
An Overview of Malaria Transmission Mechanisms, Control, and Modeling


1
Center for Development Research ZEF, University of Bonn, Genscherallee 3, 53113 Bonn, Germany
2
T.H. Chan Harvard School of Public Health, Boston, MA 02115, USA
Med. Sci. 2023, 11(1), 3; https://doi.org/10.3390/medsci11010003
Submission received: 1 December 2022
/
Revised: 11 December 2022
/
Accepted: 19 December 2022
/
Published: 23 December 2022
(This article belongs to the Section Immunology and Infectious Diseases)
{kind=link}
{kind=link}
{kind=link}
Abstract
:In sub-Saharan Africa, malaria is a leading cause of mortality and morbidity. As a result of the interplay between many factors, the control of this disease can be challenging. However, few studies have demonstrated malaria’s complexity, control, and modeling although this perspective could lead to effective policy recommendations. This paper aims to be a didactic material providing the reader with an overview of malaria. More importantly, using a system approach lens, we intend to highlight the debated topics and the multifaceted thematic aspects of malaria transmission mechanisms, while showing the control approaches used as well as the model supporting the dynamics of malaria. As there is a large amount of information on each subject, we have attempted to provide a basic understanding of malaria that needs to be further developed. Nevertheless, this study illustrates the importance of using a multidisciplinary approach to designing next-generation malaria control policies.
1. Introduction
Approximately 2000 children die daily from malaria, with 90% of the victims located in sub-Saharan Africa (SSA) [1,2]. Transmission and control of malaria are mediated by complex interactions and feedback loops among humans, mosquitoes, parasites, their environments, healthcare systems, and policy implementation at a given period of time [3]. Therefore, malaria transmission can be described as complex, nonlinear, and dynamic. The endemism of malaria in West Africa, for example, has huge economic impacts [2,4]. In Ghana, the cost of treating a single episode of malaria can reach 34% of a household’s yearly income [5]. As a result of infected people being unable to produce wealth, and as wealth is low, the risk of exposure to malaria increases. There is a negative relationship between national economic growth and malaria control expenditures [2,6]. As a result, malaria can be considered both as a cause of poverty and as a consequence of it [7].
Furthermore, past interventions have been ineffective as they were unable to account for the nonlinear nature of malaria infection. Particularly, the unexpected consequences of interventions such as mosquito resistance to chemical compounds and parasite resistance to drugs led to their abandonment. This was the case with the eradication control program that was abandoned in 1969 and the Global Malaria Control Strategy in 1992 [8]. Scientists have widely acknowledged the complexity of malaria, but a depiction of this complexity is weakly documented, and further limits the scope of malaria control.
This short review embracing several topics or thematic areas provides the reader with a piece of basic knowledge of malaria transmission, control, and modeling while illustrating its complexity. Therefore, the manuscript intends to be a didactical material, providing a brief overview of the disease, reducing information asymmetry, while arousing the curiosity of readers who can refer to more specific literature.
2. Materials and Methods
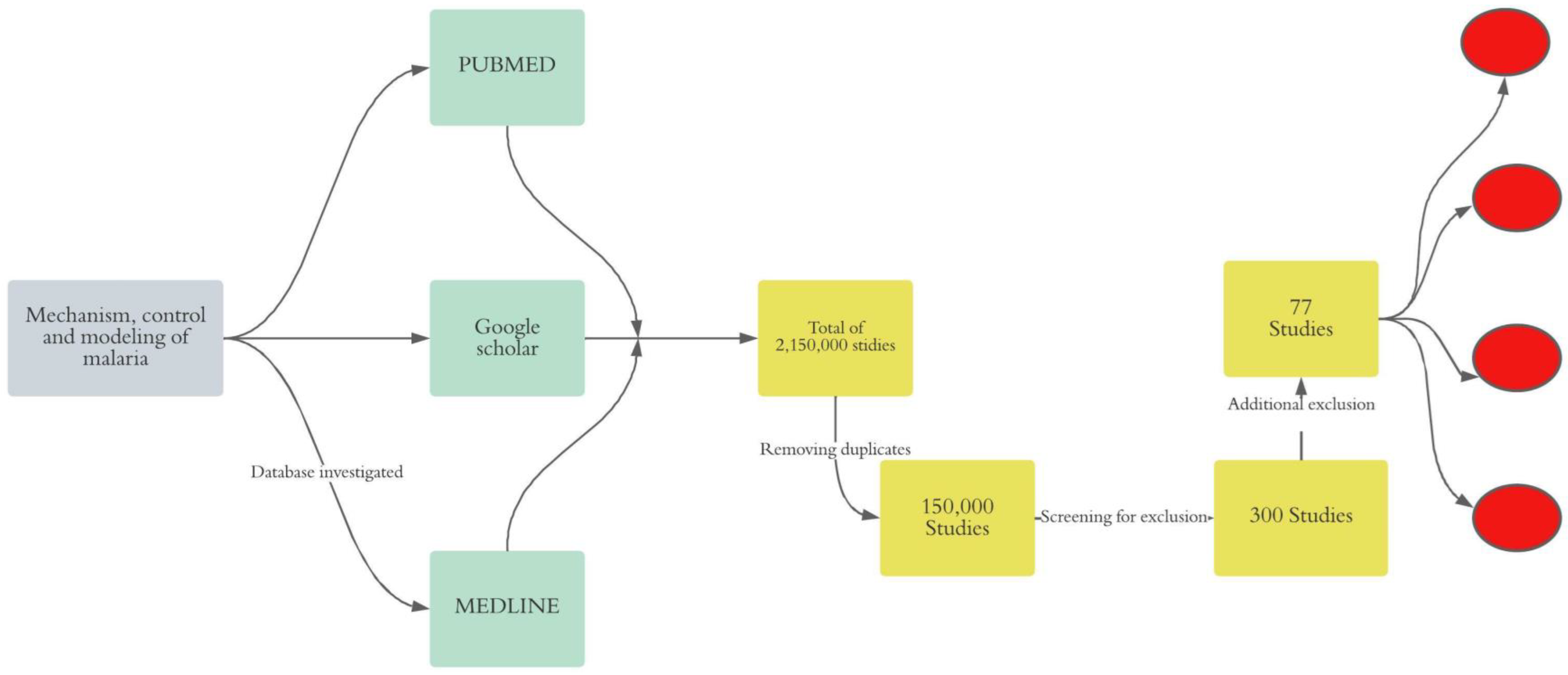
We conducted an unsystematic narrative review of online literature to gain a deeper insight into malaria transmission mechanism, control strategy, and modeling approaches. Thus, we adopted the following approach (Figure 1).
2.1. Inclusion and Exclusion Criteria for Paper Eligibility
Our search for peer-reviewed papers on malaria in the online scientific database was restricted to the period from 1950 to 2020. The rationale for this time window lies in the fact that the first malaria model was developed in the 1950s and since then, most of the recommendations for malaria control are supported by quantitative analysis. Besides, we excluded all duplicated letters, narrative reviews, case reports, and commentary. Given the tremendous research done on malaria, we limited the scope of the study to only peer-reviewed published papers in either French or English. This was to avoid evidence that may be challenging to replicate and biased translation. Within the topic of interest, we exclude all case reports which happen in non-endemic areas.
2.2. Search Strategy
We searched three databases PUBMED, Google Scholar, and MEDLINE for relevant literature on malaria from 1950 to 2020.
2.3. Screening of Studies and Evidence Extraction
Our screening process focuses on the title of the study published in peer-reviewed journals. Our next step was to review the papers’ abstracts and remove the studies that did not fit under the umbrella of the keywords.
2.4. Analysis and Gaps in the Analysis
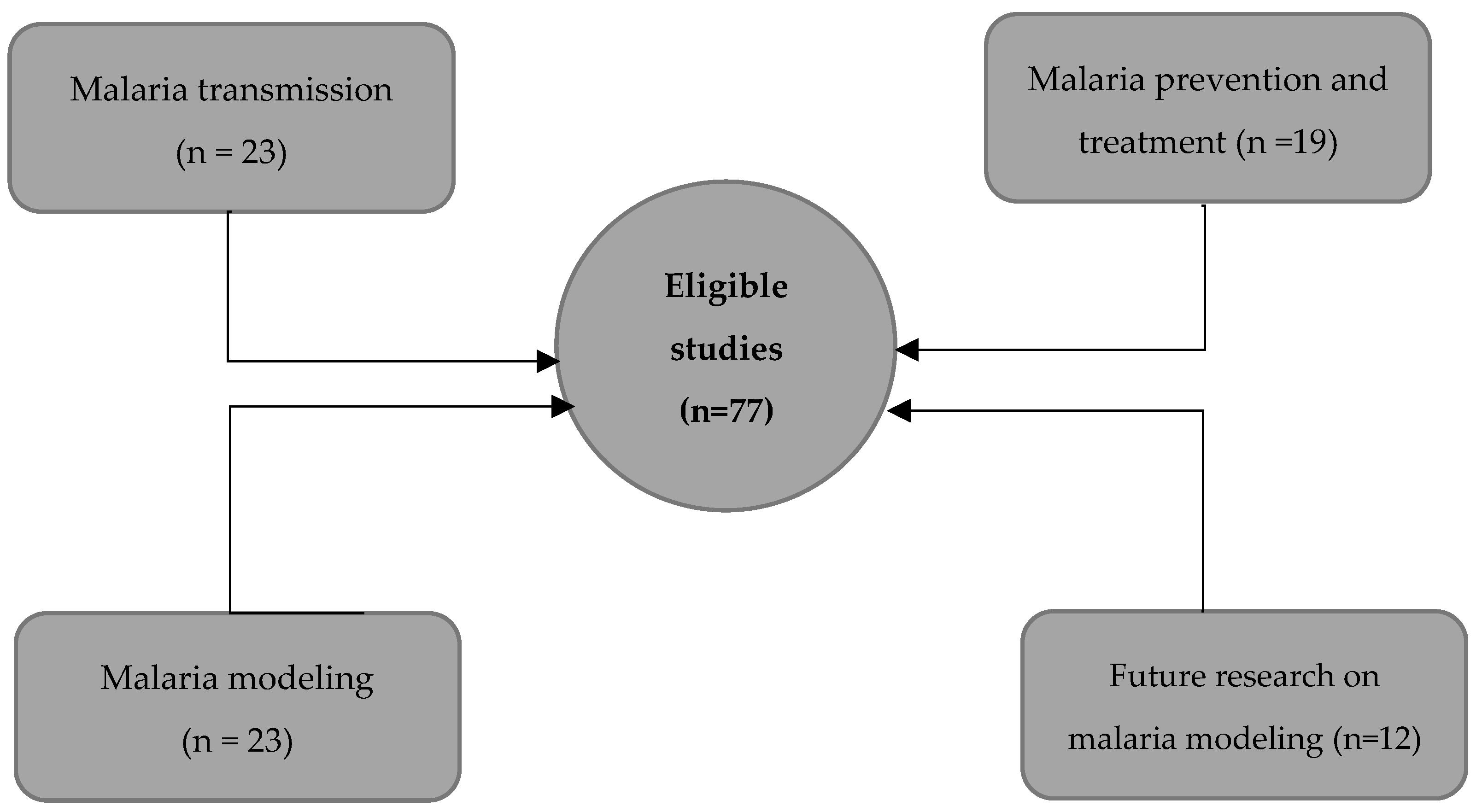
To get a comprehensive understanding of malaria, we categorized the list of 77 eligible peer-reviewed papers into topics of interest, such as mechanisms, prevention and treatment, mathematical modeling, and perspectives on modeling. The topical organization of the collected papers allowed us to get a broad perspective of the state of the art.
3. Results
We found 2,150,000 studies after the search. A total of 150,000 studies were obtained after removing duplicates. We included only 300 studies after screening both titles and abstracts. Furthermore, we removed 50 case reports, 146 studies in malaria non-endemic areas, and 16 reports including 10 thesis and 6 NGO reports.
Using the remaining 77 eligible studies, we provided an overview of each topic and complemented our narrative with WHO, CDC reports, and 10 complimentary readings. There were 23 studies supporting malaria transmission, 19 on prevention and treatment, 23 on mathematical modeling, and 12 on future research on malaria modeling (Figure 2).
3.1. Malaria Transmission Mechanism
Malaria infection is caused by interactions between humans, mosquitoes, and the pathogen (Figure 3). Female mosquitoes change during the egg-laying period when they experience a deficiency in protein and plant sucrose necessary for egg maturation. Consequently, they blood-feed on humans and can infect them after effective biting where infectious protozoans (of the genus Plasmodium) are injected into the human bloodstream [9,10].
Female mosquitoes most found in West Africa belong to the Anopheles Giles genus [11]. Anopheles gambiae (sensu stricto) prefers to use human blood (anthropophilic) to complete its gonotrophic cycle, bites primarily indoors (endophagic), and rests indoors (endophilic). As far as its feeding habits are concerned, An. gambiae is the major malaria vector in West Africa [12]. In addition, there are other vectors, such as An. arabiensis, An. funestus and An. melas able to carry P. falciparum. Similar to An. gambiae, An. funestus is anthropophilic, endophagic, and endophilic [4,13]. On the other hand, An. arabiensis is zoophilic (prefers blood-feeding animals), exophilic (bites outdoors), and exophagic (rests outdoors) [11]. An. melas is also anthropophagic, exophagic, and exophilic [12]. Therefore, P. falciparum adapts well to Anopheles species, increasing malarial vulnerability in the host. The wide range of vector conditions, feeding habits, and geographical heterogeneity in malaria transmission make malaria modeling even more challenging [14].
A mosquito’s ability to adapt to environmental conditions such as humidity and temperature, is influenced by its sensitivity to those conditions. This adaptation can take the form of a change in the life cycle length, or the biting efficiency [15,16,17,18,19]. As a long-term study carried out in Kenya (1999–2010) has shown, vectors’ feeding habits and effectiveness can change over time [20].
The transmission intensity may differ significantly across host populations because vectors’ attraction to the host varies [21]. This variation is elicited by the difference in human bodies’ microflora and their reaction to mosquito bites [22]. For instance, an infected host is more attractive to other mosquitoes than a non-infected one [23]. Likewise, mosquitoes prefer the scent of children to that of the adults [21,24].
In SSA, infectious protozoans are, Plasmodium vivax, P. ovale, P. malariae, P. knowlesi, and P. falciparum where P. falciparum accounts for 95% of malaria cases [25,26]. P. falciparum’s complete life cycle consists of an incubation period of 10 days in the vector and a period of 7 to 20 days in the human host [27]. When P. falciparum protozoans are injected into a host’s bloodstream via the vector saliva, the protozoan completes its life cycle. After entering the host bloodstream, the sporozoites (asexual protozoans) move into the liver, where they penetrate the hepatocytes (liver cells). In the host hepatocytes, the parasite divides asexually several times, a process known as schizogony [28]. Eventually, the hepatocytes break, releasing merozoites which infest red blood cells in the host bloodstream. The parasites can diverge into gametocytes after repeated divisions of the red blood cells [29]. Malaria can cause symptoms such as fever, chills, and sweat before and during the differentiation of P. falciparum into sexual stages. Even so, some hosts may not exhibit clinical symptoms and are considered asymptomatic [30]. Whenever mosquitoes bite infected people (symptomatic and asymptomatic), they primarily suck out the gametocytes [29]. Detecting asymptomatic individuals in endemic areas can be challenging, and mass testing may not always be cost-effective in detecting such individuals [31].
3.2. Malaria Prevention and Treatment
Due to recursive interactions between humans, mosquitoes, and parasites that cause malaria transmission, existing control methods might not be effective in the sense that they target either mosquitoes or parasites. Mosquitoes have been controlled mainly using the obliteration of larvae and adult stages while reducing the entomological inoculation rate. These controls include physical, biological, or chemical changes to the vector’s environment. As part of the physical modification, breeding sites are removed, and the sources of larvae are managed using drainage and weeding [32]. However, this method requires a constant working force needed for weeding and drain cleaning which led SSA governments to abandon it although it is among the most effective [33]. Chemical modification of larval environments includes the application of larvicides and insecticides. The downside of this method is that it is also costly, and mosquitoes often become resistant to insecticides [34]. Natural enemies of mosquito larvae, such as larvivorous fish and bacteria (e.g., Bacillus thuringiensis), are used in the biological method. Nevertheless, this process is hindered by the temporary persistence of natural enemies [32].
The Global Malaria Action Plan (GMAP) and the World Health Organization (WHO) recommended the use of indoor residual spray (IRS) and insecticide-treated bed nets (ITNs) for controlling mosquito adult populations in the 2010s across Africa [35]. The main chemical component of ITNs and IRS is synthetic pyrethroid, a lethal compound, that repels mosquitoes, remains in the environment, and is harmless to mammals. From 2005 to 2010, malaria cases in Ghana declined by 41% due to the successful use of ITNs and IRS [36]. The residual effects of synthetic pyrethroids in the environment resulting from IRS, ITNs, and agricultural pest management force mosquitoes to develop coping mechanisms [37]. These include indoor and outdoor spray avoidance mechanisms, earlier biting, and insecticide resistance [38].
The control of malaria parasites is achieved through three lines of defense: antimalarial drugs, seasonal prevention, and vaccination. The first line of the curative measures adopted by GMAP is the Artemisinin-based Combination Therapies (ACT). ACTs are highly potent against P. falciparum with a higher clearance rate and symptoms resorption than other curative therapies [39]. Nevertheless, P. falciparum has progressively developed ACT resistance [40]. Malaria parasite resistance in SSA is exacerbated by using counterfeit malaria drugs [41,42]. Another preventive measure is intermittent prevention, which involves administering single-dose malaria therapy to pregnant women [43]. The treatment includes the combination of primaquine, sulfadoxine-pyrimethamine, amodiaquine, methylene blue, and dihydroartemisinin-piperaquine which have been proven to be able to prevent the transmission of P. falciparum [44]. A seasonal vaccine “RTS, S/AS01” is proven effective for children under 5 years old for a short period with an efficacy rate of 82% [45,46]. The authors demonstrated that RTS, S/AS01 induces an immune response to the proteins of infected sporozoites. Vaccine components protect against clinical malaria. The effectiveness of its protective action can wane over time and with advancing age. Particularly, children between the ages of 6 to 12 weeks benefit from the vaccine more than children between the ages of 5 to 17 months.
A less explored line of malaria treatment consists of phytotherapy. For example, a study on phytochemicals identified more than 20 local plants and herbs in central Africa that can heal and prevent malaria transmission with almost zero risks of P. falciparum developing resistance against these natural compounds [47,48,49]. According to the authors, the synergistic interplay between a wide range of chemical substances in the plants enables treating malaria holistically while preventing the development of resistance [48,50].
3.3. Mathematical Modeling of Malaria
Mathematical models developed to gain insight into disease transmission have influenced past and present interventions to prevent or treat diseases. Mathematical modeling of malaria transmission began in 1911 with the Susceptible-Infectious-Removed model (SIR), which compartmentalized the population of hosts and vectors into three groups [51,52]. The compartments were denoted susceptible (S), referring to the population likely to become infected, infectious (I), composed of the fraction of the infected population, and removed/ recovered (R), accounting for the fraction of the population that either died or healed from the disease. Several assumptions were assumed in the SIR model, including a closed and finite population, a homogeneous mosquito bite rate, and a well-mixed population. Although the Ross model (SIR) was unable to adjust to the new incidence data due to its limited predictability, SIR provides insights into the intricate relationship between infected hosts and mosquito densities. Because Ross’s model was theoretical and simulation-based, it failed to document proof for campaigns for mosquito eradication.
Subsequently, George Macdonald complemented the SIR model [51] by feeding the model with real data and embedding an additional compartment for the latency period between the mosquito bite and the onset of symptoms denoted Exposed (E). Additionally, Macdonald’s study argued for eradicating mosquitoes through a massive campaign based on the theory of superinfection [53]. Consequently, between 1955 and 1969, WHO implemented widespread and rigorous mosquito eradication campaigns as part of the Global Malaria Eradication Program (GMEP). In these campaigns, the pesticide Dichlorodiphenyltrichloroethane (DDT) was successfully used to control malaria in major European and American countries [54]. The GMEP, however, failed to eradicate malaria worldwide due to mosquito resistance [55]. Apart from mosquito insecticide resistance, the absence of basic healthcare services, high malaria transmission intensity, and other socio-ecological factors hindered the success of eradication campaigns in the SSA [56].
The mathematical epidemiology of malaria has evolved steadily in recent decades from “toy models” (which were not realistic but captured the key features of the disease) to “high-level models” (which are precise and sacrifice generality) [57]. As such, complex models display common features such as the interaction between many components [58]. For example, an earlier partial differential model split the infected population into age-dependent infections in the SIR framework [59].
An important metric that summarizes the transmission of a disease is the reproductive number (R0). R0 represents the expected number of infected human hosts after effective mosquito bites in a fully susceptible population [60,61,62]. Thus, R0 provides a measurement for the intensity of transmission and contributes to the definition of disease-endemic areas when it is greater than one (R0 > 1) [63]. Heterogeneity was therefore incorporated in populations for the computation of R0. These include, for example, the age structure of the host [64], migration of the vectors and hosts [65], host beliefs and practices [66], and host income classes [67,68]. R0 also varies with the degree of complexity introduced into the compartmentalization of the population [66]. The more the modeling framework embodies the heterogeneity in the host population, the closer to real-life transmission dynamics the value obtained for R0 becomes [63,68,69,70]. The pioneering models that provided insight into malaria dynamics and its controls were unrealistic for various reasons, and maximum control has hardly been achieved in [56]. Most specifically, a large R0 indicates a higher density of mosquitoes that reduces the effectiveness of ITN in a large and likely heterogeneous host population [63].
Many mathematical models were developed combining the compartments S, E, I, and R (e.g., SIR, SIS, SI, SIRS, SEIR, SEIRS, SEI) and included the heterogeneity of the population (meta-population modeling approaches) [69]. Most of these models do not capture the dynamics of subpopulations because they tend to assume large and homogeneous populations that do not take into account the existence and specific characteristics of subpopulations [71,72]. Another issue is that they fail to account for host behavior, which influences the disease’s dynamics and control [73,74]. Besides, compartmental models are either knowledge or data-driven (e.g., Ross model, Macdonald models). The models cannot address the complex interactions between determinants that guide the disease dynamics in both cases. For example, socioeconomic determinants interfere with the parameters generally considered in the calculation of R0 such as mortality, mobility, and birth rate [66]. Thus, models that capture, realistically, the transmission process of malaria are still missing.
3.4. Malaria Modeling: Present and Future
The ability to track epidemics became more accurate with the advancement in computing power and the availability of data. The deterministic framework uses in this context provides insight into disease dynamics, but usually fails to take uncertainties into account. Stochastic models, on the other hand, incorporate randomness that may occur during the epidemic and, most importantly, accounts for a large range of uncertainty that occurs during epidemics. For example, a stochastic model was used to predict the complex transmission pattern concerning environmental changes [75]. Like deterministic models, the stochastic models help to disclose interesting features of the transmission such as the impact of the network structure, and the properties of an outbreak most specifically get insights into the rate of the surge of vector-borne diseases in a new region in a context of climate change [76,77]. Furthermore, both stochastic and deterministic models are population-based and often fail to consider the feature of individuals which alternatively the individual-based models (IBMs) do [78]. IBMs can account for heterogeneity across individual agents and spatial gradients [79,80]. However, parametrizing them remains a daunting task, especially due to the lack and accuracy of data [81]. Beyond meta-population modeling (Ross-based models), within-host modeling opened new possibilities to address malaria infectiousness while examining the interplay between hosts’ immune systems and the dynamics of the parasite. However, modeling the superinfection of malaria while tracking each parasite strain and incorporating the diverse genetic makeup of the parasites remains extremely challenging, especially in terms of the model parameterization [82,83]. Most recently, the introduction of a new type of data such as human mobility collected from data providers (Facebook, Twitter, Google, etc.) refined the field of epidemiological surveillance. These data have been used to pinpoint malaria hotspots and predict their spread [84,85,86]. The use of fine-grained mobility data opens a new prospect of modeling and documenting evidence for policy recommendations; however, privacy concerns need to be addressed [87].
4. Discussion
Malaria remains a major health challenge in Sub-Saharan Africa, despite extensive research. Several factors contribute to malaria transmission, including socio-ecological factors and changes in the human host and vector behavior, which may be channeled by parasite genetic changes. Considering malaria’s complex nature, modeling it presents numerous challenges. As an example, it can be difficult to accurately parameterize a model with a compartment of asymptomatic individuals as there is a lack of reliable historical data. However, evidence suggests that malaria symptoms could progress rapidly from none to life-threatening while leading to a lifetime burden of cognitive impairments for some patients. It is thus necessary to develop a strategy that allows for the production at a low cost of reliable primary data on asymptomatic people with the capability of identifying infectious Plasmodium [88]. Additionally, the review highlighted that for each of the thematic areas discussed, there is a web of determinants that can be studied, using different perspectives including the use of a multidisciplinary lens to examine clinically relevant malaria issues.
A historical approach was used to illustrate the evolution of the malaria model from simplistic to more elaborate models. Moreover, this review outlines the challenges of modeling, where, for instance, introducing mobile data to a mathematical framework may pose a problem of re-identification. The number and scale of parameters involved in a complex modeling approach make model parameterization challenging.
Finally, this review demonstrates that malaria can efficiently be controlled by combining several strategies. Among them are constant awareness and education campaigns in communities at risk and among individuals whose resistance to non-pharmaceutical interventions (such as ITNs) can hinder malaria control efforts.
Given the wide range of thematic areas discussed, we acknowledge that we have only scratched the surface of the various cross-cutting topics. Consequently, we did not elaborate on the host immune response to malaria nor on technological advances in diagnosing and treating malaria since they are widely covered elsewhere [89,90,91,92,93,94]. Additionally, this paper faces two challenges, namely (1) a self-selection bias that might lead to a more prominent emphasis on some cross-cutting topics, and (2) not considering recently published papers [95]. Nonetheless, this review provides readers with an overview of malaria from a global perspective.
Funding
This research received no external funding.
Institutional Review Board Statement
This study was approved by the Center for Development Research (ZEF) ethics committees, University of Bonn.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are generated from the content of publications mentioned in this manuscript and they are equally available from the corresponding authors on reasonable request.
Acknowledgments
The author wants to thank Christian Borgemeister, Daniel Callo-Concha, Lauren Childs, Deguenon Jean Marcel, Henri Tonnang, and Joshua Ntajal for their edits, comments, and suggestions. In addition, we would like to extend our gratitude to the Center for Development Research (ZEF), Germany, for providing us with the logistics required for this review.
Conflicts of Interest
The author declares no competing interests.
Abbreviations
| ACT | Artemisinin-based Combination Therapies |
| DDT | Dichlorodiphenyltrichloroethane |
| GMAP | Global Malaria Action Plan |
| GMEP | Global Malaria Eradication Program |
| IBM | Individual-based models |
| IRS | Indoor residual spraying |
| ITN | Insecticide-treated bed nets |
| R0 | reproductive number |
| SSA | Sub-Saharan Africa |
| SIR | Susceptible Infectious Removed model |
| WHO | World Health Organization |
References
- Ndugwa, R.P.; Ramroth, H.; Müller, O.; Jasseh, M.; Sié, A.; Kouyaté, B.; Greenwood, B.; Becher, H. Comparison of All-Cause and Malaria-Specific Mortality from Two West African Countries with Different Malaria Transmission Patterns. Malar. J. 2008, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, J.; Malaney, P. The Economic and Social Burden of Malaria. Nature 2002, 415, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Savi, M.K.; Callo-Concha, D.; Tonnang, H.E.Z.; Borgemeister, C. Emerging Properties of Malaria Transmission and Persistence in Urban Accra, Ghana: Evidence from a Participatory System Approach. Malar. J. 2021, 20, 321. [Google Scholar] [CrossRef] [PubMed]
- Asare, E.O.; Amekudzi, L.K. Assessing Climate Driven Malaria Variability in Ghana Using a Regional Scale Dynamical Model. Climate 2017, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Akazili, J.; Aikins, M.; Binka, F.N. Malaria Treatment in Northern Ghana: What Is the Treatment Cost per Case to Households? Afr. J. Health Sci. 2007, 14, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Jowett, M.; Miller, N.J. The Financial Burden of Malaria in Tanzania: Implications for Future Government Policy. Int. J. Health Plann. Manag. 2005, 20, 67–84. [Google Scholar] [CrossRef]
- Wells, T.N.C.; Alonso, P.L.; Gutteridge, W.E. New Medicines to Improve Control and Contribute to the Eradication of Malaria. Nat. Rev. Drug. Discov. 2009, 8, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Breman, J. Rolling Back Malaria: Action or Rhetoric? Available online: https://www.semanticscholar.org/author/J.-Breman/144538342 (accessed on 15 November 2022).
- Arifin, S.N.; Zhou, Y.; Davis, G.J.; Gentile, J.E.; Madey, G.R.; Collins, F.H. An Agent-Based Model of the Population Dynamics of Anopheles Gambiae. Malar. J. 2014, 13, 424. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why Do Female Aedes Aegypti (Diptera: Culicidae) Feed Preferentially and Frequently on Human Blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef]
- Li, Z.-X.; Pickett, J.A.; Field, L.M.; Zhou, J.-J. Identification and Expression of Odorant-Binding Proteins of the Malaria-Carrying Mosquitoes Anopheles Gambiae and Anopheles Arabiensis. Arch. Insect. Biochem. Physiol. 2005, 58, 175–189. [Google Scholar] [CrossRef]
- Tuno, N.; Kjaerandsen, J.; Badu, K.; Kruppa, T. Blood-Feeding Behavior of Anopheles Gambiae and Anopheles Melas in Ghana, Western Africa. J. Med. Entomol. 2010, 47, 28–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The Dominant Anopheles Vectors of Human Malaria in Africa, Europe and the Middle East: Occurrence Data, Distribution Maps and Bionomic Précis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, L.M.; Abuelezam, N.N.; Dye, C.; Gupta, S.; Murray, M.B.; Williams, B.G.; Buckee, C.O. Modelling Challenges in Context: Lessons from Malaria, HIV, and Tuberculosis. Epidemics 2015, 10, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eikenberry, S.E.; Gumel, A.B. Mathematical Modeling of Climate Change and Malaria Transmission Dynamics: A Historical Review. J. Math. Biol. 2018, 77, 857–933. [Google Scholar] [CrossRef]
- Adu-Prah, S.; Kofi Tetteh, E. Spatiotemporal Analysis of Climate Variability Impacts on Malaria Prevalence in Ghana. Appl. Geogr. 2015, 60, 266–273. [Google Scholar] [CrossRef]
- White, N.J.; Pukrittayakamee, S.; Hien, T.T.; Faiz, M.A.; Mokuolu, O.A.; Dondorp, A.M. Malaria. Lancet 2014, 383, 723–735. [Google Scholar] [CrossRef]
- Chaturvedi, H.K.; Mahanta, J.; Bajpai, R.C.; Pandey, A. Risk of Malaria among Febrile Patients: Retrospective Analysis of a Hospital-Based Study in an Endemic Area of Northeast India. Int. Health 2014, 6, 144–151. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Paaijmans, K.P.; Johnson, L.R.; Balzer, C.; Ben-Horin, T.; de Moor, E.; McNally, A.; Pawar, S.; Ryan, S.J.; Smith, T.C.; et al. Optimal Temperature for Malaria Transmission Is Dramatically Lower than Previously Predicted. Ecol. Lett. 2013, 16, 22–30. [Google Scholar] [CrossRef]
- Mwangangi, J.M.; Mbogo, C.M.; Orindi, B.O.; Muturi, E.J.; Midega, J.T.; Nzovu, J.; Gatakaa, H.; Githure, J.; Borgemeister, C.; Keating, J.; et al. Shifts in Malaria Vector Species Composition and Transmission Dynamics along the Kenyan Coast over the Past 20 Years. Malar. J. 2013, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Busula, A.O.; Takken, W.; de Boer, J.G.; Mukabana, W.R.; Verhulst, N.O. Variation in Host Preferences of Malaria Mosquitoes Is Mediated by Skin Bacterial Volatiles. Med. Vet. Entomol. 2017, 31, 320–326. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Qiu, Y.T.; Beijleveld, H.; Maliepaard, C.; Knights, D.; Schulz, S.; Berg-Lyons, D.; Lauber, C.L.; Verduijn, W.; Haasnoot, G.W.; et al. Composition of Human Skin Microbiota Affects Attractiveness to Malaria Mosquitoes. PLoS ONE 2011, 6, e28991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria Infection Increases Attractiveness of Humans to Mosquitoes. PLoS Biol. 2005, 3, e298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, P.; Schoone, G.; Schallig, H.; Verhage, D.; Telgt, D.; Eling, W.; Sauerwein, R. Quantification of Plasmodium Falciparum Gametocytes in Differential Stages of Development by Quantitative Nucleic Acid Sequence-Based Amplification. Mol. Biochem. Parasitol. 2004, 137, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Karnad, D.R.; Nor, M.B.M.; Richards, G.A.; Baker, T.; Amin, P. Council of the World Federation of Societies of Intensive and Critical Care Medicine Intensive Care in Severe Malaria: Report from the Task Force on Tropical Diseases by the World Federation of Societies of Intensive and Critical Care Medicine. J. Crit. Care 2018, 43, 356–360. [Google Scholar] [CrossRef]
- Kafy, H.T.; Ismail, B.A.; Mnzava, A.P.; Lines, J.; Abdin, M.S.E.; Eltaher, J.S.; Banaga, A.O.; West, P.; Bradley, J.; Cook, J.; et al. Impact of Insecticide Resistance in Anopheles Arabiensis on Malaria Incidence and Prevalence in Sudan and the Costs of Mitigation. Proc. Natl. Acad. Sci. USA 2017, 114, E11267–E11275. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine (US) Committee for the Study on Malaria Prevention and Control. Malaria: Obstacles and Opportunities; Oaks, S.C., Mitchell, V.S., Pearson, G.W., Carpenter, C.C.J., Eds.; National Academies Press (US): Washington, DC, USA, 1991; ISBN 978-0-309-04527-8. [Google Scholar]
- Yam, X.Y.; Preiser, P.R. Host Immune Evasion Strategies of Malaria Blood Stage Parasite. Mol. BioSyst. 2017, 13, 2498–2508. [Google Scholar] [CrossRef]
- Talman, A.M.; Blagborough, A.M.; Sinden, R.E. A Plasmodium Falciparum Strain Expressing GFP throughout the Parasite’s Life-Cycle. PLoS ONE 2010, 5, e9156. [Google Scholar] [CrossRef] [Green Version]
- Bousema, T.; Drakeley, C. Determinants of Malaria Transmission at the Population Level. Cold. Spring. Harb. Perspect. Med. 2017, 7, a025510. [Google Scholar] [CrossRef]
- Sturrock, H.J.W.; Hsiang, M.S.; Cohen, J.M.; Smith, D.L.; Greenhouse, B.; Bousema, T.; Gosling, R.D. Targeting Asymptomatic Malaria Infections: Active Surveillance in Control and Elimination. PLoS Med. 2013, 10, e1001467. [Google Scholar] [CrossRef]
- Tusting, L.S.; Thwing, J.; Sinclair, D.; Fillinger, U.; Gimnig, J.; Bonner, K.E.; Bottomley, C.; Lindsay, S.W. Mosquito Larval Source Management for Controlling Malaria. Cochrane. Database. Syst. Rev. 2013, 8, CD008923. [Google Scholar] [CrossRef]
- Fillinger, U.; Lindsay, S.W. Larval Source Management for Malaria Control in Africa: Myths and Reality. Malar. J. 2011, 10, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Roll Back Malaria Partnership. The Global Malaria Action Plan. In For a Free Malaria World; WHO: Geneva, Switzerland, 2008; p. 274. [Google Scholar]
- Aregawi, M.; Malm, K.L.; Wahjib, M.; Kofi, O.; Allotey, N.-K.; Yaw, P.N.; Abba-Baffoe, W.; Segbaya, S.; Owusu-Antwi, F.; Kharchi, A.T.; et al. Effect of Anti-Malarial Interventions on Trends of Malaria Cases, Hospital Admissions and Deaths, 2005–2015, Ghana. Malar. J. 2017, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Kudom, A.A.; Anane, L.N.; Afoakwah, R.; Adokoh, C.K. Relating High Insecticide Residues in Larval Breeding Habitats in Urban Residential Areas to the Selection of Pyrethroid Resistance in Anopheles Gambiae s.l. (Diptera: Culicidae) in Akim Oda, Ghana. J. Med. Entomol. 2018, 55, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Moshi, I.R.; Ngowo, H.; Dillip, A.; Msellemu, D.; Madumla, E.P.; Okumu, F.O.; Coetzee, M.; Mnyone, L.L.; Manderson, L. Community Perceptions on Outdoor Malaria Transmission in Kilombero Valley, Southern Tanzania. Malar. J. 2017, 16, 274. [Google Scholar] [CrossRef] [Green Version]
- Eastman, R.T.; Fidock, D.A. Artemisinin-Based Combination Therapies: A Vital Tool in Efforts to Eliminate Malaria. Nat. Rev. Microbiol. 2009, 7, 864–874. [Google Scholar] [CrossRef] [Green Version]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium Falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Yasuoka, J.; Jimba, M.; Levins, R. Application of Loop Analysis for Evaluation of Malaria Control Interventions. Malar. J. 2014, 13, 140. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Allan, E.L.; Mamadu, I.; Hall, Z.; Green, M.D.; Swamidos, I.; Dwivedi, P.; Culzoni, M.J.; Fernandez, F.M.; Garcia, G.; et al. Prevalence of Substandard and Falsified Artemisinin-Based Combination Antimalarial Medicines on Bioko Island, Equatorial Guinea. BMJ Glob. Health 2017, 2, e000409. [Google Scholar] [CrossRef] [Green Version]
- Briand, V.; Cottrell, G.; Massougbodji, A.; Cot, M. Intermittent Preventive Treatment for the Prevention of Malaria during Pregnancy in High Transmission Areas. Malar. J. 2007, 6, 160. [Google Scholar] [CrossRef]
- Dicko, A.; Roh, M.E.; Diawara, H.; Mahamar, A.; Soumare, H.M.; Lanke, K.; Bradley, J.; Sanogo, K.; Kone, D.T.; Diarra, K.; et al. Efficacy and Safety of Primaquine and Methylene Blue for Prevention of Plasmodium Falciparum Transmission in Mali: A Phase 2, Single-Blind, Randomised Controlled Trial. Lancet. Infect. Dis. 2018, 18, 627–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otieno, L.; Oneko, M.; Otieno, W.; Abuodha, J.; Owino, E.; Odero, C.; Mendoza, Y.; Andagalu, B.; Awino, N.; Ivinson, K.; et al. Safety and Immunogenicity of RTS,S/AS01 Malaria Vaccine in Infants and Children with WHO Stage 1 or 2 HIV Disease: A Randomised, Double-Blind, Controlled Trial. Lancet. Infect. Dis. 2016, 16, 1134–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, B.; Dicko, A.; Sagara, I.; Zongo, I.; Tinto, H.; Cairns, M.; Kuepfer, I.; Milligan, P.; Ouedraogo, J.-B.; Doumbo, O.; et al. Seasonal Vaccination against Malaria: A Potential Use for an Imperfect Malaria Vaccine. Malar. J. 2017, 16, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2016, 22, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarkang, P.A.; Appiah-Opong, R.; Ofori, M.F.; Ayong, L.S.; Nyarko, A.K. Application of Multi-Target Phytotherapeutic Concept in Malaria Drug Discovery: A Systems Biology Approach in Biomarker Identification. Biomark. Res. 2016, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Vlietinck, A.J.; Pieters, L.; Apers, S.; Cimanga, K.; Mesia, K.; Tona, L. The Value of Central-African Traditional Medicine for Lead Finding: Some Case Studies. J. Ethnopharmacol. 2015, 174, 607–617. [Google Scholar] [CrossRef]
- Ginsburg, H.; Deharo, E. A Call for Using Natural Compounds in the Development of New Antimalarial Treatments—An Introduction. Malar. J. 2011, 10, S1. [Google Scholar] [CrossRef] [Green Version]
- Ross, R. The Prevention of Malaria; E.P. Dutton & Company: New York, NY, USA, 1910. [Google Scholar]
- Ross, R. Some Quantitative Studies in Epidemiology. Nature 1911, 87, 466–467. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, G. The Analysis of Equilibrium in Malaria. Trop. Dis. Bull. 1952, 49, 813–829. [Google Scholar]
- Nájera, J.A.; González-Silva, M.; Alonso, P.L. Some Lessons for the Future from the Global Malaria Eradication Programme (1955-1969). PLoS Med. 2011, 8, e1000412. [Google Scholar] [CrossRef] [Green Version]
- World Health Assembly. 22 Re-Examination of the Global Strategy of Malaria Eradication: Report of the Director-General; World Health Organization: Geneva, Switzerland, 1969.
- Smith, T.A.; Chitnis, N.; Penny, M.; Tanner, M. Malaria Modeling in the Era of Eradication. Cold. Spring. Harb. Perspect. Med. 2017, 7, a025460. [Google Scholar] [CrossRef]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton Landmarks in Biology; Princeton University Press: Princeton, NJ, USA, 2001; ISBN 978-0-691-08861-7. [Google Scholar]
- Colizza, V.; Barthélemy, M.; Barrat, A.; Vespignani, A. Epidemic Modeling in Complex Realities. CR Biol. 2007, 330, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Kermack, W.O.; McKendrick, A.G. Contributions to the Mathematical Theory of Epidemics—I. Bltn. Mathcal. Biol. 1991, 53, 33–55. [Google Scholar] [CrossRef]
- Aikins, M.K.; Pickering, H.; Greenwood, B.M. Attitudes to Malaria, Traditional Practices and Bednets (Mosquito Nets) as Vector Control Measures: A Comparative Study in Five West African Countries. J. Trop. Med. Hyg. 1994, 97, 81–86. [Google Scholar]
- Smith, D.L.; Hay, S.I.; Noor, A.M.; Snow, R.W. Predicting Changing Malaria Risk after Expanded Insecticide-Treated Net Coverage in Africa. Trends Parasitol. 2009, 25, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L.; Dushoff, J.; Snow, R.W.; Hay, S.I. The Entomological Inoculation Rate and Plasmodium Falciparum Infection in African Children. Nature 2005, 438, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L.; McKenzie, F.E.; Snow, R.W.; Hay, S.I. Revisiting the Basic Reproductive Number for Malaria and Its Implications for Malaria Control. PLoS Biol. 2007, 5, e42. [Google Scholar] [CrossRef]
- Heesterbeek, H.; Anderson, R.M.; Andreasen, V.; Bansal, S.; de Angelis, D.; Dye, C.; Eames, K.T.D.; Edmunds, W.J.; Frost, S.D.W.; Funk, S.; et al. Modeling Infectious Disease Dynamics in the Complex Landscape of Global Health. Science 2015, 347, aaa4339. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.M.; Le Menach, A.; Pothin, E.; Eisele, T.P.; Gething, P.W.; Eckhoff, P.A.; Moonen, B.; Schapira, A.; Smith, D.L. Mapping Multiple Components of Malaria Risk for Improved Targeting of Elimination Interventions. Malar. J. 2017, 16, 459. [Google Scholar] [CrossRef] [Green Version]
- Agusto, F.B.; Teboh-Ewungkem, M.I.; Gumel, A.B. Mathematical Assessment of the Effect of Traditional Beliefs and Customs on the Transmission Dynamics of the 2014 Ebola Outbreaks. BMC Med. 2015, 13, 96. [Google Scholar] [CrossRef] [Green Version]
- Mushayabasa, S.; Bhunu, C.P.; Webb, C.; Dhlamini, M. A Mathematical Model for Assessing the Impact of Poverty on Yaws Eradication. Appl. Math. Model. 2012, 36, 1653–1667. [Google Scholar] [CrossRef] [Green Version]
- Ross, R. Mosquito Brigades and How to Organise Them/by Ronald Ross; Longmans, Green: London, UK, 1902. [Google Scholar]
- Hethcote, H.W. A Thousand and one epidemic models. In Proceedings of the Frontiers in Mathematical Biology; Levin, S.A., Ed.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 504–515. [Google Scholar]
- Xia, S.; Zhou, X.-N.; Liu, J. Systems Thinking in Combating Infectious Diseases. Infect. Dis. Poverty 2017, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Mecoli, M.; de Angelis, V.; Brailsford, S.C. Using System Dynamics to Evaluate Control Strategies for Mosquito-Borne Diseases Spread by Human Travel. Comput. Oper. Res. 2013, 40, 2219–2228. [Google Scholar] [CrossRef]
- Mandal, S.; Sarkar, R.R.; Sinha, S. Mathematical Models of Malaria—A Review. Malar. J. 2011, 10, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agusto, F.B.; del Valle, S.Y.; Blayneh, K.W.; Ngonghala, C.N.; Goncalves, M.J.; Li, N.; Zhao, R.; Gong, H. The Impact of Bed-Net Use on Malaria Prevalence. J. Biol. 2013, 320, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngonghala, C.N.; Del Valle, S.Y.; Zhao, R.; Mohammed-Awel, J. Quantifying the Impact of Decay in Bed-Net Efficacy on Malaria Transmission. J. Biol. 2014, 363, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Le, P.V.V.; Kumar, P.; Ruiz, M.O. Stochastic Lattice-Based Modelling of Malaria Dynamics. Malar. J. 2018, 17, 250. [Google Scholar] [CrossRef] [PubMed]
- Dangerfield, C.E.; Ross, J.V.; Keeling, M.J. Integrating Stochasticity and Network Structure into an Epidemic Model. J. R Soc. Interface 2009, 6, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Parham, P.E.; Michael, E. Outbreak Properties of Epidemic Models: The Roles of Temporal Forcing and Stochasticity on Pathogen Invasion Dynamics. J. Biol. 2011, 271, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.R.; Trauer, J.M.; Gambhir, M.; Richards, J.S.; Maude, R.J.; Keith, J.M.; Flegg, J.A. Agent-Based Models of Malaria Transmission: A Systematic Review. Malar. J. 2018, 17, 299. [Google Scholar] [CrossRef] [Green Version]
- Maude, R.J.; Pontavornpinyo, W.; Saralamba, S.; Aguas, R.; Yeung, S.; Dondorp, A.M.; Day, N.P.; White, N.J.; White, L.J. The Last Man Standing Is the Most Resistant: Eliminating Artemisinin-Resistant Malaria in Cambodia. Malar J. 2009, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, M.A.; Prosper, O.; Lopiano, K.; Ruktanonchai, N.; Caughlin, T.T.; Martcheva, M.; Osenberg, C.W.; Smith, D.L. Spatial Heterogeneity, Host Movement and Mosquito-Borne Disease Transmission. PLoS ONE 2015, 10, e0127552. [Google Scholar] [CrossRef] [PubMed]
- Okell, L.C.; Paintain, L.S.; Webster, J.; Hanson, K.; Lines, J. From Intervention to Impact: Modelling the Potential Mortality Impact Achievable by Different Long-Lasting, Insecticide-Treated Net Delivery Strategies. Malar. J. 2012, 11, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, G.L.; Smith, D.L.; Fidock, D.A. Malaria’s Missing Number: Calculating the Human Component of R0 by a Within-Host Mechanistic Model of Plasmodium Falciparum Infection and Transmission. PLoS Comput. Biol. 2013, 9, e1003025. [Google Scholar] [CrossRef] [PubMed]
- Frost, S.D.W.; Pybus, O.G.; Gog, J.R.; Viboud, C.; Bonhoeffer, S.; Bedford, T. Eight Challenges in Phylodynamic Inference. Epidemics 2015, 10, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Buckee, C.O.; Wesolowski, A.; Eagle, N.; Hansen, E.; Snow, R.W. Mobile Phones and Malaria: Modeling Human and Parasite Travel. Travel. Med. Infect. Dis. 2013, 11, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Ruktanonchai, N.W.; De Leenheer, P.; Tatem, A.J.; Alegana, V.A.; Caughlin, T.T.; Zu Erbach-Schoenberg, E.; Lourenço, C.; Ruktanonchai, C.W.; Smith, D.L. Identifying Malaria Transmission Foci for Elimination Using Human Mobility Data. PLoS Comput. Biol. 2016, 12, e1004846. [Google Scholar] [CrossRef] [Green Version]
- Ruktanonchai, N.W.; Bhavnani, D.; Sorichetta, A.; Bengtsson, L.; Carter, K.H.; Córdoba, R.C.; Le Menach, A.; Lu, X.; Wetter, E.; zu Erbach-Schoenberg, E.; et al. Census-Derived Migration Data as a Tool for Informing Malaria Elimination Policy. Malar. J. 2016, 15, 273. [Google Scholar] [CrossRef] [Green Version]
- Santos-Lozada, A.R.; Howard, J.T.; Verdery, A.M. How Differential Privacy Will Affect Our Understanding of Health Disparities in the United States. Proc. Natl. Acad. Sci. USA 2020, 117, 13405–13412. [Google Scholar] [CrossRef]
- Marino, A.; Bivona, D.A.; Bonacci, P. Updates in Central Nervous System Malaria: Literature Review and Considerations. Curr. Opin. Infect. Dis. 2022, 35, 255–261. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Bhardwaj, J.; Goyal, M.; Prakash, K.; Adnan, M.; Alreshidi, M.M.; Patel, M.; Soni, A.; Redman, W. Immune Responses in Liver and Spleen against Plasmodium Yoelii Pre-Erythrocytic Stages in Swiss Mice Model. J. Adv. Res. 2020, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.K.; Kong, T.F.; Ng, C.S.; Chen, L.; Huang, Y.; Bhagat, A.A.S.; Nguyen, N.-T.; Preiser, P.R.; Han, J. Micromagnetic Resonance Relaxometry for Rapid Label-Free Malaria Diagnosis. Nat. Med. 2014, 20, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.K.; Ng, T.-T.; Loh, T.P. Machine Learning Assistive Rapid, Label-Free Molecular Phenotyping of Blood with Two-Dimensional NMR Correlational Spectroscopy. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, E.; Ferrauto, G.; Schwarzer, E.; Gianolio, E.; Valente, E.; Ulliers, D.; Aime, S.; Skorokhod, O. Relaxometric Studies of Erythrocyte Suspensions Infected by Plasmodium Falciparum: A Tool for Staging Infection and Testing Anti-Malarial Drugs. Magn. Reson. Med. 2020, 84, 3366–3378. [Google Scholar] [CrossRef]
- Arndt, L.; Koleala, T.; Orbán, Á.; Ibam, C.; Lufele, E.; Timinao, L.; Lorry, L.; Butykai, Á.; Kaman, P.; Molnár, A.P.; et al. Magneto-Optical Diagnosis of Symptomatic Malaria in Papua New Guinea. Nat. Commun. 2021, 12, 969. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Hashimoto, M.; Nagatomi, K.; Nogami, T.; Sofue, Y.; Hayashi, T.; Ido, Y.; Yatsushiro, S.; Abe, K.; Kajimoto, K.; et al. Field Evaluation of a Quantitative, and Rapid Malaria Diagnostic System Using a Fluorescent Blue-Ray Optical Device. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Li, A.; Talluri, K. Reviews and Self-Selection Bias with Operational Implications. Manag. Sci. 2021, 67, 7472–7492. [Google Scholar] [CrossRef]
Figure 1.
Synthesis of the method. In gray, the final objective of the review is shown, in green, the investigated databases are indicated, in yellow, the exclusion process is indicated, and in red, the clustering of the selected papers is shown.
Figure 1.
Synthesis of the method. In gray, the final objective of the review is shown, in green, the investigated databases are indicated, in yellow, the exclusion process is indicated, and in red, the clustering of the selected papers is shown.
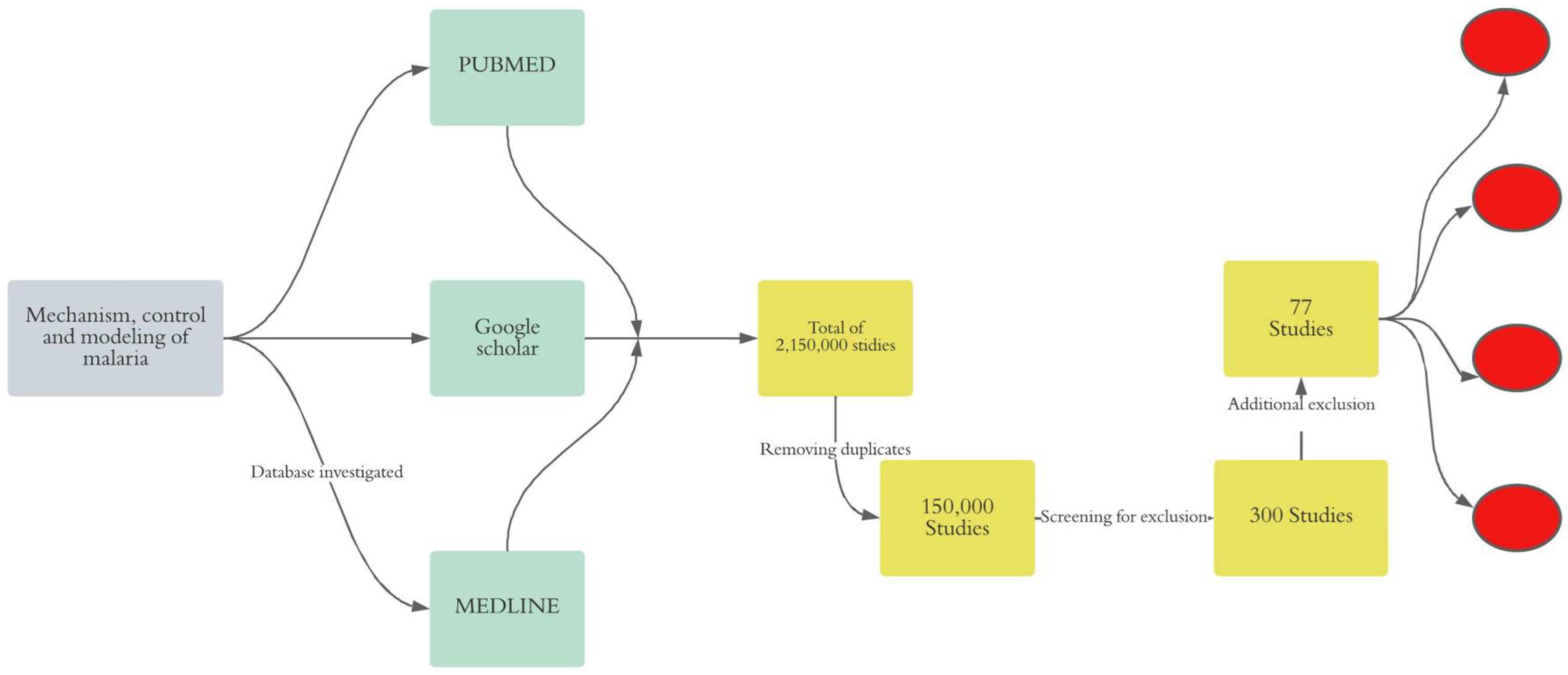
Figure 2.
Overview of the thematic distribution of the selected papers.
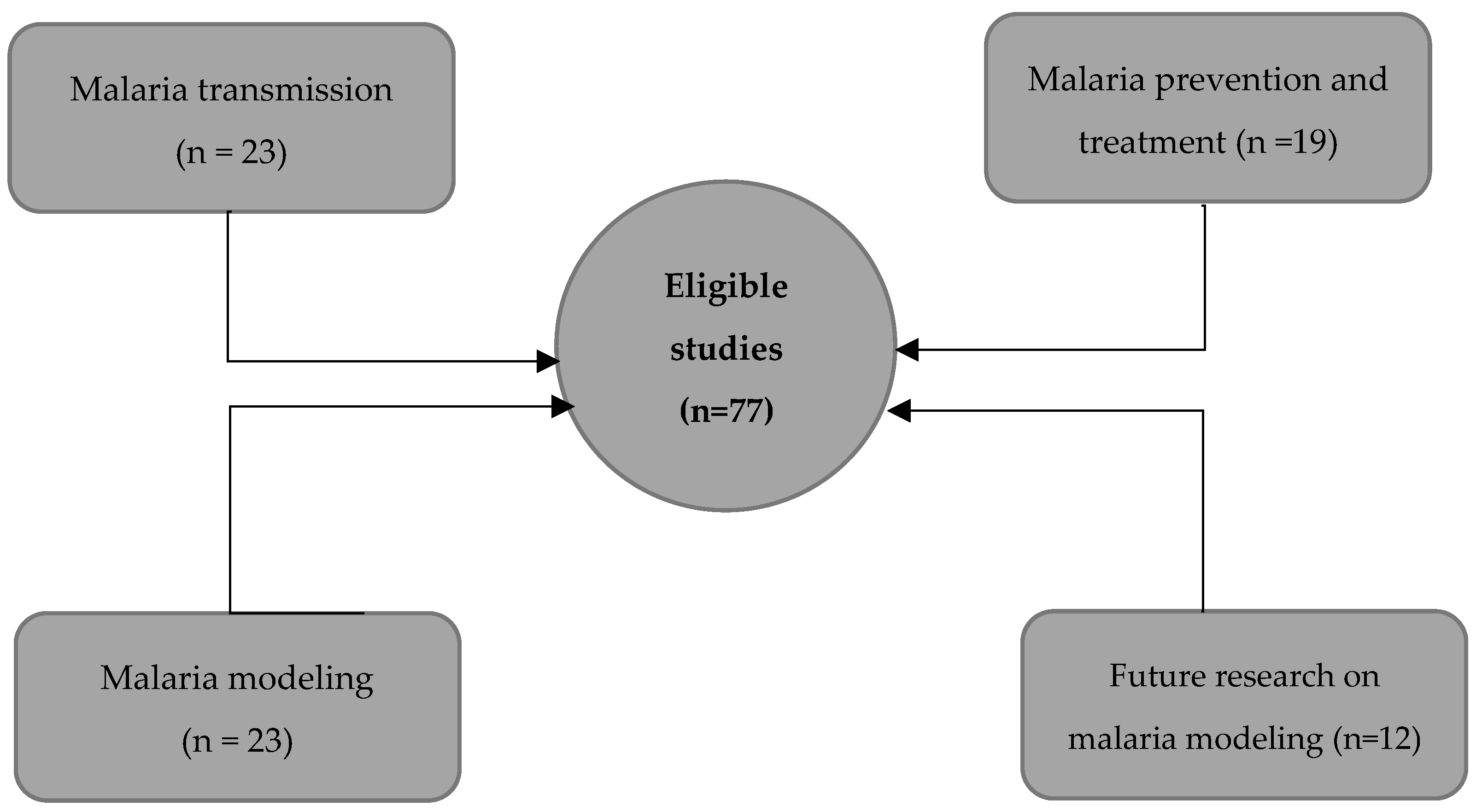
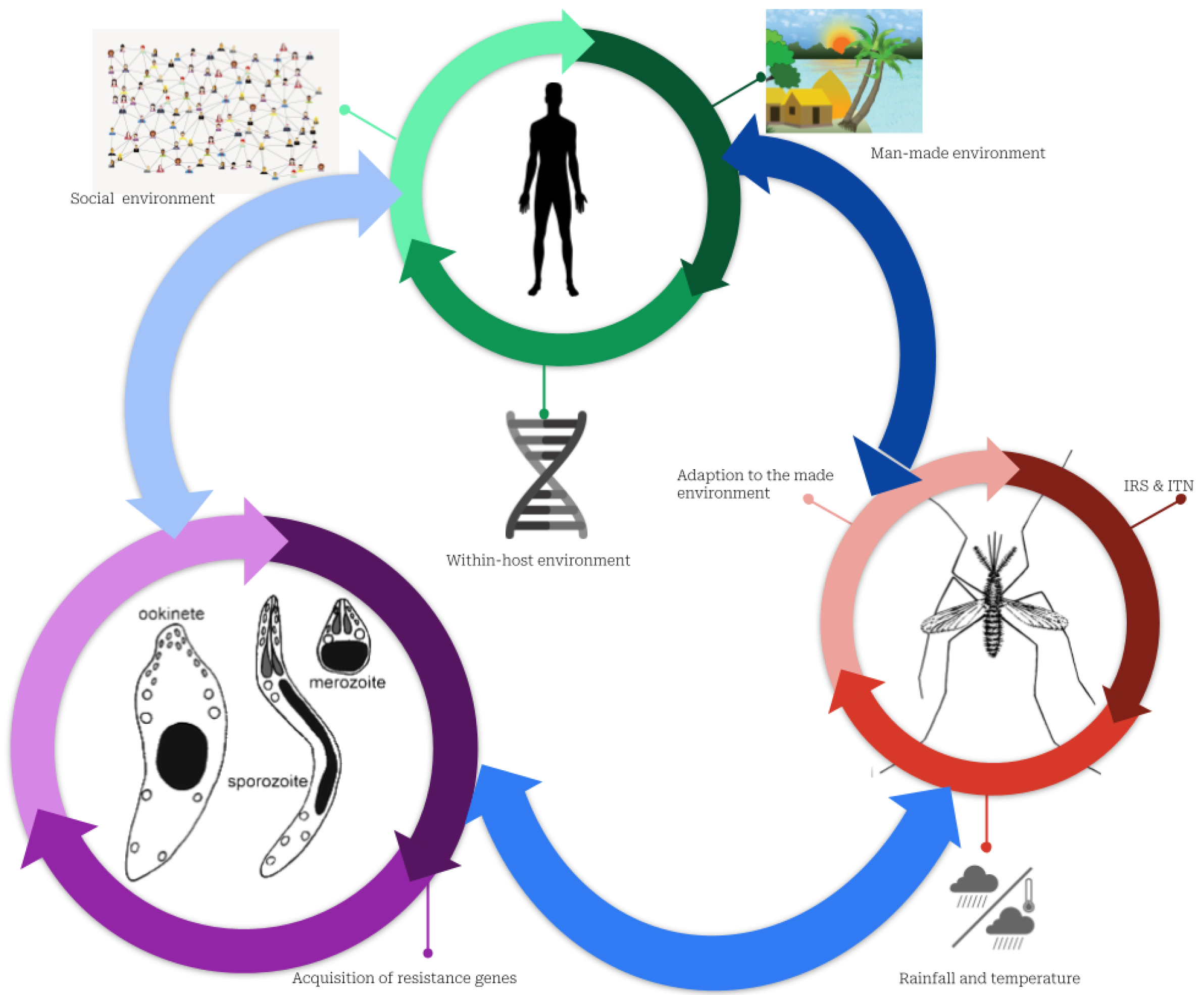
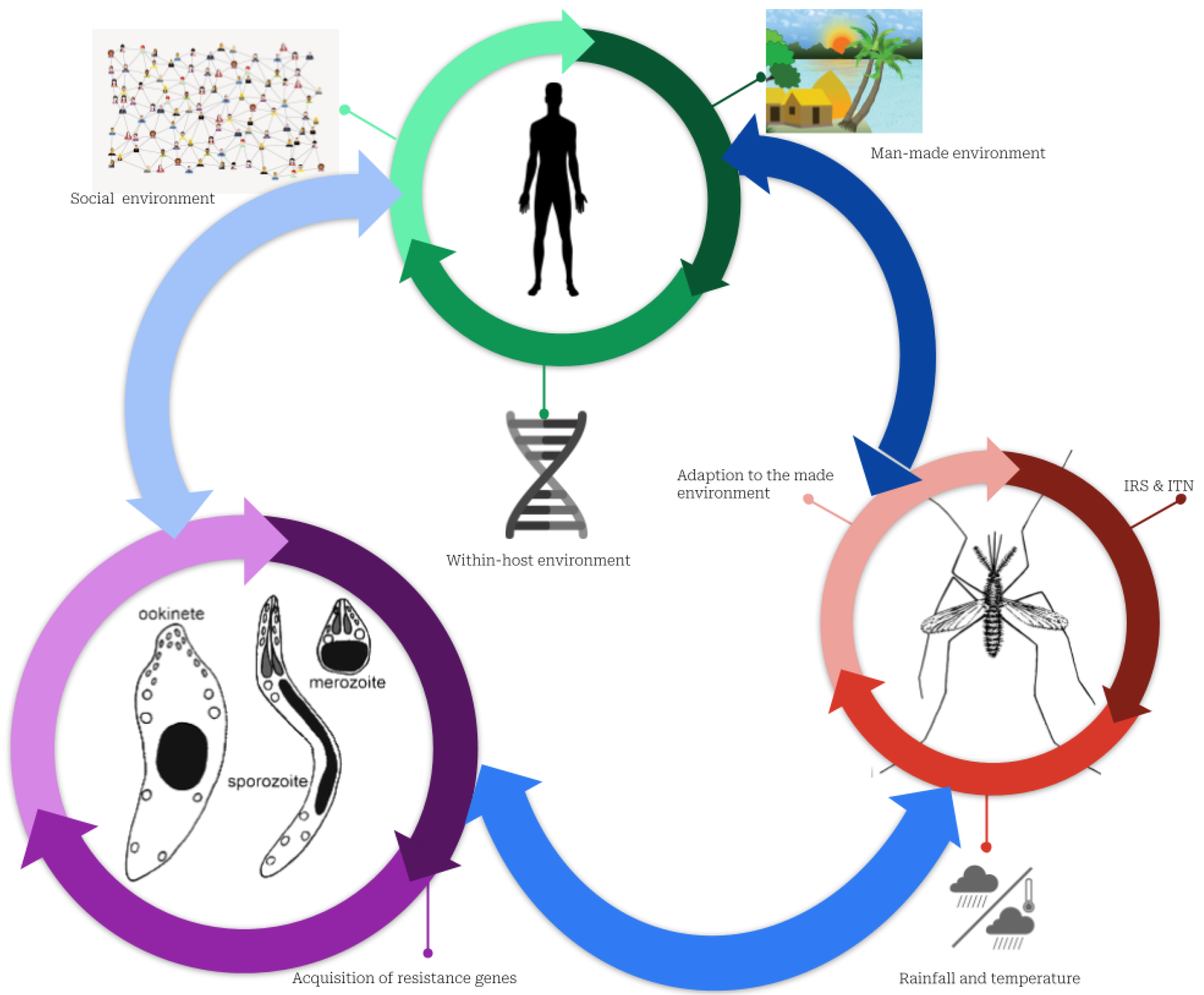
Figure 3.
Complex interactions between humans, mosquitos, and parasites explaining the persistence of malaria. Plasmodium parasites, mosquitoes, and humans constitute three subsystems that interact with one another. The green cycle represents the human sub-system (SS). In addition to the micro-environment, which includes housing conditions, network connectivity, socioeconomic conditions, and the environment within the host (i.e., the individual’s genetic makeup), this SS is characterized by the social environment (social interaction, culture, and behavior regarding malaria risk-taking). The red indicates the SS of mosquitoes defined by the growth conditions favored by humans using or not either ITNs or IRS, and their adaptation to their living environment, including the harsh environmental conditions of cities, such as dirty water and weather conditions. In purple, Plasmodium parasite SS is characterized by interactions with the environment inside the host that may drive resistance to malaria drugs.
Figure 3.
Complex interactions between humans, mosquitos, and parasites explaining the persistence of malaria. Plasmodium parasites, mosquitoes, and humans constitute three subsystems that interact with one another. The green cycle represents the human sub-system (SS). In addition to the micro-environment, which includes housing conditions, network connectivity, socioeconomic conditions, and the environment within the host (i.e., the individual’s genetic makeup), this SS is characterized by the social environment (social interaction, culture, and behavior regarding malaria risk-taking). The red indicates the SS of mosquitoes defined by the growth conditions favored by humans using or not either ITNs or IRS, and their adaptation to their living environment, including the harsh environmental conditions of cities, such as dirty water and weather conditions. In purple, Plasmodium parasite SS is characterized by interactions with the environment inside the host that may drive resistance to malaria drugs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Savi, M.K. An Overview of Malaria Transmission Mechanisms, Control, and Modeling. Med. Sci. 2023, 11, 3. https://doi.org/10.3390/medsci11010003
AMA Style
Savi MK. An Overview of Malaria Transmission Mechanisms, Control, and Modeling. Medical Sciences. 2023; 11(1):3. https://doi.org/10.3390/medsci11010003
Chicago/Turabian StyleSavi, Merveille Koissi. 2023. "An Overview of Malaria Transmission Mechanisms, Control, and Modeling" Medical Sciences 11, no. 1: 3. https://doi.org/10.3390/medsci11010003
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.