwaj, R.; Sundar, D.; Dubey, V.K. Probing the Molecular Mechanism of Hypericin-Induced Parasite Death Provides Insight into the Role of Spermidine beyond Redox Metabolism in Leishmania donovani. Antimicrob. Agents Chemother. 2015, 59, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Dubey, V.K. Quantitative Proteome Analysis of Leishmania donovani under Spermidine Starvation. PLoS ONE 2016, 11, e0154262. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Kumari, E.; Bhardwaj, R.; Kumar, R.; Dubey, V.K. Molecular events leading to death of Leishmania donovani under spermidine starvation after hypericin treatment. Chem. Biol. Drug Des. 2017, 90, 962–971. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, P.; Vantieghem, A.; Merlevede, W.; de Witte, P.A. Hypericin in cancer treatment: More light on the way. Int. J. Biochem. Cell Biol. 2001, 34, 221–241. [Google Scholar] [CrossRef]
- Garg, A.D.; Agostinis, P. ER stress, autophagy and immunogenic cell death in photodynamic therapy-induced anti-cancer immune responses. Photochem. Photobiol. Sci. 2014, 13, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Lopera, A.; Velásquez, A.; Clementino, L.; Robledo, S.; Montoya, A.; de Freitas, L.; Bezzon, V.; Fontana, C.R.; Garcia, C.; Graminha, M.A.S. Solution-combustion synthesis of doped TiO 2 compounds and its potential antileishmanial activity mediated by photodynamic therapy. J. Photochem. Photobiol. B Biol. 2018, 183, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Montoya, A.; Daza, A.; Muñoz, D.; Ríos, K.; Taylor, V.; Cedeño, D.; Vélez, I.D.; Echeverri, F.; Robledo, S.M. Development of a Novel Formulation with Hypericin To Treat Cutaneous Leishmaniasis Based on Photodynamic Therapy in In Vitro and in vivo Studies. Antimicrob. Agents Chemother. 2015, 59, 5804–5813. [Google Scholar] [CrossRef] [Green Version]
- Sepúlveda, A.A.L.; Velásquez, A.M.A.; Linares, I.A.P.; de Almeida, L.; Fontana, C.R.; Garcia, C.; Graminha, M.A.S. Efficacy of photodynamic therapy using TiO2 nanoparticles doped with Zn and hypericin in the treatment of cutaneous Leishmaniasis caused by Leishmania amazonensis. Photodiagnosis Photodyn. Ther. 2020, 30, 101676. [Google Scholar] [CrossRef]
- Vidhya, V.M.; Dubey, V.K.; Ponnuraj, K. Identification of two natural compound inhibitors of Leishmania donovani Spermidine Synthase (SpdS) through molecular docking and dynamic studies. J. Biomol. Struct. Dyn. 2017, 36, 2678–2693. [Google Scholar] [CrossRef]
- Grover, A.; Katiyar, S.; Jeyakanthan, J.; Dubey, V.; Sundar, D. Mechanistic insights into the dual inhibition strategy for checking Leishmaniasis. J. Biomol. Struct. Dyn. 2012, 30, 474–487. [Google Scholar] [CrossRef]
- Jagu, E.; Pomel, S.; Pethe, S.; Loiseau, P.M.; Labruère, R. Polyamine-based analogs and conjugates as antikinetoplastid agents. Eur. J. Med. Chem. 2017, 139, 982–1015. [Google Scholar] [CrossRef]
- Gil, Z.; Martinez-Sotillo, N.; Pinto-Martinez, A.; Mejias, F.; Martinez, J.C.; Galindo, I.; Oldfield, E.; Benaim, G. SQ109 inhibits proliferation of Leishmania donovani by disruption of intracellular Ca2+ homeostasis, collapsing the mitochondrial electrochemical potential (ΔΨm) and affecting acidocalcisomes. Parasitol. Res. 2020, 119, 649–657. [Google Scholar] [CrossRef]
- Kandpal, M.; Tekwani, B.L.; Chauhan, P.; Bhaduri, A. Correlation between inhibition of growth and arginine transport of Leishmania donovani promastigotes in vitro by diamidines. Life Sci. 1996, 59, PL175–PL180. [Google Scholar] [CrossRef]
- Tavares, J.; Ouaissi, A.; Lin, P.; Tomás, A.; Cordeiro-Da-Silva, A. Differential effects of polyamine derivative compounds against Leishmania infantum promastigotes and axenic amastigotes. Int. J. Parasitol. 2005, 35, 637–646. [Google Scholar] [CrossRef]
- Balaña-Fouce, R.; Ordóñez, D.; Alunda, J.M. Putrescine transport system in Leishmania infantum promastigotes. Mol. Biochem. Parasitol. 1989, 35, 43–50. [Google Scholar] [CrossRef]
- Basselin, M.; Coombs, G.H.; Barrett, M.P. Putrescine and spermidine transport in Leishmania. Mol. Biochem. Parasitol. 2000, 109, 37–46. [Google Scholar] [CrossRef]
- Gonzalez, N.S.; Algranati, I.D. Regulation of putrescine uptake in Leishmania mexicana promastigotes. Cell. Mol. Biol. 1994, 40, 907–914. [Google Scholar]
- Hasne, M.-P.; Ullman, B. Identification and Characterization of a Polyamine Permease from the Protozoan Parasite Leishmania major. J. Biol. Chem. 2005, 280, 15188–15194. [Google Scholar] [CrossRef] [Green Version]
- Kandpal, M.; Tekwani, B.L. Polyamine transport systems of Leishmania donovani promastigotes. Life Sci. 1997, 60, 1793–1801. [Google Scholar] [CrossRef]
- Avila, J.; Polegre, M.A. Uptake and metabolism of S-adenosyl-l-methionine by Leishmania mexicana and Leishmania braziliensis promastigotes. Mol. Biochem. Parasitol. 1993, 58, 123–134. [Google Scholar] [CrossRef]
- Dridi, L.; Ouameur, A.A.; Ouellette, M. High Affinity S-Adenosylmethionine Plasma Membrane Transporter of Leishmania Is a Member of the Folate Biopterin Transporter (FBT) Family. J. Biol. Chem. 2010, 285, 19767–19775. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, F.; Derbécourt, T.; Robert-Gero, M. Proton-ATPase activities involved in the uptake of an S-adenosylmethionine analogue. Mol. Biochem. Parasitol. 1998, 92, 99–109. [Google Scholar] [CrossRef]
- Kandpal, M.; Fouce, R.B.; Pal, A.; Guru, P.Y.; Tekwani, B.L. Kinetics and molecular characteristics of arginine transport by Leishmania donovani promastigotes. Mol. Biochem. Parasitol. 1995, 71, 193–201. [Google Scholar] [CrossRef]
- Shaked-Mishan, P.; Suter-Grotemeyer, M.; Yoel-Almagor, T.; Holland, N.; Zilberstein, D.; Rentsch, D. A novel high-affinity arginine transporter from the human parasitic protozoan Leishmania donovani. Mol. Microbiol. 2006, 60, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Alexander, E.T.; Minton, A.; Peters, M.C.; Phanstiel, O., IV; Gilmour, S.K. A novel polyamine blockade therapy activates an anti-tumor immune response. Oncotarget 2017, 8, 84140–84152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, L.D.; Purgato, S.; Murray, J.; Xiao, L.; Yu, D.M.T.; Hanssen, K.M.; Giorgi, F.M.; Carter, D.R.; Gifford, A.J.; Valli, E.; et al. Inhibition of polyamine synthesis and uptake reduces tumor progression and prolongs survival in mouse models of neuroblastoma. Sci. Transl. Med. 2019, 11, eaau1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gitto, S.B.; Pandey, V.; Oyer, J.L.; Copik, A.J.; Hogan, F.C.; Phanstiel, I.O.; Altomare, D.A. Difluoromethylornithine Combined with a Polyamine Transport Inhibitor Is Effective against Gemcitabine Resistant Pancreatic Cancer. Mol. Pharm. 2018, 15, 369–376. [Google Scholar] [CrossRef]
- Khan, A.; Gamble, L.D.; Upton, D.H.; Ung, C.; Yu, D.M.T.; Ehteda, A.; Pandher, R.; Mayoh, C.; Hébert, S.; Jabado, N.; et al. Dual targeting of polyamine synthesis and uptake in diffuse intrinsic pontine gliomas. Nat. Commun. 2021, 12, 971. [Google Scholar] [CrossRef]
- Sayer, M.H., Jr.; Reddy, V.S.; Tamang, D.G.; Västermark, Å. The Transporter Classification Database. Nucleic Acids Res. 2014, 42, D251–D258. [Google Scholar] [CrossRef]
- Mamani-Huanca, M.; Muxel, S.; Acuña, S.; Floeter-Winter, L.; Barbas, C.; López-Gonzálvez, Á. Metabolomic Reprogramming of C57BL/6-Macrophages during Early Infection with L. amazonensis. Int. J. Mol. Sci. 2021, 22, 6883. [Google Scholar] [CrossRef]
- Macedo, J.P.; Currier, R.B.; Wirdnam, C.; Horn, D.; Alsford, S.; Rentsch, D. Ornithine uptake and the modulation of drug sensitivity in Trypanosoma brucei. FASEB J. 2017, 31, 4649–4660. [Google Scholar] [CrossRef] [Green Version]
- Henriques, C.; Miller, M.P.; Catanho, M.; De Carvalho, T.M.U.; Krieger, M.A.; Probst, C.M.; De Souza, W.; Degrave, W.; Amara, S.G. Identification and functional characterization of a novel arginine/ornithine transporter, a member of a cationic amino acid transporter subfamily in the Trypanosoma cruzi genome. Parasites Vectors 2015, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Aoki, J.I.; Muxel, S.M.; Zampieri, R.A.; Acuña, S.M.; Fernandes, J.C.R.; Vanderlinde, R.H.; de Sales, M.C.O.P.; Floeter-Winter, L.M. L-arginine availability and arginase activity: Characterization of amino acid permease 3 in Leishmania amazonensis. PLOS Negl. Trop. Dis. 2017, 11, e0006025. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Salgado, A.G.; Wirdnam, C.; Meier, S.; Grotemeyer, M.S.; Inbar, E.; Mäser, P.; Zilberstein, D.; Sigel, E.; Bütikofer, P.; et al. Trypanosoma brucei eflornithine transporter AAT6 is a low-affinity low-selective transporter for neutral amino acids. Biochem. J. 2014, 463, 9–18. [Google Scholar] [CrossRef]
- Vincent, I.M.; Creek, D.; Watson, D.G.; Kamleh, M.A.; Woods, D.J.; Wong, P.E.; Burchmore, R.J.S.; Barrett, M.P. A Molecular Mechanism for Eflornithine Resistance in African Trypanosomes. PLoS Pathog. 2010, 6, e1001204. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.A.; Wang, C.C. A Trypanosoma brucei mutant resistant to α-difluoromethylornithine. Mol. Biochem. Parasitol. 1987, 22, 9–17. [Google Scholar] [CrossRef]
- Hasne, M.-P.; Coppens, I.; Soysa, R.; Ullman, B. A high-affinity putrescine-cadaverine transporter fromTrypanosoma cruzi. Mol. Microbiol. 2010, 76, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Ulrich, P.N.; Storey, M.; Johnson, D.; Tischer, J.; Tovar, J.A.; Moreno, S.N.J.; Orlando, R.; Docampo, R. Proteomic Analysis of the Acidocalcisome, an Organelle Conserved from Bacteria to Human Cells. PLoS Pathog. 2014, 10, e1004555. [Google Scholar] [CrossRef] [Green Version]
- McMillan, J.M.; Walle, U.K.; Walle, T. S-adenosyl-l-methionine: Transcellular transport and uptake by Caco-2 cells and hepatocytes. J. Pharm. Pharmacol. 2005, 57, 599–605. [Google Scholar] [CrossRef]
- Phelouzat, M.A.; Basselin, M.; Lawrence, F.; Robert-Gero, M. Sinefungin shares AdoMet-uptake system to enter Leishmania donovani promastigotes. Biochem. J. 1995, 305, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Croft, S.L.; Sundar, S.; Fairlamb, A.H. Drug Resistance in Leishmaniasis. Clin. Microbiol. Rev. 2006, 19, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, S.; Guerin, P.; Caljon, G.; Croft, S.L.; Maes, L. Evaluating drug resistance in visceral leishmaniasis: The challenges. Parasitology 2016, 145, 453–463. [Google Scholar] [CrossRef]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.-C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl. Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef] [PubMed]
- Roatt, B.M.; de Oliveira Cardoso, J.M.; De Brito, R.C.F.; Coura-Vital, W.; de Oliveira Aguiar-Soares, R.D.; Reis, A.B. Recent advances and new strategies on leishmaniasis treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef] [PubMed]
- Adaui, V.; Schnorbusch, K.; Zimic, M.; Gutiérrez, A.; Decuypere, S.; Vanaerschot, M.; DE Doncker, S.; Maes, I.; Llanos-Cuentas, A.; Chappuis, F.; et al. Comparison of gene expression patterns among Leishmania braziliensis clinical isolates showing a different in vitro susceptibility to pentavalent antimony. Parasitology 2010, 138, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Equbal, A.; Suman, S.S.; Anwar, S.; Singh, K.P.; Zaidi, A.; Sardar, A.H.; Das, P.; Ali, V. Stage-Dependent Expression and Up-Regulation of Trypanothione Synthetase in Amphotericin B Resistant Leishmania donovani. PLoS ONE 2014, 9, e97600. [Google Scholar] [CrossRef] [Green Version]
- Pérez, V.G.; García-Hernandez, R.; Corpas-López, V.; Tomas, A.; Martín-Sanchez, J.; Castanys, S.; Gamarro, F. Decreased antimony uptake and overexpression of genes of thiol metabolism are associated with drug resistance in a canine isolate of Leishmania infantum. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Kulshrestha, A.; Sharma, V.; Singh, R.; Salotra, P. Comparative transcript expression analysis of miltefosine-sensitive and miltefosine-resistant Leishmania donovani. Parasitol. Res. 2014, 113, 1171–1184. [Google Scholar] [CrossRef]
- Mittal, M.; Sundar, S.; Rai, S.; Goyal, N.; Ravinder, A.; Gupta, S. Characterization of natural antimony resistance in Leishmania donovani isolates. Am. J. Trop. Med. Hyg. 2007, 76, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Padmanabhan, P.K.; Singh, S.; Roy, G.; Girard, I.; Chatterjee, M.; Ouellette, M.; Madhubala, R. Role of ABC transporter MRPA, -glutamylcysteine synthetase and ornithine decarboxylase in natural antimony-resistant isolates of Leishmania donovani. J. Antimicrob. Chemother. 2006, 59, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, R.; Dey, S.; Xu, N.; Gage, D.; Lightbody, J.; Ouellette, M.; Rosen, B.P. Trypanothione overproduction and resistance to antimonials and arsenicals in Leishmania. Proc. Natl. Acad. Sci. USA 1996, 93, 10383–10387. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Roberts, S.; Ullman, B.; Madhubala, R. A quantitative proteomic screen to identify potential drug resistance mechanism in α-difluoromethylornithine (DFMO) resistant Leishmania donovani. J. Proteom. 2014, 102, 44–59. [Google Scholar] [CrossRef]
- Walker, J.; Gongora, R.; Vasquez, J.-J.; Drummelsmith, J.; Burchmore, R.; Roy, G.; Ouellette, M.; Gomez, M.A.; Saravia, N.G. Discovery of factors linked to antimony resistance in Leishmania panamensis through differential proteome analysis. Mol. Biochem. Parasitol. 2012, 183, 166–176. [Google Scholar] [CrossRef]
- Wyllie, S.; Mandal, G.; Singh, N.; Sundar, S.; Fairlamb, A.H.; Chatterjee, M. Elevated levels of tryparedoxin peroxidase in antimony unresponsive Leishmania donovani field isolates. Mol. Biochem. Parasitol. 2010, 173, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Wyllie, S.; Vickers, T.J.; Fairlamb, A.H. Roles of Trypanothione S -Transferase and Tryparedoxin Peroxidase in Resistance to Antimonials. Antimicrob. Agents Chemother. 2008, 52, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Goyeneche-Patino, D.A.; Valderrama, L.; Walker, J.; Saravia, N.G. Antimony Resistance and Trypanothione in Experimentally Selected and Clinical Strains of Leishmania panamensis. Antimicrob. Agents Chemother. 2008, 52, 4503–4506. [Google Scholar] [CrossRef] [Green Version]
- Mohebali, M.; Kazemirad, E.; Hajjaran, H.; Kazemirad, E.; Oshaghi, M.A.; Raoofian, R.; Teimouri, A. Gene expression analysis of antimony resistance in Leishmania tropica using quantitative real-time PCR focused on genes involved in trypanothione metabolism and drug transport. Arch. Dermatol. Res. 2019, 311, 9–17. [Google Scholar] [CrossRef]
- Decuypere, S.; Rijal, S.; Yardley, V.; De Doncker, S.; Laurent, T.; Khanal, B.; Chappuis, F.; Dujardin, J.-C. Gene Expression Analysis of the Mechanism of Natural Sb(V) Resistance in Leishmania donovani Isolates from Nepal. Antimicrob. Agents Chemother. 2005, 49, 4616–4621. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, M.S.; Comini, M.A.; Resende, B.V.; Santi, A.M.M.; Zoboli, A.P.; Moreira, D.S.; Murta, S.M. Ornithine decarboxylase or gamma-glutamylcysteine synthetase overexpression protects Leishmania (Vianna) guyanensis against antimony. Exp. Parasitol. 2017, 175, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Haimeur, A.; Guimond, C.; Pilote, S.; Mukhopadhyay, R.; Rosen, B.P.; Poulin, R.; Ouellette, M. Elevated levels of polyamines and trypanothione resulting from overexpression of the ornithine decarboxylase gene in arsenite-resistant Leishmania. Mol. Microbiol. 1999, 34, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Canuto, G.A.B.; Castilho-Martins, E.A.; Tavares, M.F.M.; Rivas, L.; Barbas, C.; López-Gonzálvez, Á. Multi-analytical platform metabolomic approach to study miltefosine mechanism of action and resistance in Leishmania. Anal. Bioanal. Chem. 2014, 406, 3459–3476. [Google Scholar] [CrossRef]
- Basselin, M.; Badet-Denisot, M.-A.; Lawrence, F.; Gero, M.R. Effects of Pentamidine on Polyamine Level and Biosynthesis in Wild-Type, Pentamidine-Treated, and Pentamidine-Resistant Leishmania. Exp. Parasitol. 1997, 85, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Basselin, M.; Lawrence, F.; Robert-Gero, M. Altered transport properties of pentamidine-resistant Leishmania donovani and L. amazonensis promastigotes. Parasitol. Res. 1997, 83, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Coons, T.; Hanson, S.; Bitonti, A.; Mccann, P.; Ullman, B. Alpha-difluoromethylornithine resistance in Leishmania donovani is associated with increased ornithine decarboxylase activity. Mol. Biochem. Parasitol. 1990, 39, 77–89. [Google Scholar] [CrossRef]
- D’Antonio, E.L.; Ullman, B.; Roberts, S.C.; Dixit, U.G.; Wilson, M.E.; Hai, Y.; Christianson, D.W. Crystal structure of arginase from Leishmania mexicana and implications for the inhibition of polyamine biosynthesis in parasitic infections. Arch. Biochem. Biophys. 2013, 535, 163–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hai, Y.; Christianson, D.W. Crystal structures of Leishmania mexicana arginase complexed with α,α-disubstituted boronic amino-acid inhibitors. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2016, 72, 300–306. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. donovani | L. major | L. braziliensis | T. brucei | Mus musculus | Homo sapiens | |
|---|---|---|---|---|---|---|
| L. donovani | 100% | 91.5% | 64.2% | 24.5% | 25.1% | 23.9% |
| L. major | 100% | 63.6% | 24.1% | 24.7% | 23.6% | |
| L. braziliensis | 100% | 26.3% | 25.3% | 25.9% | ||
| T. brucei | 100% | 58.6% | 59.7% | |||
| Mus musculus | 100% | 90.7% | ||||
| Homo sapiens | 100% |
| Leishmania Species | Inhibition of Parasites Growth In Vitro | Reference |
|---|---|---|
| L. donovani | EC50 40 µM in axenic amastigotes; DFMO protected macrophages from infection | [107] |
| No growth inhibition of promastigotes, but EC50 50 µM in intracellular amastigotes | [59] | |
| 5 mM effectively inhibited promastigote growth | [138] | |
| EC50 30 µM in promastigotes | [58] | |
| EC50 125 µM in promastigotes | [137] | |
| L. infantum | EC50 38 µM in promastigotes | [133] |
| L. mexicana | No effect in promastigotes | [138] |
| 10 mM DFMO suppressed promastigote growth after seven passages or 28 days of growth | [76] | |
| 5 mM DFMO suppressed promastigote growth after three passages or 10 days of growth | [134] | |
| L. major | No effect in promastigotes | [138] |
| L. braziliensis guyanensis | 5 mM effectively inhibited promastigote growth | [138] |
| Leishmania Species | Type of Rodent | Amount, Administration, and Efficacy | References |
|---|---|---|---|
| L. donovani | BALB/c mice | 2% in drinking water for 3 weeks reduced liver parasite burden by 93% but had no effect on parasite numbers in the spleen | [111] |
| BALB/c mice | 1% and 3% in drinking water for 7 days suppressed liver burden by 16% and 53%, respectively | [138] | |
| Golden Hamster | 2% in drinking water 2 days after infection and continued for 4 days reduced infection in liver and spleen by 90% and 99%, respectively | [140] | |
| L. infantum | BALB/c mice | 100 mg/kg subcutaneous for 5 days and 200 mg/kg subcutaneous for 42 days reduced infection in liver by 85% and 98%, respectively | [139] |
| L. mexicana | BALB/c mice | 2% and 4% in drinking water for 3 weeks reduced infections by 12% and 20% | [138] |
| L. braziliensis guyanensis | BALB/c mice | 2, 4, and 5% in drinking water reduced lesion size by 100%, 43%, and 81%, respectively | [138] |
| Compound | Inhibition of Recombinant L. donovani ODC (Ki) | Inhibition of Promastigotes (IC50) | Inhibition of Intracellular Amastigotes (IC50) in J774A.1Macrophages | References |
|---|---|---|---|---|
| APA | 1.0 nM | 42 ± 8 µM | 5 ± 2.0 µM | [59,118,141] |
| GAPA | 60 µM | 36 ± 7.0 µM | 9 ± 1.0 µM | [141,142] |
| L. donovani | L. major | L. braziliensis | T. brucei | T. cruzi | Mus musculus | Homo sapiens | |
|---|---|---|---|---|---|---|---|
| L. donovani | 100% | 97.6% | 88.5% | 61.8% | 70.9% | 26.2% | 26.5% |
| L. major | 100% | 88.5% | 61.9% | 70.4% | 26.2% | 26.5% | |
| L. braziliensis | 100% | 61.9% | 70.1% | 27.0% | 27.0% | ||
| T. brucei | 100% | 68.3% | 26.8% | 26.9% | |||
| T. cruzi | 100% | 25.9% | 25.9% | ||||
| Mus musculus | 100% | 98.2% | |||||
| Homo sapiens | 100% |
| L. donovani | L. major | T. brucei | T. cruzi | |
|---|---|---|---|---|
| L. donovani | 100% | 89% | 40.8% | 40.2% |
| L. major | 100% | 40.4% | 41.9% | |
| T. brucei | 100% | 48.5% | ||
| T. cruzi | 100% |
| L. donovani ADOMETDC versus L. donovani Prozyme | L. major ADOMETDC versus L. major Prozyme | T. brucei ADOMETDC versus T. brucei Prozyme | T. cruzi ADOMETDC versus T. cruzi Prozyme |
|---|---|---|---|
| 24.4% | 23.1% | 23.2% | 26.3% |
| L. donovani | L. major | L. braziliensis | T. cruzi | T. brucei | Mus musculus | Homo sapiens | |
|---|---|---|---|---|---|---|---|
| L. donovani | 100% | 96.7% | 89.0% | 65.6% | 67.0% | 45.6% | 45.4% |
| L. major | 100% | 88.7% | 65.9% | 66.0% | 44.3% | 44.1% | |
| L. braziliensis | 100% | 65.9% | 65.3% | 44.3% | 42.8% | ||
| T. cruzi | 100% | 71.8% | 43.1% | 42.3% | |||
| T. brucei | 100% | 44.6% | 43.8% | ||||
| Mus musculus | 100% | 94.7% | |||||
| Homo sapiens | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carter, N.S.; Kawasaki, Y.; Nahata, S.S.; Elikaee, S.; Rajab, S.; Salam, L.; Alabdulal, M.Y.; Broessel, K.K.; Foroghi, F.; Abbas, A.; et al. Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target. Med. Sci. 2022, 10, 24. https://doi.org/10.3390/medsci10020024
Carter NS, Kawasaki Y, Nahata SS, Elikaee S, Rajab S, Salam L, Alabdulal MY, Broessel KK, Foroghi F, Abbas A, et al. Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target. Medical Sciences. 2022; 10(2):24. https://doi.org/10.3390/medsci10020024
Chicago/Turabian StyleCarter, Nicola S., Yumena Kawasaki, Surbhi S. Nahata, Samira Elikaee, Sara Rajab, Leena Salam, Mohammed Y. Alabdulal, Kelli K. Broessel, Forogh Foroghi, Alyaa Abbas, and et al. 2022. "Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target" Medical Sciences 10, no. 2: 24. https://doi.org/10.3390/medsci10020024
APA StyleCarter, N. S., Kawasaki, Y., Nahata, S. S., Elikaee, S., Rajab, S., Salam, L., Alabdulal, M. Y., Broessel, K. K., Foroghi, F., Abbas, A., Poormohamadian, R., & Roberts, S. C. (2022). Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target. Medical Sciences, 10(2), 24. https://doi.org/10.3390/medsci10020024