
The ESCRT-III Protein Chmp1 Regulates Lipid Storage in the Drosophila Fat Body


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Genetics
2.2. Protein, Triglyceride, and DNA Measurements
2.3. CAFÉ Assay
2.4. Gene Expression Analysis
2.5. Statistics
3. Results
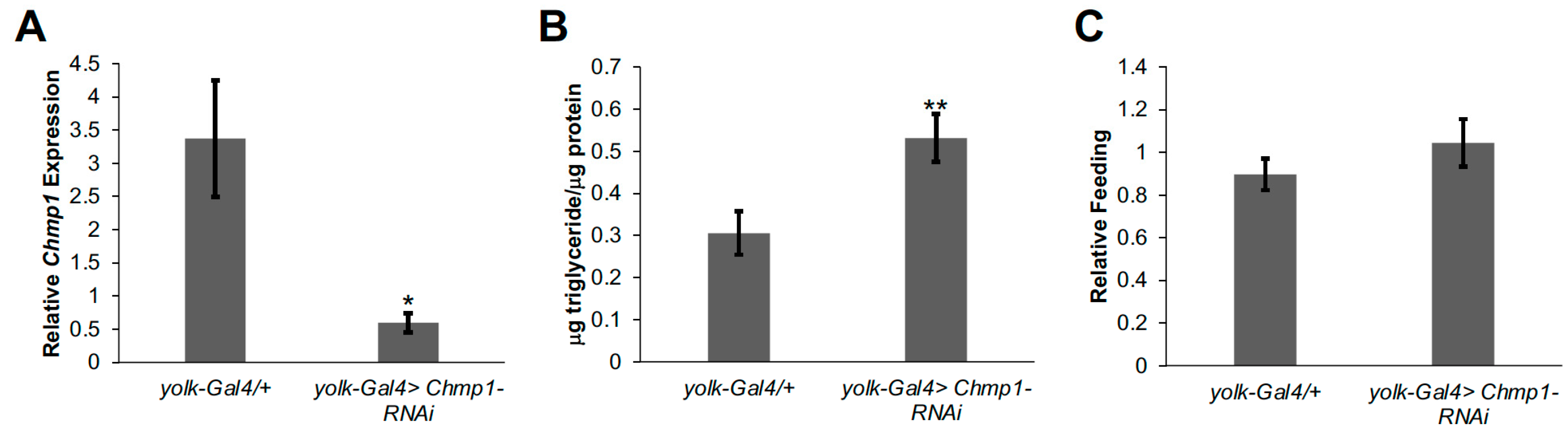
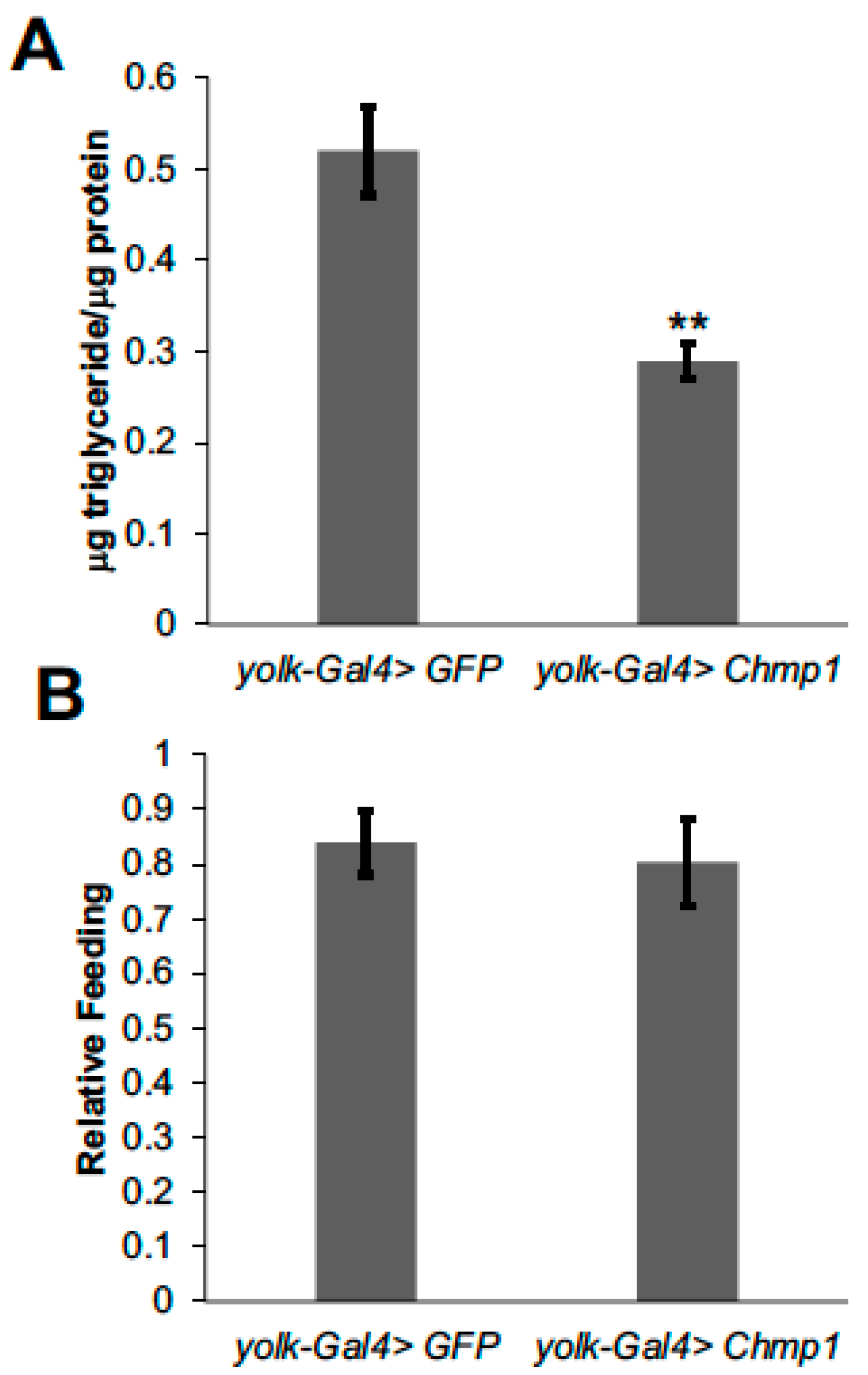
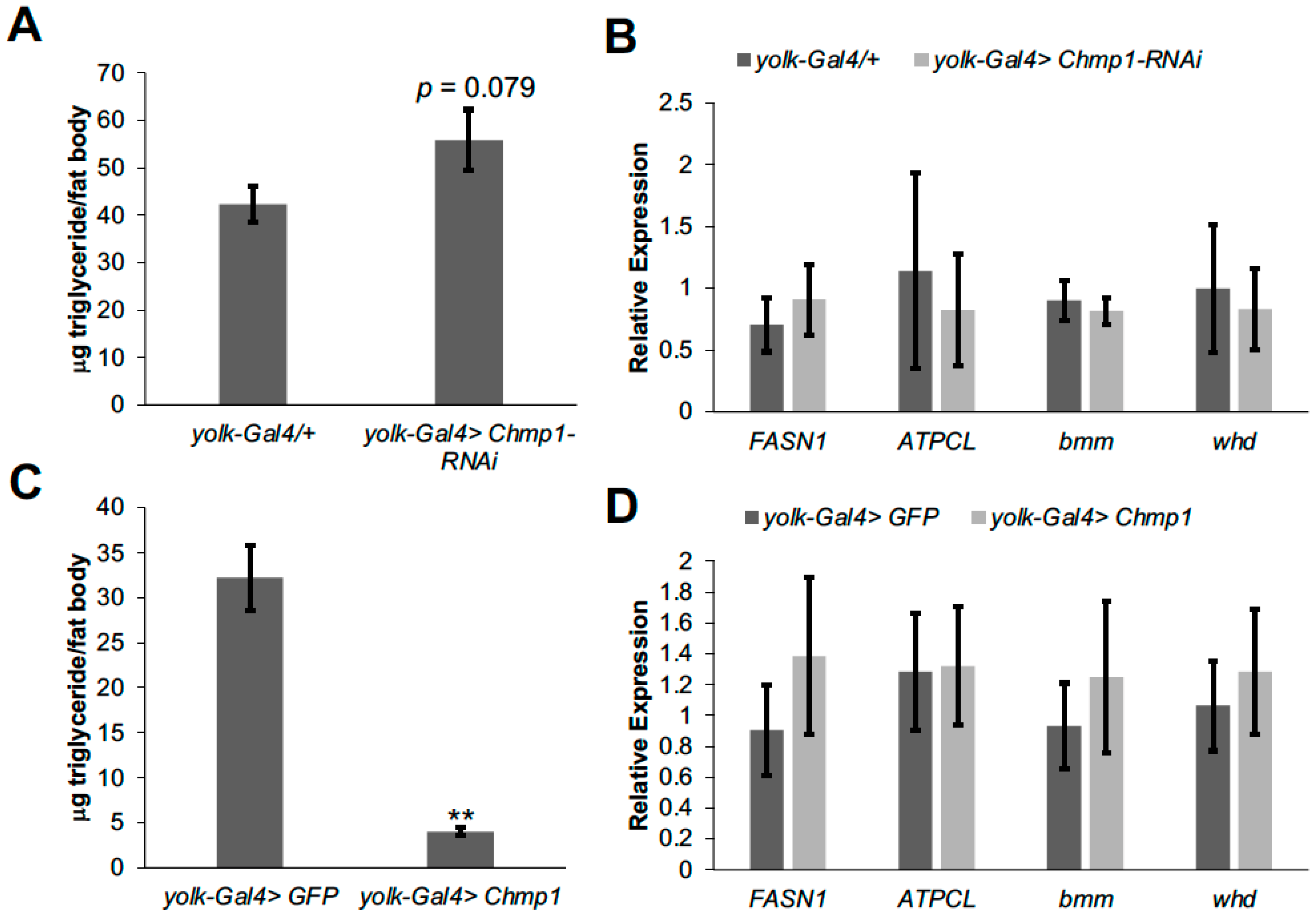
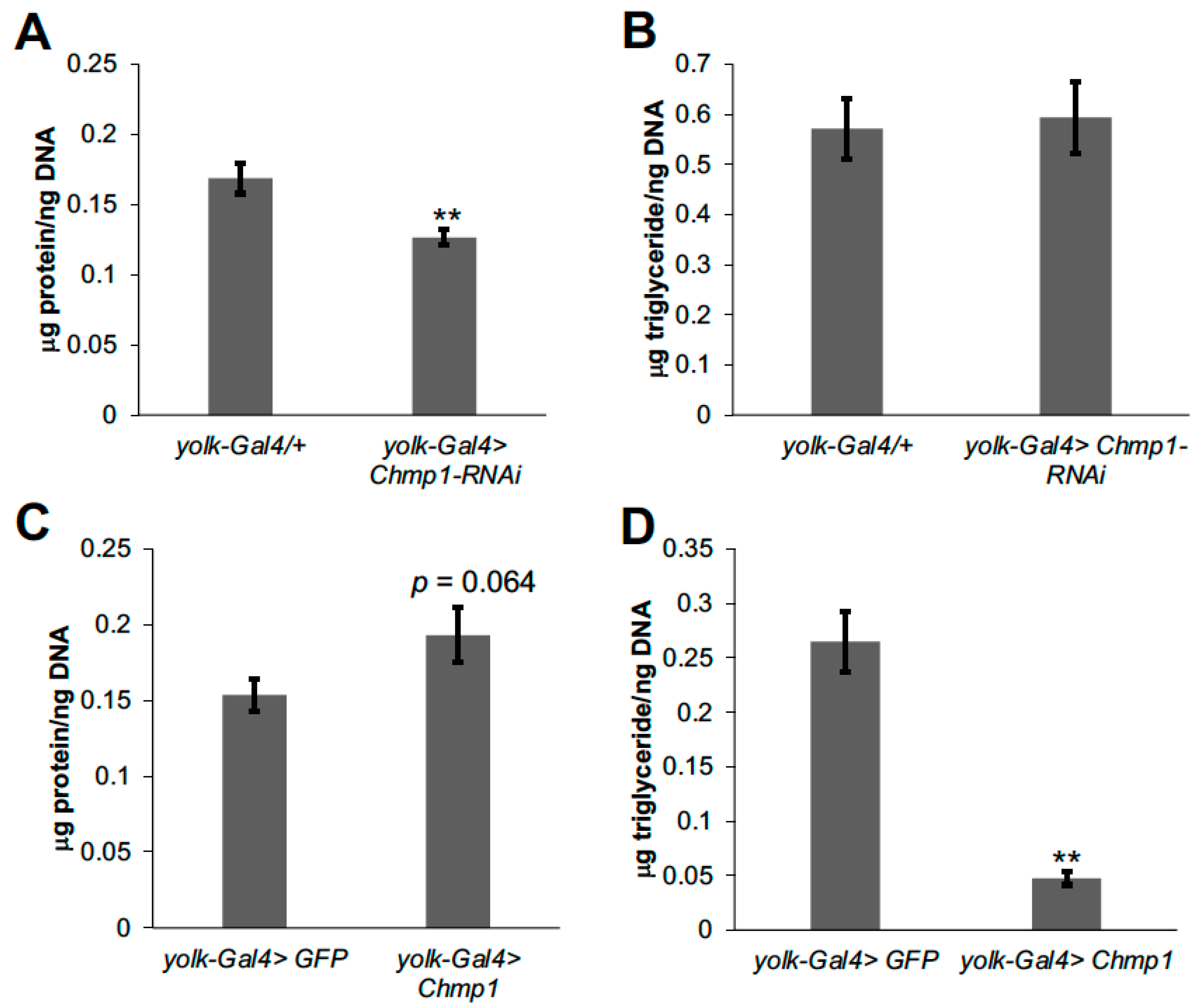
3.1. Chmp1 Acts in the Drosophila Fat Body to Regulate Triglyceride Storage
3.2. Chmp1 Does Not Alter the Expression of Lipid Metabolic Enzyme Genes in the Drosophila Fat Body
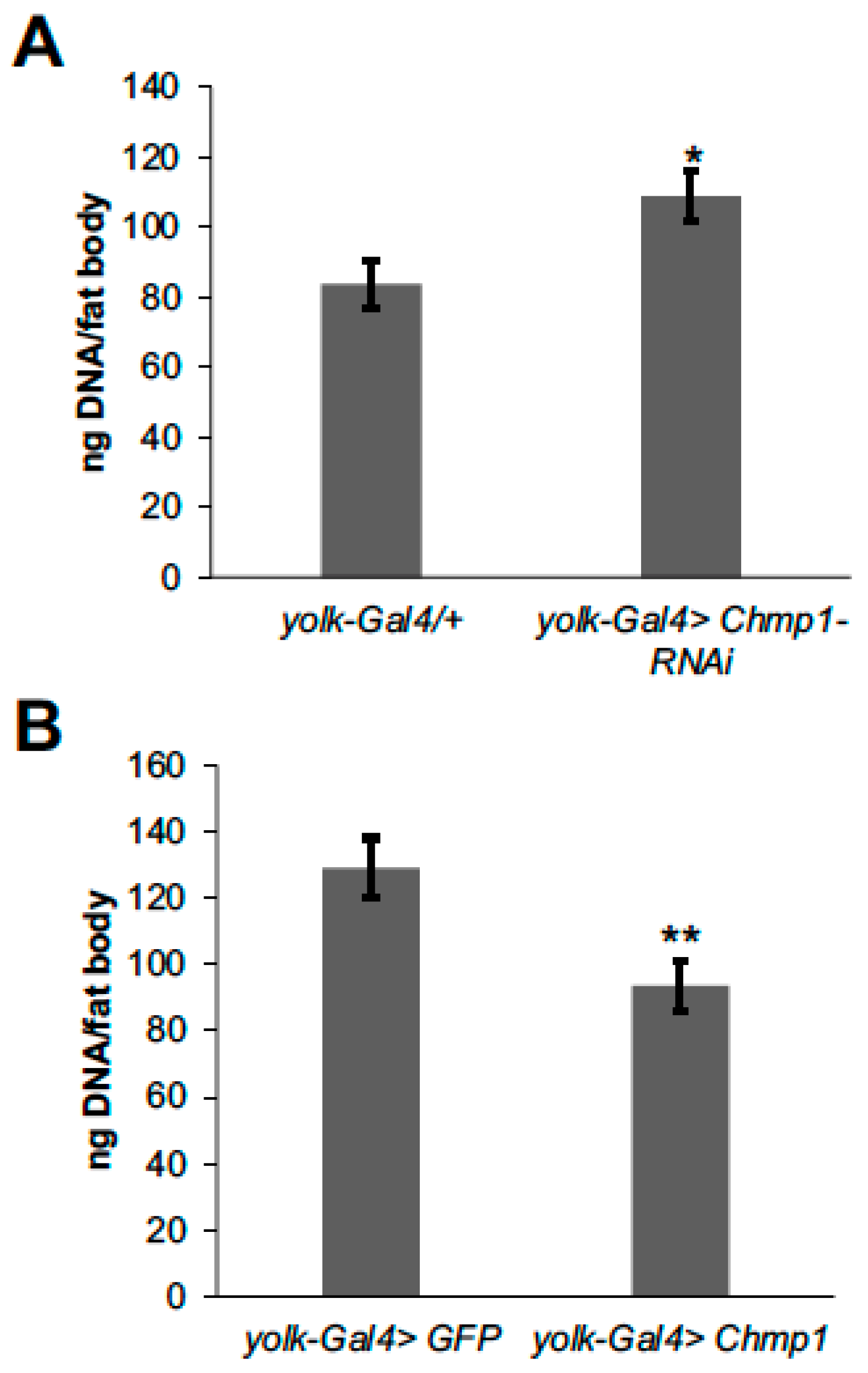
3.3. Chmp1 Regulates Fat Body Cell Number and Size and the Amount of Stored Fat Body Triglyceride
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiu, H.; Schlegel, V. Impact of nutrient overload on metabolic homeostasis. Nutr. Rev. 2018, 76, 693–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghupathi, W.; Raghupathi, V. An Empirical Study of Chronic Diseases in the United States: A Visual Analytics Approach. Int. J. Environ. Res. Public Health 2018, 15, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heier, C.; Klishch, S.; Stilbytska, O.; Semaniuk, U.; Lushchak, O. The Drosophila model to interrogate triacylglycerol biology. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158924. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Kuhnlein, R.P. Drosophila as a model to study obesity and metabolic disease. J. Exp. Biol. 2018, 221, jeb163881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beller, M.; Sztalryd, C.; Southall, N.; Bell, M.; Jackle, H.; Auld, D.S.; Oliver, B. COPI complex is a regulator of lipid homeostasis. PLoS Biol. 2008, 6, e292. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Walther, T.C.; Rao, M.; Stuurman, N.; Goshima, G.; Terayama, K.; Wong, J.S.; Vale, R.D.; Walter, P.; Farese, R.V. Functional genomic screen reveals genes involved in lipid-droplet formation and utilization. Nature 2008, 453, 657–661. [Google Scholar] [CrossRef] [Green Version]
- Pospisilik, J.A.; Schramek, D.; Schnidar, H.; Cronin, S.J.; Nehme, N.T.; Zhang, X.; Knauf, C.; Cani, P.D.; Aumayr, K.; Todoric, J.; et al. Drosophila genome-wide obesity screen reveals hedgehog as a determinant of brown versus white adipose cell fate. Cell 2010, 140, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Beraud-Dufour, S.; Balch, W. A journey through the exocytic pathway. J. Cell Sci. 2002, 115, 1779–1780. [Google Scholar] [CrossRef]
- Hurley, J.H.; Hanson, P.I. Membrane budding and scission by the ESCRT machinery: It’s all in the neck. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [Google Scholar] [CrossRef]
- Michelet, X.; Djeddi, A.; Legouis, R. Developmental and cellular functions of the ESCRT machinery in pluricellular organisms. Biol. Cell 2010, 102, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Ouahoud, S.; Fiet, M.D.; Martinez-Montanes, F.; Ejsing, C.S.; Kuss, O.; Roden, M.; Markgraf, D.F. Lipid droplet consumption is functionally coupled to vacuole homeostasis independent of lipophagy. J. Cell Sci. 2018, 131, jcs213876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickerson, D.P.; West, M.; Odorizzi, G. Did2 coordinates Vps4-mediated dissociation of ESCRT-III from endosomes. J. Cell Biol. 2006, 175, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Obita, T.; Saksena, S.; Ghazi-Tabatabai, S.; Gill, D.J.; Perisic, O.; Emr, S.D.; Williams, R.L. Structural basis for selective recognition of ESCRT-III by the AAA ATPase Vps4. Nature 2007, 449, 735–739. [Google Scholar] [CrossRef]
- Georgel, P.; Naitza, S.; Kappler, C.; Ferrandon, D.; Zachary, D.; Swimmer, C.; Kopczynski, C.; Duyk, G.; Reichhart, J.M.; Hoffmann, J.A. Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev. Cell 2001, 1, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Valentine, M.; Hogan, J.; Collier, S. The Drosophila Chmp1 protein determines wing cell fate through regulation of epidermal growth factor receptor signaling. Dev. Dyn. 2014, 243, 977–987. [Google Scholar] [CrossRef]
- Bennick, R.A.; Nagengast, A.A.; DiAngelo, J.R. The SR proteins SF2 and RBP1 regulate triglyceride storage in the fat body of Drosophila. Biochem. Biophys. Res. Commun. 2019, 516, 928–933. [Google Scholar] [CrossRef]
- DiAngelo, J.R.; Birnbaum, M.J. Regulation of fat cell mass by insulin in Drosophila melanogaster. Mol. Cell. Biol. 2009, 29, 6341–6352. [Google Scholar] [CrossRef] [Green Version]
- Ja, W.W.; Carvalho, G.B.; Mak, E.M.; de la Rosa, N.N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of Drosophila and the CAFE assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8253–8256. [Google Scholar] [CrossRef] [Green Version]
- Marguerat, S.; Bahler, J. Coordinating genome expression with cell size. Trends Genet. 2012, 28, 560–565. [Google Scholar] [CrossRef]
- Rodahl, L.M.; Haglund, K.; Sem-Jacobsen, C.; Wendler, F.; Vincent, J.P.; Lindmo, K.; Rusten, T.E.; Stenmark, H. Disruption of Vps4 and JNK function in Drosophila causes tumour growth. PLoS ONE 2009, 4, e4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaccari, T.; Rusten, T.E.; Menut, L.; Nezis, I.P.; Brech, A.; Stenmark, H.; Bilder, D. Comparative analysis of ESCRT-I, ESCRT-II and ESCRT-III function in Drosophila by efficient isolation of ESCRT mutants. J. Cell Sci. 2009, 122, 2413–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacomin, A.C.; Fauvarque, M.O.; Taillebourg, E. A functional endosomal pathway is necessary for lysosome biogenesis in Drosophila. BMC Cell Biol. 2016, 17, 36. [Google Scholar] [CrossRef] [Green Version]
- Stump, C.L.; Casero, R.A.; Phanstiel, O.; DiAngelo, J.R.; Nowotarski, S.L. Elucidating the Role of Chmp1 Overexpression in the Transport of Polyamines in Drosophila melanogaster. Med Sci. 2022, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [Green Version]
- Morales, T.S.; Avis, E.C.; Paskowski, E.K.; Shabar, H.; Nowotarski, S.L.; DiAngelo, J.R. The Role of Spermidine Synthase (SpdS) and Spermine Synthase (Sms) in Regulating Triglyceride Storage in Drosophila. Med. Sci. 2021, 9, 27. [Google Scholar] [CrossRef]
- Yuan, F.; Zhang, L.; Cao, Y.; Gao, W.; Zhao, C.; Fang, Y.; Zahedi, K.; Soleimani, M.; Lu, X.; Fang, Z.; et al. Spermidine/spermine N1-acetyltransferase-mediated polyamine catabolism regulates beige adipocyte biogenesis. Metabolism 2018, 85, 298–304. [Google Scholar] [CrossRef]
- Li, J.; Belogortseva, N.; Porter, D.; Park, M. Chmp1A functions as a novel tumor suppressor gene in human embryonic kidney and ductal pancreatic tumor cells. Cell Cycle 2008, 7, 2886–2893. [Google Scholar] [CrossRef]
- You, Z.; Xin, Y.; Liu, Y.; Sun, J.; Zhou, G.; Gao, H.; Xu, P.; Chen, Y.; Chen, G.; Zhang, L.; et al. Chmp1A acts as a tumor suppressor gene that inhibits proliferation of renal cell carcinoma. Cancer Lett. 2012, 319, 190–196. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fruin, A.M.; Leon, K.E.; DiAngelo, J.R. The ESCRT-III Protein Chmp1 Regulates Lipid Storage in the Drosophila Fat Body. Med. Sci. 2023, 11, 5. https://doi.org/10.3390/medsci11010005
Fruin AM, Leon KE, DiAngelo JR. The ESCRT-III Protein Chmp1 Regulates Lipid Storage in the Drosophila Fat Body. Medical Sciences. 2023; 11(1):5. https://doi.org/10.3390/medsci11010005
Chicago/Turabian StyleFruin, Austin M., Kelly E. Leon, and Justin R. DiAngelo. 2023. "The ESCRT-III Protein Chmp1 Regulates Lipid Storage in the Drosophila Fat Body" Medical Sciences 11, no. 1: 5. https://doi.org/10.3390/medsci11010005
APA StyleFruin, A. M., Leon, K. E., & DiAngelo, J. R. (2023). The ESCRT-III Protein Chmp1 Regulates Lipid Storage in the Drosophila Fat Body. Medical Sciences, 11(1), 5. https://doi.org/10.3390/medsci11010005