Cellular and Animal Model Studies on the Growth Inhibitory Effects of Polyamine Analogues on Breast Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
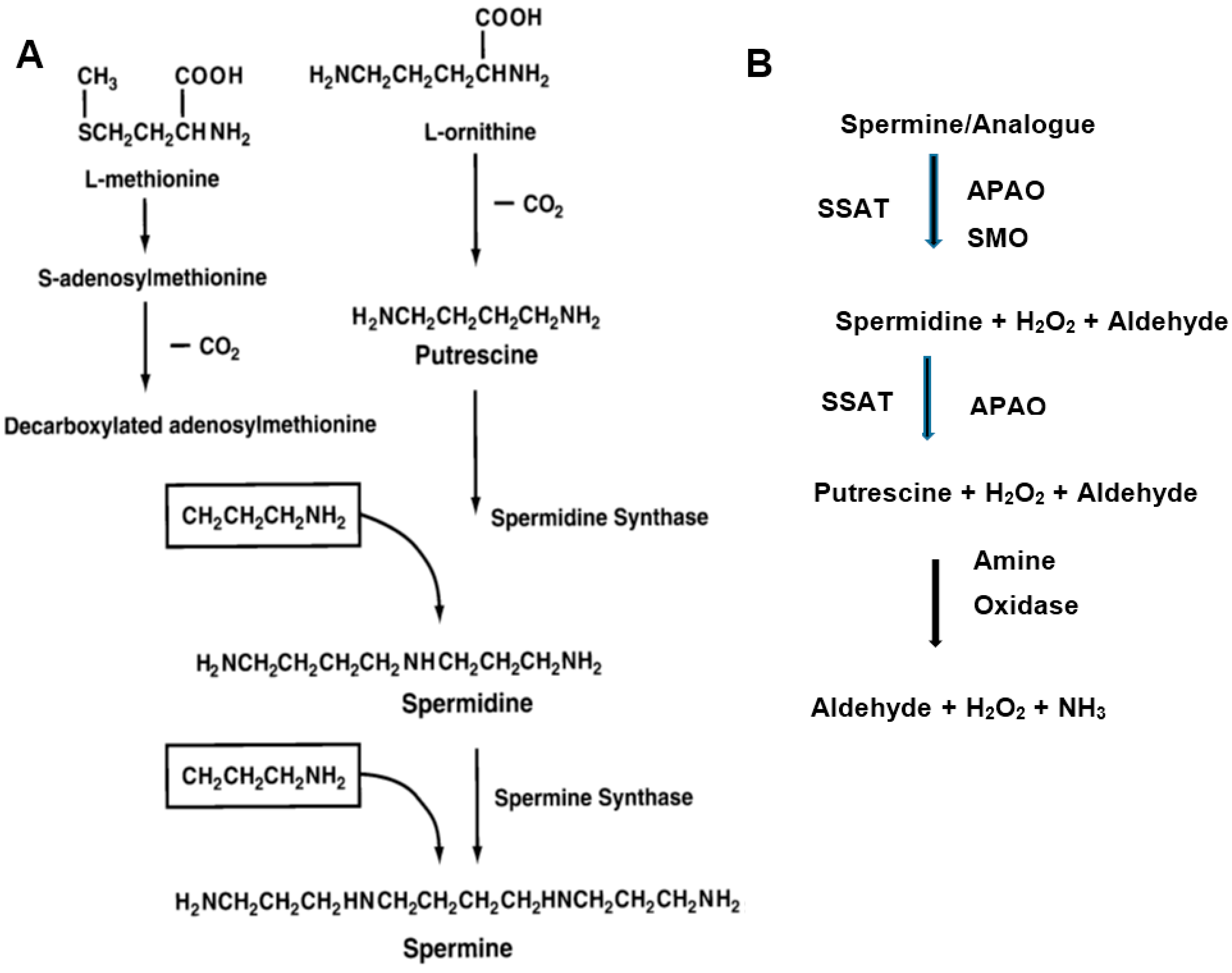
2. Polyamine Metabolism and Breast Cancer
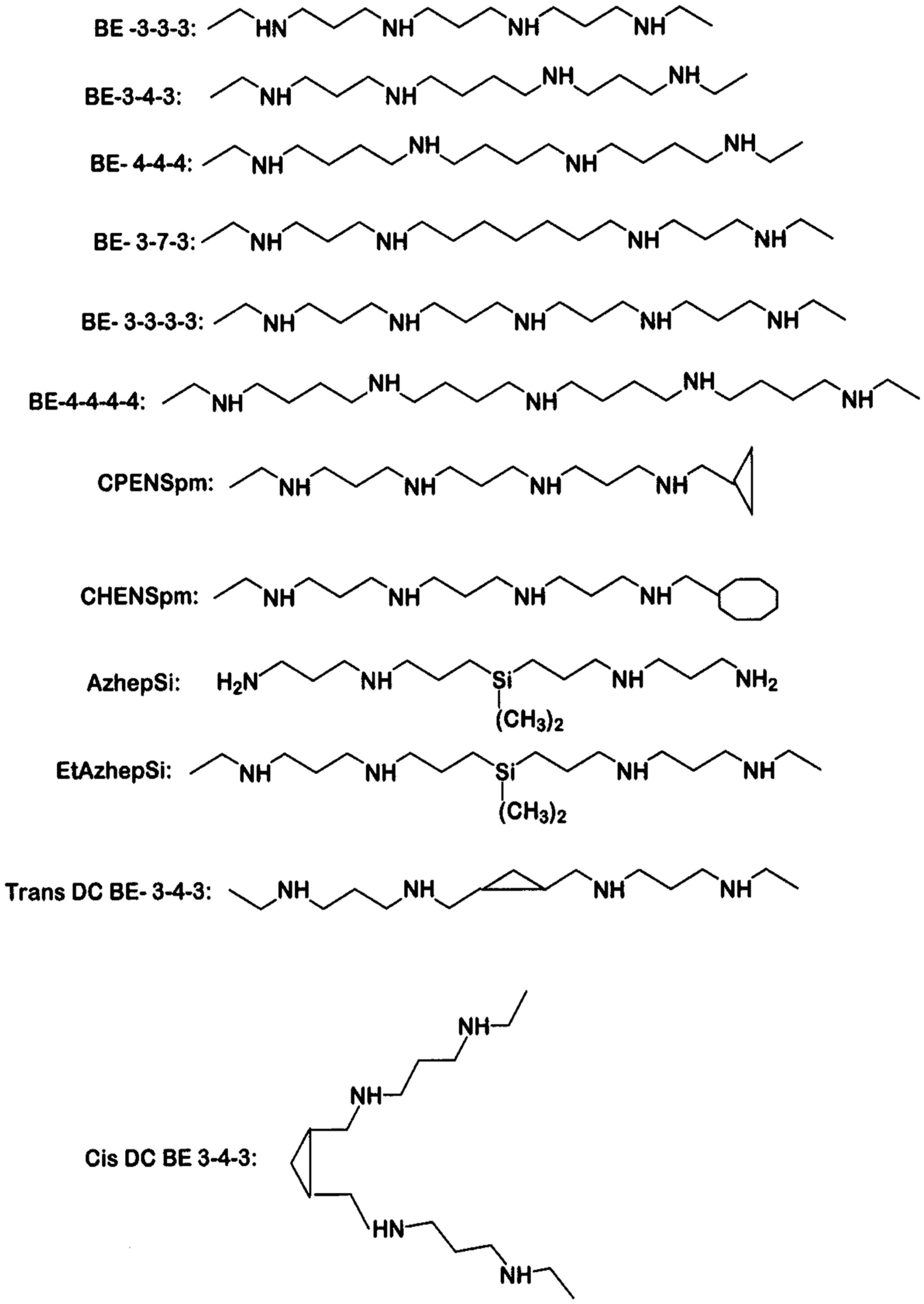
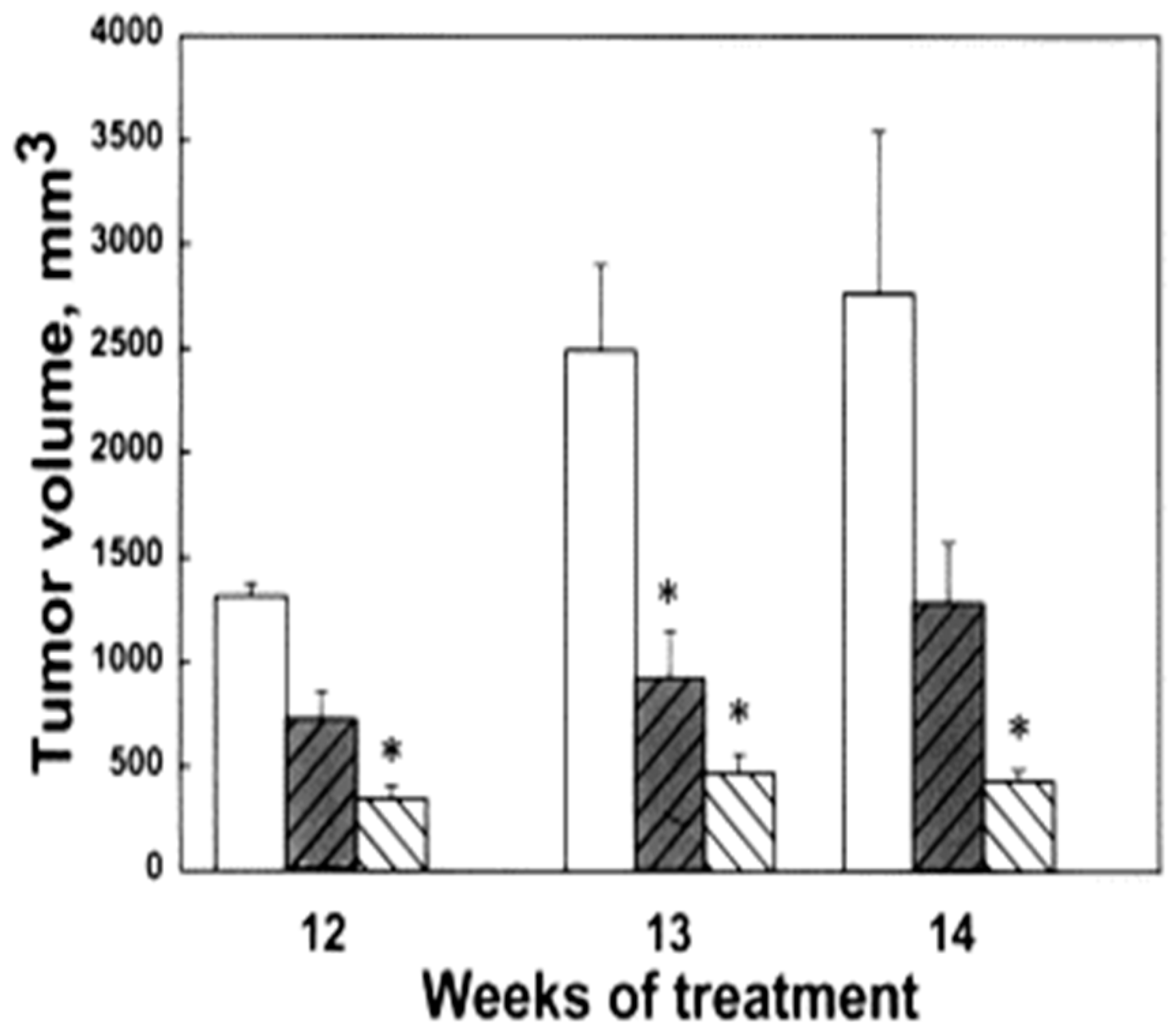
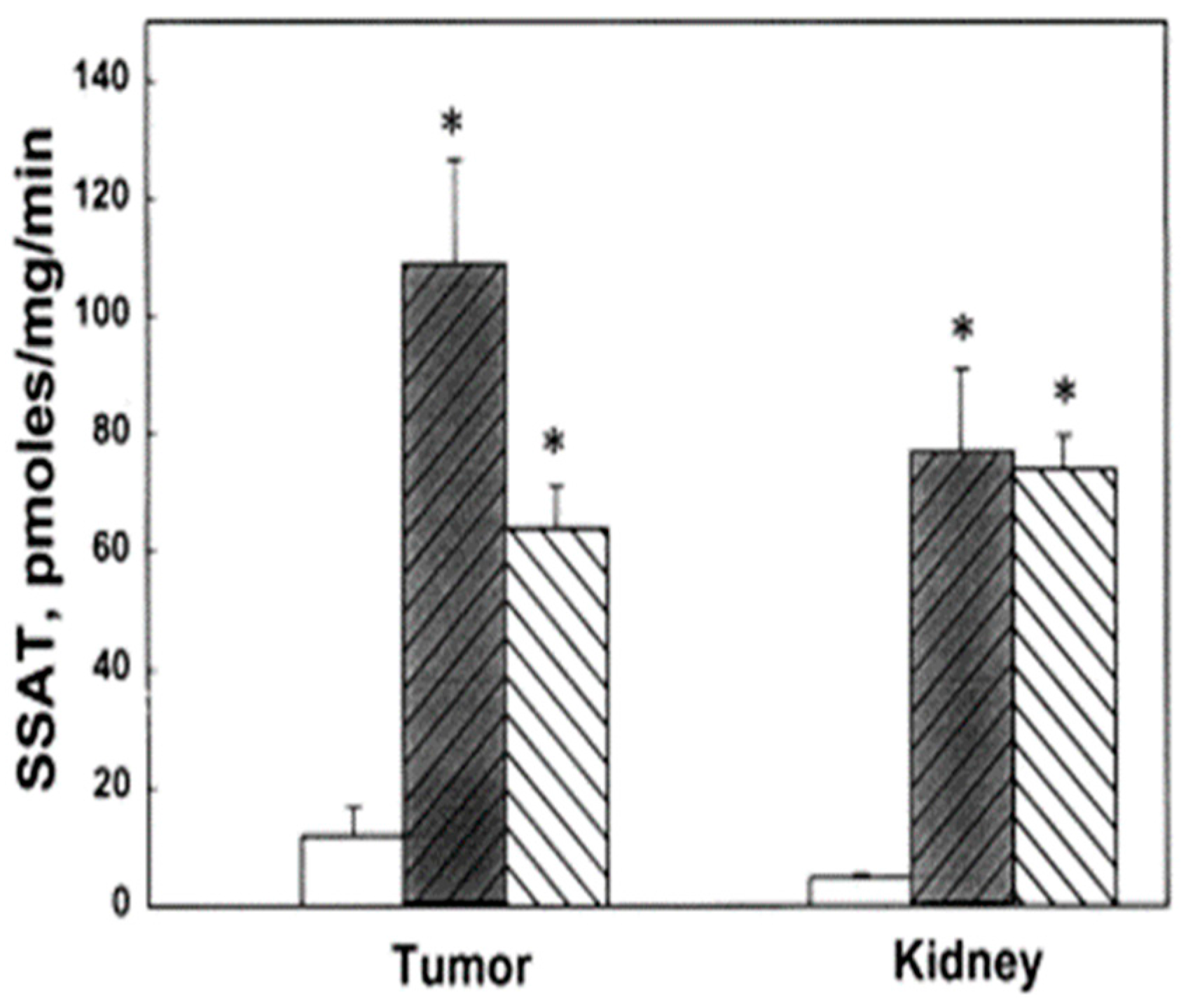
3. Polyamine Analogues and Breast Cancer Therapeutics
4. Conclusions
Author Contributions
Conflicts of Interest
References
- World Cancer Research Fund International, Breast Cancer Statistics. 2017. Available online: http://www.wcrf.org/int/cancer-facts-figures/data-specific-cancers/breast-cancer-statistics (accessed on 15 January 2018).
- American Cancer Society. Cancer Facts and Figures 2017; American Cancer Society: Atlanta, GA, USA, 2017. [Google Scholar]
- Polyak, K. Heterogeneity in breast cancer. J. Clin. Investig. 2011, 121, 3786–3788. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.; Tarulli, G.; Portman, N.; Hickey, T.E.; Tilley, W.D.; Palmieri, C. Pushing estrogen receptor around in breast cancer. Endocr. Relat. Cancer 2016, 23, T227–T241. [Google Scholar] [CrossRef] [PubMed]
- Nagini, S. Breast cancer: Current molecular therapeutic targets and new players. Anticancer Agents Med. Chem. 2017, 17, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Gallo, M.A.; Thomas, T.J. Estrogen receptors as targets for drug development for breast cancer, osteoporosis and cardiovascular diseases. Curr. Cancer Drug Targets 2004, 4, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.T.; Jordan, V.C. The biological role of estrogen receptors alpha and beta in cancer. Crit. Rev. Oncol. Hematol. 2004, 50, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Maximov, P.Y.; Lee, T.M.; Jordan, V.C. The discovery and development of selective estrogen receptor modulators (SERMs) for clinical practice. Curr. Clin. Pharmacol. 2013, 8, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, D.; Kumari, K.M.; Avin, S.; Babu, V.A.M. The evolutionary tale and future directions of aromatase inhibitors in breast carcinoma. Anticancer Agents Med. Chem. 2017, 17, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Baselga, J.; Coleman, R.E.; Cortés, J.; Janni, W. Advances in the management of HER2-positive early breast cancer. Crit. Rev. Oncol. Hematol. 2017, 119, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; He, G.; Yan, S.; Chen, C.; Song, L.; Rosol, T.J.; Deng, X. Triple-negative breast cancer: Is there a treatment on the horizon? Oncotarget 2017, 8, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.R.; Massarweh, S.A. Endocrine therapy and strategies to overcome therapeutic resistance in breast cancer. Curr. Probl. Cancer 2016, 40, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Balabhadrapathruni, S.; Gallo, M.A.; Thomas, T.J. Development of polyamine analogs as cancer therapeutic agents. Oncol. Res. 2002, 13, 123–135. [Google Scholar] [PubMed]
- Cervelli, M.; Pietropaoli, S.; Signore, F.; Amendola, R.; Mariottini, P. Polyamines metabolism and breast cancer: State of the art and perspectives. Breast Cancer Res. Treat. 2014, 148, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Vijayanathan, V.; Agostinelli, E.; Thomas, T.; Thomas, T.J. Innovative approaches to the use of polyamines for DNA nanoparticle preparation for gene therapy. Amino Acids 2014, 46, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.E.; Hahm, H.A.; McCloskey, D.E.; Woster, P.M.; Casero, R.A., Jr. Clinical aspects of cell death in breast cancer: The polyamine pathway as a new target for treatment. Endocr. Relat. Cancer 1999, 6, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Murray-Stewart, T.R.; Woster, P.M.; Casero, R.A., Jr. Targeting polyamine metabolism for cancer therapy and prevention. Biochem. J. 2016, 473, 2937–2953. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Polyamines in cell growth and cell death: Molecular mechanisms and therapeutic applications. Cell. Mol. Life Sci. 2001, 58, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining mysteries of molecular biology: The role of polyamines in the cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Polyamine metabolism and cancer. J. Cell. Mol. Med. 2003, 7, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Nowotarski, S.L.; Woster, P.M.; Casero, R.A., Jr. Polyamines and cancer: Implications for chemotherapy and chemoprevention. Expert Rev. Mol. Med. 2013, 15, e3. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.J.; Tajmir-Riahi, H.A.; Thomas, T. Polyamine-DNA interactions and development of gene delivery vehicles. Amino Acids 2016, 48, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.J.; Thomas, T. Collapse of DNA in packaging and cellular transport. Int. J. Biol. Macromol. 2018, 109, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Chanphai, P.; Thomas, T.J.; Tajmir-Riahi, H.A. Conjugation of biogenic and synthetic polyamines with serum proteins: A comprehensive review. Int. J. Biol. Macromol. 2016, 92, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Bignon, E.; Chan, C.H.; Morell, C.; Monari, A.; Ravanat, J.L.; Dumont, E. Molecular dynamics insights into polyamine-DNA binding modes: Implications for cross-link selectivity. Chemistry 2017, 23, 12845–12852. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Thomas, T.J.; Lewis, J.S.; Klinge, C.M.; Shirahata, A.; Gelinas, C.; Thomas, T. Regulation of estrogenic and nuclear factor κB functions by polyamines and their role in polyamine analog-induced apoptosis of breast cancer cells. Oncogene 2001, 20, 1715–1729. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Thomas, T.; Shirahata, A.; Sigal, L.H.; Thomas, T.J. Activation of nuclear factor κB by polyamines in breast cancer cells. Biochemistry 1999, 38, 14763–14774. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jin, L.; Casero, R.A.; Davidson, N.E.; Huang, Y. Role of ornithine decarboxylase in regulation of estrogen receptor alpha expression and growth in human breast cancer cells. Breast Cancer Res. Treat. 2012, 136, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Vijayanathan, V.; Thomas, T.J.; Nair, S.K.; Shirahata, A.; Gallo, M.A.; Thomas, T. Bending of the estrogen response element by polyamines and estrogen receptors α and β: A fluorescence resonance energy transfer study. Int. J. Biochem. Cell Biol. 2006, 38, 1191–1206. [Google Scholar] [CrossRef] [PubMed]
- Casero, R.A.; Pegg, A.E. Polyamine catabolism and disease. Biochem. J. 2009, 421, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Agostinelli, E.; Vianello, F.; Magliulo, G.; Thomas, T.; Thomas, T.J. Nanoparticle strategies for cancer therapeutics: Nucleic acids, polyamines, bovine serum amine oxidase and iron oxide nanoparticles. Int. J. Oncol. 2015, 46, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.K.; Chaturvedi, R.; Piazuelo, M.J.; Coburn, L.A.; Williams, C.S.; Delgado, A.G.; Casero, R.A.; Pegg, A.G.; Schwartz, D.A.; Wilson, K.T. Increased expression and cellular localization of spermine oxidase in ulcerative colitis and relationship to disease activity. Inflamm. Bowel Dis. 2010, 16, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Agostinelli, E.; Arancia, G.; Vedova, L.D.; Belli, F.; Marra, M.; Salvi, M.; Toninello, A. The biological functions of polyamine oxidation products by amine oxidases: Perspectives of clinical applications. Amino Acids 2004, 27, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.J.; Wallace, H.M. The polyamine transport system as a target for anticancer drug development. Amino Acids 2010, 38, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Kahana, C. Antizyme and antizyme inhibitor, a regulatory tango. Cell. Mol. Life Sci. 2009, 66, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Murray-Stewart, T.; Casero, R.A. Regulation of polyamine metabolism by curcumin for cancer prevention and therapy. Med. Sci. (Basel) 2017, 5, 38. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.J.; Thomas, T.; John, S.; Hsu, H.C.; Yang, P.; Keinänen, T.A.; Hyvönen, M.T. Tamoxifen metabolite endoxifen interferes with the polyamine pathway in breast cancer. Amino Acids 2016, 48, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Estradiol control of ornithine decarboxylase mRNA, enzyme activity, and polyamine levels in MCF-7 breast cancer cells: Therapeutic implications. Breast Cancer Res. Treat. 1994, 29, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Regulation of cyclin B1 by estradiol and polyamines in MCF-7 breast cancer cells. Cancer Res. 1994, 54, 1077–1084. [Google Scholar] [PubMed]
- Manni, A.; Badger, B.; Glikman, P.; Bartholomew, M.; Santner, S.; Demers, L. Individual and combined effects of α-difluoromethylornithine and ovariectomy on the growth and polyamine milieu of experimental breast cancer in rats. Cancer Res. 1989, 49, 3529–3534. [Google Scholar] [PubMed]
- Alexiou, G.A.; Lianos, G.D.; Ragos, V.; Galani, V.; Kyritsis, A.P. Difluoromethylornithine in cancer: New advances. Future Oncol. 2017, 13, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Faaland, C.A.; Adhikarakunnathu, S.; Thomas, T.J. Structure-activity relations of S-adenosylmethionine decarboxylase inhibitors on the growth of MCF-7 breast cancer cells. Breast Cancer Res. Treat. 1996, 39, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, M.; Bellavia, G.; Fratini, E.; Amendola, R.; Polticelli, F.; Barba, M.; Federico, R.; Signore, F.; Gucciardo, G.; Grillo, R.; et al. Spermine oxidase (SMO) activity in breast tumor tissues and biochemical analysis of the anticancer spermine analogues BENSpm and CPENSpm. BMC Cancer 2010, 10, 555. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M. The physiological role of the polyamines. Eur. J. Clin. Investig. 2000, 30, 1–3. [Google Scholar] [CrossRef]
- Hu, X.; Washington, S.; Verderame, M.F.; Demers, L.M.; Mauger, D.; Manni, A. Biological activity of the S-adenosylmethionine decarboxylase inhibitor SAM486A in human breast cancer cells in vitro and in vivo. Int. J. Oncol. 2004, 25, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Richert, M.M.; Phadke, P.A.; Matters, G.; DiGirolamo, D.J.; Washington, S.; Demers, L.M.; Bond, J.S.; Manni, A.; Welch, D.R. Metastasis of hormone-independent breast cancer to lung and bone is decreased by α-difluoromethylornithine treatment. Breast Cancer Res. 2005, 7, R819. [Google Scholar] [CrossRef] [PubMed]
- Fabian, C.J.; Kimler, B.F.; Brady, D.A.; Mayo, M.S.; Chang, C.H.; Ferraro, J.A.; Zalles, C.M.; Stanton, A.L.; Masood, S.; Grizzle, W.E.; et al. A phase II breast cancer chemoprevention trial of oral α-difluoromethylornithine: Breast tissue, imaging, and serum and urine biomarkers. Clin. Cancer Res. 2002, 8, 3105–3117. [Google Scholar] [PubMed]
- Lao, C.D.; Backoff, P.; Shotland, L.I.; McCarty, D.; Eaton, T.; Ondrey, F.G.; Viner, J.L.; Spechler, S.J.; Hawk, E.T.; Brenner, D.E. Irreversible ototoxicity associated with difluoromethylornithine. Cancer Epidemiol. Biomarkers Prev. 2004, 13, 1250–1252. [Google Scholar] [PubMed]
- Porter, C.W.; Bergeron, R.J. Spermidine requirement for cell proliferation in eukaryotic cells: Structural specificity and quantitation. Science 1983, 219, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Israel, M.; Zol, E.C.; Muhammad, N.; Modest, E.J. Synthesis and antitumor evaluation of the presumed cytotoxic metabolites of spermine and N,N′-bis(3-aminopropyl)nonane-1,9-diamine. J. Med. Chem. 1973, 16, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Keinänen, T.A.; Hyvönen, M.T.; Alhonen, L.; Vepsäläinen, J.; Khomutov, A.R. Selective regulation of polyamine metabolism with methylated polyamine analogues. Amino Acids 2014, 46, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Thomas, T.J.; Thomas, T.; Singh, M.; Mann, R.A. Differential effects of polyamine homologues on the prevention of DL-α-difluoromethylornithine-mediated inhibition of malignant cell growth and normal immune response. Cancer Res. 1992, 52, 1840–1847. [Google Scholar] [PubMed]
- Wang, Y.; Murray-Stewart, T.; Devereux, W.; Hacker, A.; Frydman, B.; Woster, P.M.; Casero, R.A., Jr. Properties of purified recombinant human polyamine oxidase, PAOh1/SMO. Biochem. Biophys. Res. Commun. 2003, 304, 605–611. [Google Scholar] [CrossRef]
- Sjögren, T.; Wassvik, C.M.; Snijder, A.; Aagaard, A.; Kumanomidou, T.; Barlind, L.; Kaminski, T.P.; Kashima, A.; Yokota, T.; Fjellström, O. The structure of murine N1-acetylspermine oxidase reveals molecular details of vertebrate polyamine catabolism. Biochemistry 2017, 56, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.W.; Ganis, B.; Vinson, T.; Marton, L.J.; Kramer, D.L.; Bergeron, R.J. Comparison and characterization of growth inhibition in L1210 cells by α-difluoromethylornithine, an inhibitor of ornithine decarboxylase, and N1,N8-bis(ethyl)spermidine, an apparent regulator of the enzyme. Cancer Res. 1986, 46, 6279–6285. [Google Scholar] [PubMed]
- Porter, CW.; Bergeron, R.J. Regulation of polyamine biosynthetic activity by spermidine and spermine analogs—A novel antiproliferative strategy. Adv. Exp. Med. Biol. 1988, 250, 677–690. [Google Scholar] [PubMed]
- Bernacki, R.J.; Bergeron, R.J.; Porter, C.W. Antitumor activity of N,N′-bis(ethyl)spermine homologues against human MALME-3 melanoma xenografts. Cancer Res. 1992, 52, 2424–2430. [Google Scholar] [PubMed]
- Bergeron, R.J.; Neims, A.H.; McManis, J.S.; Hawthorne, T.R.; Vinson, J.R.; Bortell, R.; Ingeno, M.J. Synthetic polyamine analogues as antineoplastics. J. Med. Chem. 1988, 31, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.V.; Goodwin, A.C.; Hacker-Prietz, A.; Sugar, E.; Woster, P.M.; Casero, R.A., Jr. Knockdown of ornithine decarboxylase antizyme 1 causes loss of uptake regulation leading to increased N1,N11-bis(ethyl)norspermine (BENSpm) accumulation and toxicity in NCI H157 lung cancer cells. Amino Acids 2012, 42, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Pledgie, A.; Casero, R.A., Jr.; Davidson, N.E. Molecular mechanisms of polyamine analogs in cancer cells. Anticancer Drugs 2005, 16, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Seiler, N. Thirty years of polyamine-related approaches to cancer therapy. Retrospect and prospect. Part 2. Structural analogues and derivatives. Curr. Drug Targets 2003, 4, 565–585. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.E.; Mank, A.R.; Prestigiacomo, L.J.; Bergeron, R.J.; Casero, R.A., Jr. Growth inhibition of hormone-responsive and -resistant human breast cancer cells in culture by N1,N12-bis(ethyl)spermine. Cancer Res. 1993, 53, 2071–2075. [Google Scholar] [PubMed]
- Faaland, C.A.; Thomas, T.J.; Balabhadrapathruni, S.; Langer, T.; Mian, S.; Shirahata, A.; Gallo, M.A.; Thomas, T. Molecular correlates of the action of bis(ethyl)polyamines in breast cancer cell growth inhibition and apoptosis. Biochem. Cell Biol. 2000, 78, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Manni, A.; Wecher, R.; Verderame, M.F.; Mauger, D. Cooperativity between the polyamine pathway and HER2neu in transformation of human mammary epithelial cells in culture: Role of the MAPK pathway. Int. J. Cancer 1998, 76, 563–570. [Google Scholar] [CrossRef]
- Guy, C.T.; Webster, M.A.; Schaller, M.; Parsons, T.J.; Cardiff, R.D.; Muller, W.J. Expression of the neu protooncogene in the mammary epithelium of transgenic mice induces metastatic disease. Proc. Natl. Acad. Sci. USA 1992, 89, 10578–10582. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Antony, T.; Haddad, S.; Amenta, P.; Shirahata, A.; Thomas, T.J.; Thomas, T. Antitumor effects of bis(ethyl)polyamine analogs on mammary tumor development in FVB/NTgN (MMTVneu) transgenic mice. Cancer Lett. 1999, 146, 15–23. [Google Scholar] [CrossRef]
- Wolff, A.C.; Armstrong, D.K.; Fetting, J.H.; Carducci, M.K.; Riley, C.D.; Bender, J.F.; Casero, R.A., Jr.; Davidson, N.E. A Phase II study of the polyamine analog N1,N11-diethylnorspermine (DENSpm) daily for five days every 21 days in patients with previously treated metastatic breast cancer. Clin. Cancer Res. 2003, 9, 5922–5928. [Google Scholar] [PubMed]
- Balabhadrapathruni, S.; Santhakumaran, L.M.; Thomas, T.J.; Shirahata, A.; Gallo, M.A.; Thomas, T. Bis(ethyl)norspermine potentiates the apoptotic activity of the pure antiestrogen ICI 182780 in breast cancer cells. Oncol. Rep. 2005, 13, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Holst, C.M.; Staaf, J.; Jönsson, G.; Hegardt, C.; Oredsson, S.M. Molecular mechanisms underlying N1,N11-diethylnorspermine-induced apoptosis in a human breast cancer cell line. Anticancer Drugs 2008, 19, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Vijayanathan, V.; Venkiteswaran, S.; Nair, S.K.; Verma, A.; Thomas, T.J.; Zhu, B.T.; Thomas, T. Physiologic levels of 2-methoxyestradiol interfere with nongenomic signaling of 17β-estradiol in human breast cancer cells. Clin. Cancer Res. 2006, 12, 2038–2048. [Google Scholar] [CrossRef] [PubMed]
- Pledgie-Tracy, A.; Billam, M.; Hacker, A.; Sobolewski, M.D.; Woster, P.M.; Zhang, Z.; Casero, R.A.; Davidson, N.E. The role of the polyamine catabolic enzymes SSAT and SMO in the synergistic effects of standard chemotherapeutic agents with a polyamine analogue in human breast cancer cell lines. Cancer Chemother. Pharmacol. 2010, 65, 1067–1081. [Google Scholar] [CrossRef] [PubMed]
- Mahjoub, M.A.; Bakhshinejad, B.; Sadeghizadeh, M.; Babashah, S. Combination treatment with dendrosomal nanocurcumin and doxorubicin improves anticancer effects on breast cancer cells through modulating CXCR4/NF-κB/Smo regulatory network. Mol. Biol. Rep. 2017, 44, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Bunjobpol, W.; Dulloo, I.; Igarashi, K.; Concin, N.; Matsuo, K.; Sabapathy, K. Suppression of acetylpolyamine oxidase by selected AP-1 members regulates DNp73 abundance: Mechanistic insights for overcoming DNp73-mediated resistance to chemotherapeutic drugs. Cell Death Differ. 2014, 21, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Obakan, P.; Arısan, E.D.; Ozfiliz, P.; Çoker-Gurkan, A. Palavan-Unsal, N. Purvalanol A is a strong apoptotic inducer via activating polyamine catabolic pathway in MCF-7 estrogen receptor positive breast cancer cells. Mol. Biol. Rep. 2014, 41, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Pledgie, A.; Huang, Y.; Hacker, A.; Zhang, Z.; Woster, P.M.; Davidson, N.E.; Casero, R.A., Jr. Spermine oxidase SMO(PAOh1), Not N1-acetylpolyamine oxidase PAO, is the primary source of cytotoxic H2O2 in polyamine analogue-treated human breast cancer cell lines. J. Biol. Chem. 2005, 280, 39843–39851. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.K.; Verma, A.; Thomas, T.J.; Chou, T.C.; Gallo, M.A.; Shirahata, A.; Thomas, T. Synergistic apoptosis of MCF-7 breast cancer cells by 2-methoxyestradiol and bis(ethyl)norspermine. Cancer Lett. 2007, 250, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.M.; Fiuza, SM.; Marques, M.P.; Persson, L.; Oredsson, S. Increased breast cancer cell toxicity by palladination of the polyamine analogue N1N11-bis(ethyl)norspermine. Amino Acids 2014, 46, 339–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, T.J.; Shah, N.; Faaland, C.A.; Gallo, M.A.; Yurkow, E.; Satyaswaroop, P.G.; Thomas, T. Effects of a bis(benzyl)spermine analog on MCF-7 breast cancer cells in culture and nude mice xenografts. Oncol. Rep. 1977, 4, 5–13. [Google Scholar] [CrossRef]
- Casero, R.A., Jr.; Woster, P.M. Recent advances in the development of polyamine analogues as antitumor agents. J. Med. Chem. 2009, 52, 4551–4573. [Google Scholar] [CrossRef] [PubMed]
- Jagu, E.; Pomel, S.; Pethe, S.; Loiseau, P.M.; Labruère, R. Polyamine-based analogs and conjugates as antikinetoplastid agents. Eur. J. Med. Chem. 2017, 139, 982–1015. [Google Scholar] [CrossRef] [PubMed]
- Valasinas, A.; Reddy, V.K.; Blokhin, A.V.; Basu, H.S.; Bhattacharya, S.; Sarkar, A.; Marton, L.J.; Frydman, B. Long-chain polyamines (oligoamines) exhibit strong cytotoxicities against human prostate cancer cells. Bioorg. Med. Chem. 2003, 11, 4121–4131. [Google Scholar] [CrossRef]
- Huang, Y.; Keen, J.C.; Hager, E.; Smith, R.; Hacker, A.; Frydman, B.; Valasinas, A.L.; Reddy, V.K.; Marton, L.J.; Casero, R.A., Jr.; et al. Regulation of polyamine analogue cytotoxicity by c-Jun in human MDA-MB-435 cancer cells. Mol. Cancer Res. 2004, 2, 81–88. [Google Scholar] [PubMed]
- Zhu, Q.; Huang, Y.; Marton, L.J.; Woster, P.M.; Davidson, N.E.; Casero, R.A., Jr. Polyamine analogs modulate gene expression by inhibiting lysine-specific demethylase 1 (LSD1) and altering chromatin structure in human breast cancer cells. Amino Acids 2012, 42, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Pledgie, A.; Rubin, E.; Marton, L.J.; Woster, P.M.; Sukumar, S.; Casero, R.A., Jr.; Davidson, N.E. Role of p53/p21(Waf1/Cip1) in the regulation of polyamine analogue-induced growth inhibition and cell death in human breast cancer cells. Cancer Biol. Ther. 2005, 4, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Vujcic, S.; Halmekyto, M.; Diegelman, P.; Gan, G.; Kramer, D.L.; Janne, J.; Porter, C.W. Effects of conditional overexpression of spermidine/spermine N1-acetyltransferase on polyamine pool dynamics, cell growth, and sensitivity to polyamine analogs. J. Biol. Chem. 2000, 275, 38319–38328. [Google Scholar] [CrossRef] [PubMed]
- Akyol, Z.; Çoker-Gürkan, A.; Arisan, E.D.; Obakan-Yerlikaya, P.; Palavan-Ünsal, N. DENSpm overcame Bcl-2 mediated resistance against Paclitaxel treatment in MCF-7 breast cancer cells via activating polyamine catabolic machinery. Biomed. Pharmacother. 2016, 84, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.C.; Woster, P.M.; Yager, J.D.; Casero, R.A., Jr. The role of polyamine catabolism in polyamine analogue-induced programmed cell death. Proc. Natl. Acad. Sci. USA 1997, 94, 11557–11562. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feith, D.J.; Welsh, P.; Coleman, C.S.; Lopez, C.; Woster, P.M.; O’Brien, T.G.; Pegg, A.E. Studies of the mechanism by which increased spermidine/spermine N1-acetyltransferase activity increases susceptibility to skin carcinogenesis. Carcinogenesis 2007, 28, 2404–2411. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.C.; Jadallah, S.; Toubaji, A.; Lecksell, K.; Hicks, J.L.; Kowalski, J.; Bova, G.S.; De Marzo, A.M.; Netto, G.J.; Casero, R.A., Jr. Increased spermine oxidase expression in human prostate cancer and prostatic intraepithelial neoplasia tissues. Prostate 2008, 68, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Greene, E.; Murray Stewart, T.; Goodwin, A.C.; Baylin, S.B.; Woster, P.M.; Casero, R.A., Jr. Inhibition of lysine-specific demethylase 1 by polyamine analogues results in reexpression of aberrantly silenced genes. Proc. Natl. Acad. Sci. USA 2007, 104, 8023–8028. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Casero, R.A., Jr. Mammalian polyamine catabolism: A therapeutic target, a pathological problem, or both? J. Biochem. 2006, 139, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Chang, H.; Li, H.; Wang, S. Induction of reactive oxygen species: An emerging approach for cancer therapy. Apoptosis 2017, 22, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.M.; Thomas, T.; Wada, M.; Sigal, L.H.; Shirahata, A.; Thomas, T.J. Facilitation of the cellular uptake of a triplex-forming oligonucleotide by novel polyamine analogues: Structure-activity relationships. Biochemistry 1999, 38, 13328–13337. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Murray-Stewart, T.; Wang, Y.; Yu, F.; Li, J.; Marton, L.J.; Casero, R.A., Jr.; Oupický, D. Self-immolative nanoparticles for simultaneous delivery of microRNA and targeting of polyamine metabolism in combination cancer therapy. J. Control. Release 2017, 246, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Ucal, S.; Häkkinen, M.R.; Alanne, A.L.; Alhonen, L.; Vepsäläinen, J.; Keinänen, T.A.; Hyvönen, M.T. Controlling of N-Alkylpolyamine Analogue Metabolism by Selective Deuteration. Biochem. J. 2018, 475, 663–676. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, T.J.; Thomas, T. Cellular and Animal Model Studies on the Growth Inhibitory Effects of Polyamine Analogues on Breast Cancer. Med. Sci. 2018, 6, 24. https://doi.org/10.3390/medsci6010024
Thomas TJ, Thomas T. Cellular and Animal Model Studies on the Growth Inhibitory Effects of Polyamine Analogues on Breast Cancer. Medical Sciences. 2018; 6(1):24. https://doi.org/10.3390/medsci6010024
Chicago/Turabian StyleThomas, T. J., and Thresia Thomas. 2018. "Cellular and Animal Model Studies on the Growth Inhibitory Effects of Polyamine Analogues on Breast Cancer" Medical Sciences 6, no. 1: 24. https://doi.org/10.3390/medsci6010024
APA StyleThomas, T. J., & Thomas, T. (2018). Cellular and Animal Model Studies on the Growth Inhibitory Effects of Polyamine Analogues on Breast Cancer. Medical Sciences, 6(1), 24. https://doi.org/10.3390/medsci6010024