The Mechanism of Androgen Actions in PCOS Etiology


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Relationship between Hyperandrogenism and the Pathophysiology of PCOS
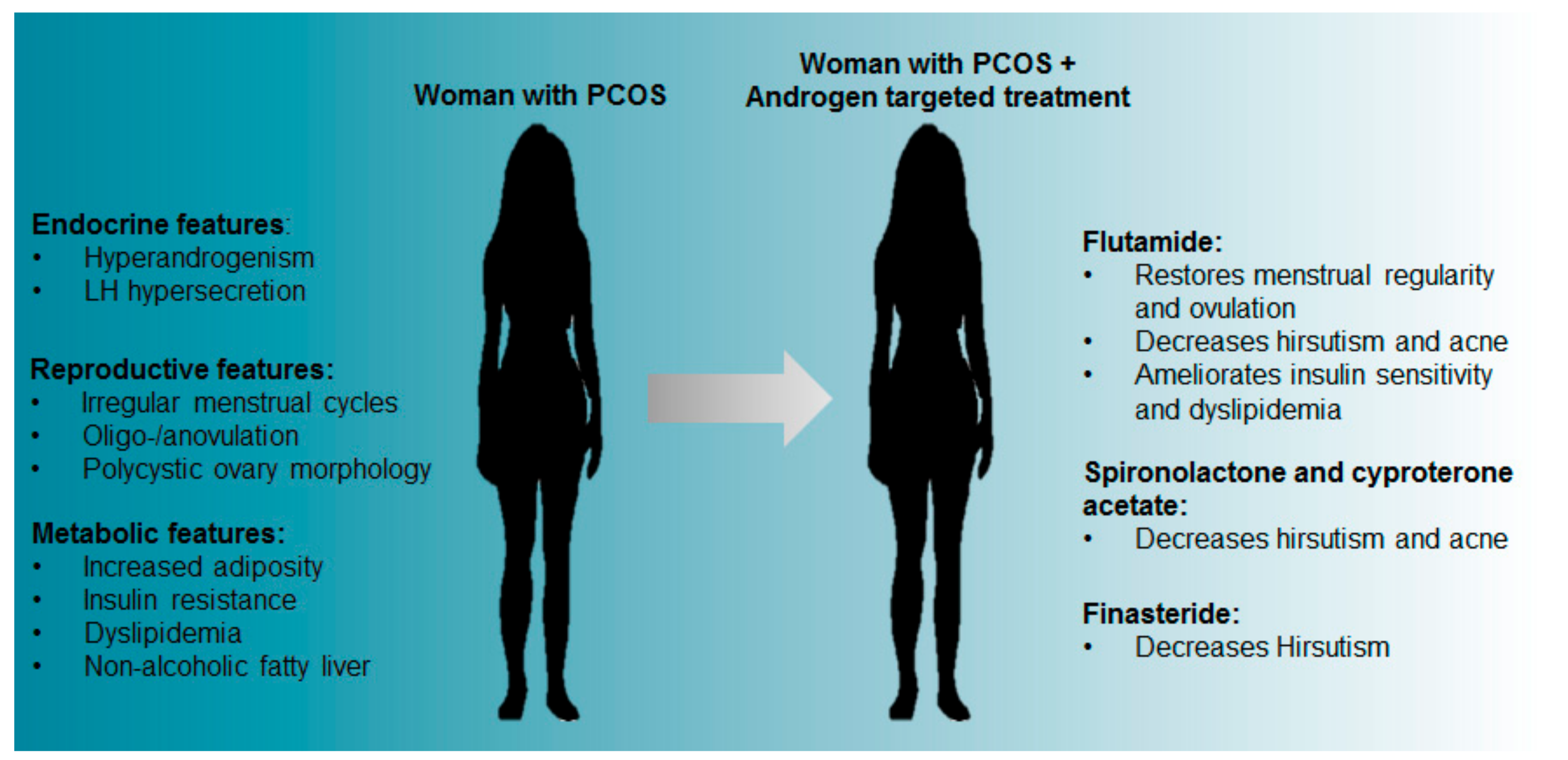
3. Clinical Targeting of Androgen Excess—Potential for Mitigating Against PCOS
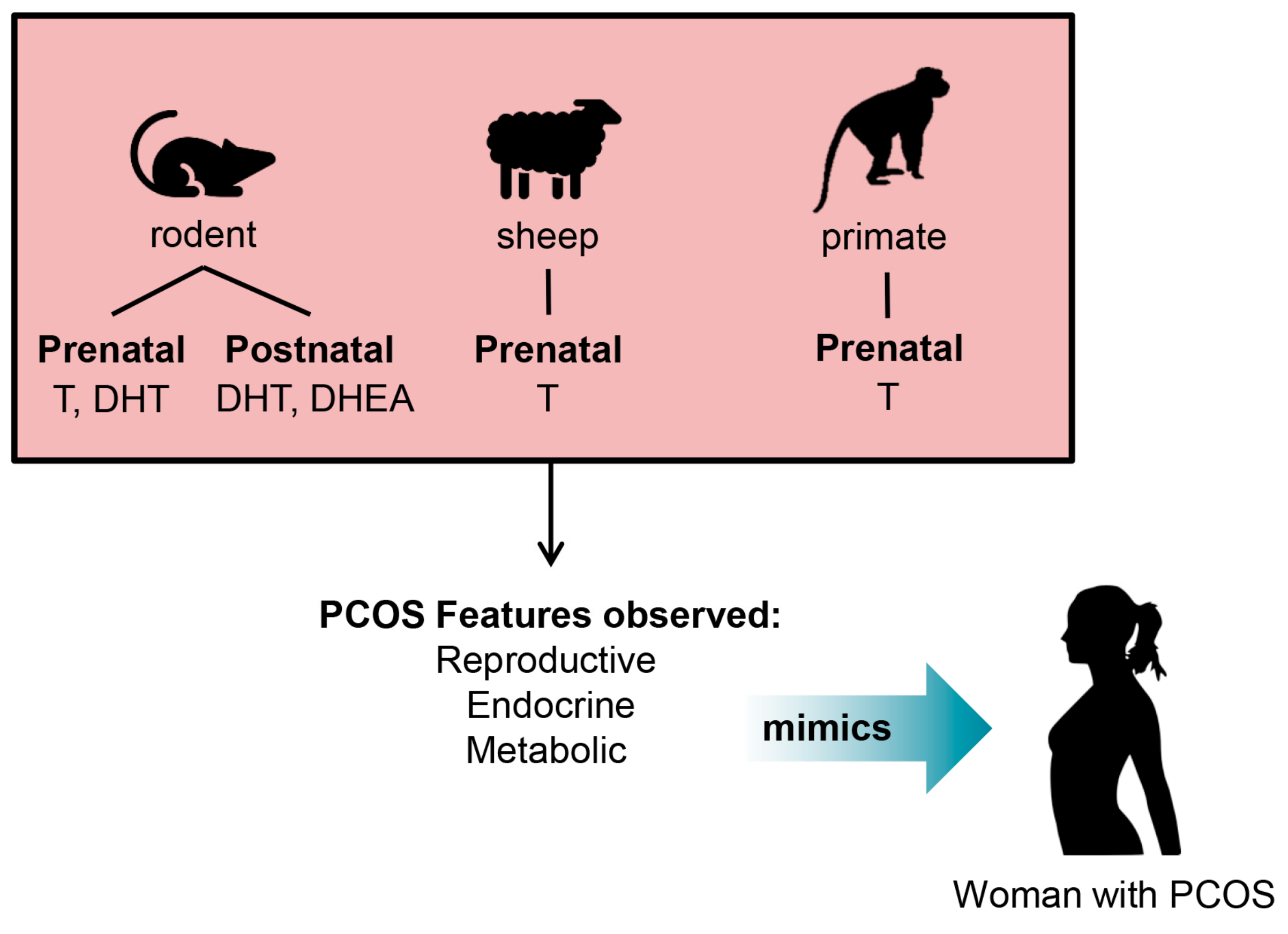
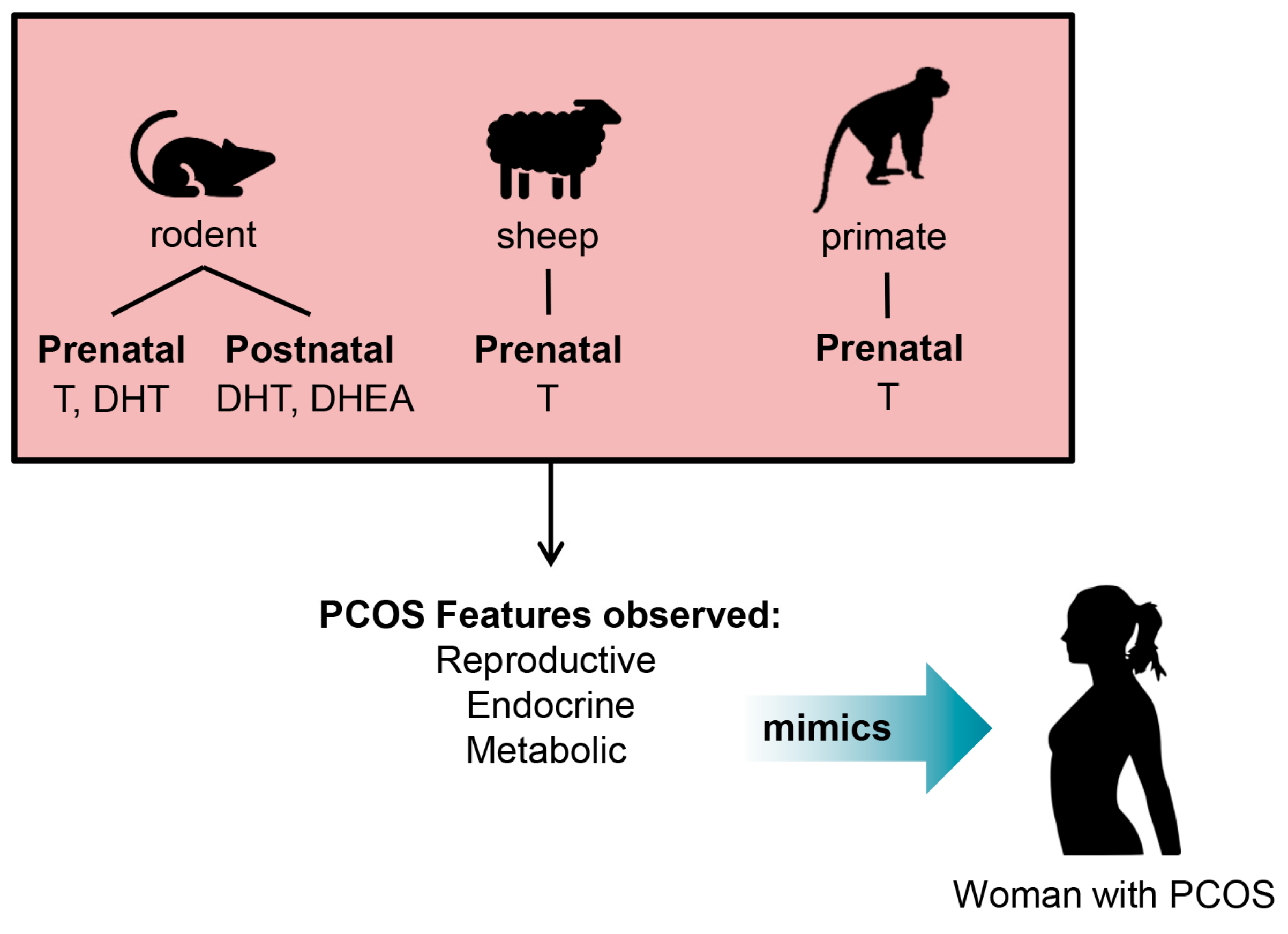
4. Development of Pre-Clinical Animal Models of PCOS
5. Insights into the Origin of PCOS from Studies in Pre-Clinical PCOS Animal Models
6. Translation of Basic Research in PCOS for the Development of Androgen-Targeted Interventions
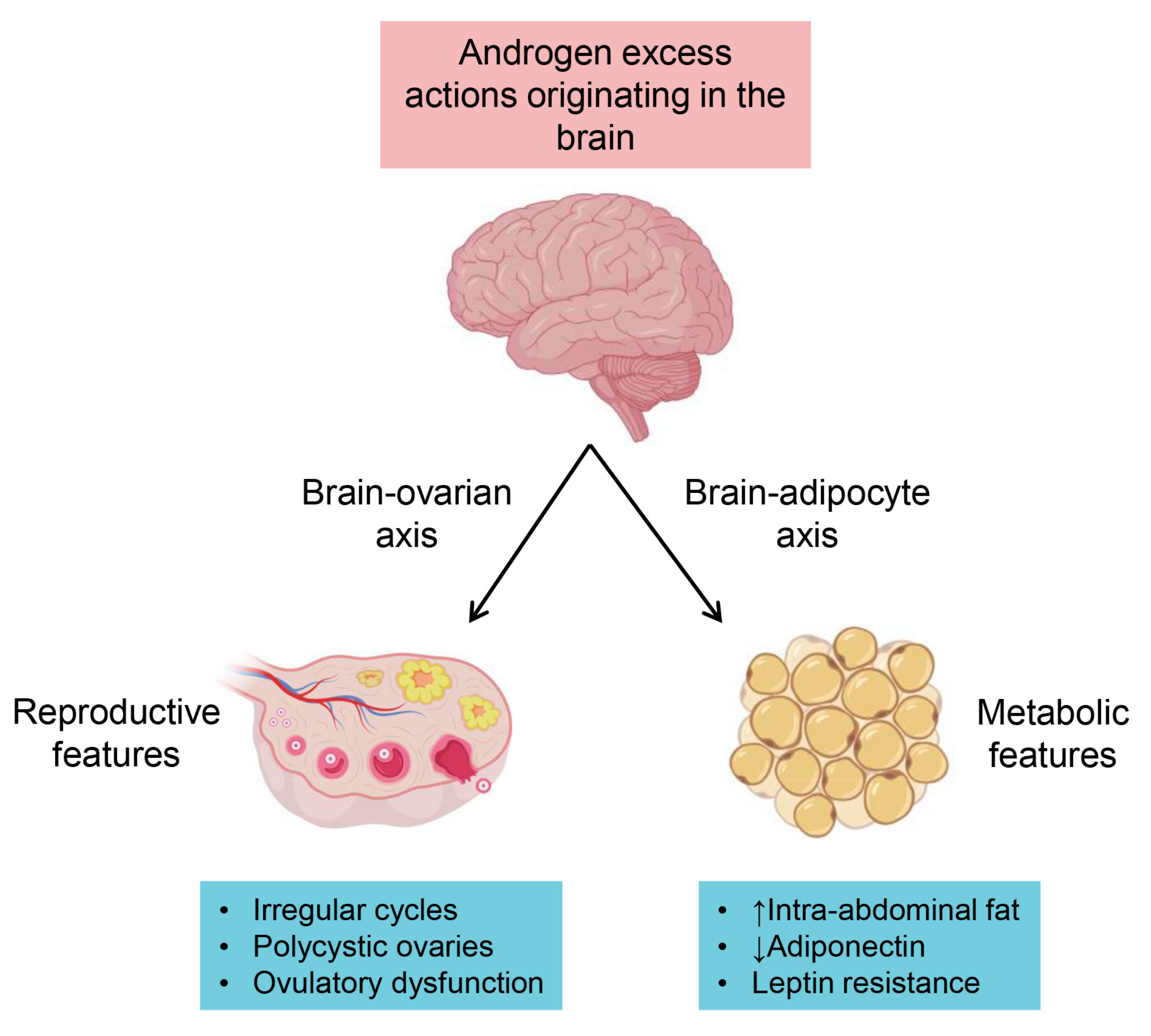
6.1. Neuroendocrine Pathways
6.2. Metabolic Pathways
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- March, W.A.; Moore, V.M.; Willson, K.J.; Phillips, D.I.W.; Norman, R.J.; Davies, M.J. The prevalence of polycystic ovary syndrome in a community sample assessed under contrasting diagnostic criteria. Hum. Reprod. 2010, 25, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Oberfield, S.E.; Stener-Victorin, E.; Marshall, J.C.; Laven, J.S.; Legro, R.S. Scientific Statement on the Diagnostic Criteria, Epidemiology, Pathophysiology, and Molecular Genetics of Polycystic Ovary Syndrome. Endocr. Rev. 2015, 36, 487–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J.; The International PCOS Network. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Clin. Endocrinol. 2018, 89, 251–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dokras, A.; Saini, S.; Gibson-Helm, M.; Schulkin, J.; Cooney, L.; Teede, H. Gaps in knowledge among physicians regarding diagnostic criteria and management of polycystic ovary syndrome. Fertil. Steril. 2017, 107, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.A.; Teede, H.J. PCOS: Refining diagnostic features in PCOS to optimize health outcomes. Nat. Rev. Endocrinol. 2016, 12, 630–631. [Google Scholar] [CrossRef]
- Joham, A.E.; Palomba, S.; Hart, R. Polycystic Ovary Syndrome, Obesity, and Pregnancy. Semin. Reprod. Med. 2016, 34, 93–101. [Google Scholar] [CrossRef]
- Doherty, D.A.P.; Newnham, J.P.; Bower, C.; Hart, R. Implications of Polycystic Ovary Syndrome for Pregnancy and for the Health of Offspring. Obstet. Gynecol. 2015, 125, 1397–1406. [Google Scholar] [CrossRef]
- Shorakae, S.; Boyle, J.; Teede, H. Polycystic ovary syndrome: A common hormonal condition with major metabolic sequelae that physicians should know about. Intern. Med. J. 2014, 44, 720–726. [Google Scholar] [CrossRef]
- Livadas, S.; Pappas, C.; Karachalios, A.; Marinakis, E.; Tolia, N.; Drakou, M.; Kaldrymides, P.; Panidis, D.; Diamanti-Kandarakis, E. Prevalence and impact of hyperandrogenemia in 1218 women with polycystic ovary syndrome. Endocrine 2014, 47, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Keefe, C.C.; Goldman, M.M.; Zhang, K.; Clarke, N.; Reitz, R.E.; Welt, C.K. Simultaneous Measurement of Thirteen Steroid Hormones in Women with Polycystic Ovary Syndrome and Control Women Using Liquid Chromatography-Tandem Mass Spectrometry. PLoS ONE 2014, 9, e93805. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Falbo, A.; Chiossi, G.; Muscogiuri, G.; Fornaciari, E.; Orio, F.; Tolino, A.; Colao, A.; La Sala, G.B.; Zullo, F. Lipid profile in nonobese pregnant women with polycystic ovary syndrome: A prospective controlled clinical study. Steroids 2014, 88, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, M.A.; Russo, G.T.; Pedone, A.; Pizzo, A.; Borrielli, I.; Stabile, G.; Artenisio, A.C.; Amato, A.; Calvani, M.; Cucinotta, D.; et al. Very High Frequency of the Polymorphism for the Insulin Receptor Substrate 1 (IRS-1) at Codon 972 (Glycine972Arginine) in Southern Italian Women with Polycystic Ovary Syndrome. Horm. Metab. Res. 2010, 42, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, M.A.; Vita, R.; Di Bari, F.; Le, D.M.; Trimarchi, F.; Benvenga, S. Gly972Arg of IRS-1 and Lys121Gln of PC-1 polymorphisms act in opposite way in polycystic ovary syndrome. J. Endocrinol. Investig. 2017, 40, 367–376. [Google Scholar] [CrossRef]
- Hague, W.M.; Adams, J.; Rodda, C.; Brook, C.G.D.; Bruyn, R.; Grant, D.B.; Jacobs, H.S. The prevalence of polycystic ovaries in patients with congenital adrenal hyperplasia and their close relatives. Clin. Endocrinol. 1990, 33, 501–510. [Google Scholar] [CrossRef]
- Spinder, T.; Spijkstra, J.J.; Tweel, J.G.V.D.; Burger, C.W.; Van Kessel, H.; Hompes, P.G.A.; Gooren, L.J.G. The Effects of Long Term Testosterone Administration on Pulsatile Luteinizing Hormone Secretion and on Ovarian Histology in Eugonadal Female to Male Transsexual Subjects. J. Clin. Endocrinol. Metab. 1989, 69, 151–157. [Google Scholar] [CrossRef]
- Pache, T.D.; Fauser, B.C. Polycystic ovaries in female-to-male transsexuals. Clin. Endocrinol. 1993, 39, 702–703. [Google Scholar]
- Nelson, V.L.; Legro, R.S.; Strauss, J.F.; McAllister, J.M. Augmented Androgen Production Is a Stable Steroidogenic Phenotype of Propagated Theca Cells from Polycystic Ovaries. Mol. Endocrinol. 1999, 13, 946–957. [Google Scholar] [CrossRef]
- Zimmerman, Y.; Eijkemans, M.J.C.; Coelingh Bennink, H.J.T.; Blankenstein, M.A.; Fauser, B.C.J.M. The effect of combined oral contraception on testosterone levels in healthy women: A systematic review and meta-analysis. Hum. Reprod. Updat. 2014, 20, 76–105. [Google Scholar] [CrossRef]
- Conway, G.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Franks, S.; Gambineri, A.; Kelestimur, F.; Macut, D.; Micic, D.; Pasquali, R.; et al. The polycystic ovary syndrome: A position statement from the European Society of Endocrinology. Eur. J. Endocrinol. 2014, 171, P1–P29. [Google Scholar] [CrossRef] [PubMed]
- Legro, R.S.; Arslanian, S.A.; Ehrmann, D.A.; Hoeger, K.M.; Murad, M.H.; Pasquali, R.; Welt, C.K. Diagnosis and Treatment of Polycystic Ovary Syndrome: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2013, 98, 4565–4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildiz, B.O. Oral contraceptives in polycystic ovary syndrome: Risk-benefit assessment. Semin. Reprod. Med. 2008, 26, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, N.; Simoncini, T.; Genazzani, A.D. Hormonal contraceptive choice for women with PCOS: A systematic review of randomized trials and observational studies. Gynecol. Endocrinol. 2014, 30, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Moghetti, P.; Tosi, F.; Tosti, A.; Negri, C.; Misciali, C.; Perrone, F.; Caputo, M.; Muggeo, M.; Castello, R. Comparison of Spironolactone, Flutamide, and Finasteride Efficacy in the Treatment of Hirsutism: A Randomized, Double Blind, Placebo-Controlled Trial 1. J. Clin. Endocrinol. Metab. 2000, 85, 89–94. [Google Scholar] [CrossRef]
- Venturoli, S.; Marescalchi, O.; Colombo, F.M.; Macrelli, S.; Ravaioli, B.; Bagnoli, A.; Paradisi, R.; Flamigni, C. A Prospective Randomized Trial Comparing Low Dose Flutamide, Finasteride, Ketoconazole, and Cyproterone Acetate-Estrogen Regimens in the Treatment of Hirsutism. J. Clin. Endocrinol. Metab. 1999, 84, 1304–1310. [Google Scholar] [CrossRef]
- Calaf, J.; López, E.; Millet, A.; Alcañiz, J.; Fortuny, A.; Vidal, O.; Callejo, J.; Escobar-Jiménez, F.; Torres, E.; Espinós, J.J. Long-Term Efficacy and Tolerability of Flutamide Combined with Oral Contraception in Moderate to Severe Hirsutism: A 12-Month, Double-Blind, Parallel Clinical Trial. J. Clin. Endocrinol. Metab. 2007, 92, 3446–3452. [Google Scholar] [CrossRef] [Green Version]
- De Leo, V.; Lanzetta, D.; D’Antona, D.; La Marca, A.; Morgante, G. Hormonal Effects of Flutamide in Young Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1998, 83, 99–102. [Google Scholar] [CrossRef]
- Paradisi, R.; Fabbri, R.; Battaglia, C.; Venturoli, S. Ovulatory effects of flutamide in the polycystic ovary syndrome. Gynecol. Endocrinol. 2013, 29, 391–395. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Mitrakou, A.; Raptis, S.; Tolis, G.; Duleba, A.J. The Effect of a Pure Antiandrogen Receptor Blocker, Flutamide, on the Lipid Profile in the Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1998, 83, 2699–2705. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Caldwell, A.S.L.; Riepsamen, A.H.; Lin, D.; Gonzalez, M.B.; Robker, R.L.; Ledger, W.L.; Gilchrist, R.B.; Handelsman, D.J.; A Walters, K. A Hyperandrogenic Environment Causes Intrinsic Defects That Are Detrimental to Follicular Dynamics in a PCOS Mouse Model. Endocrinology 2019, 160, 699–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulian, E.; Sartorato, P.; Benedini, S.; Bäro, G.; Armanini, D.; Mantero, F.; Scaroni, C. Spironolactone in the treatment of polycystic ovary syndrome: Effects on clinical features, insulin sensitivity and lipid profile. J. Endocrinol. Investig. 2005, 28, 49–53. [Google Scholar] [CrossRef]
- Tartagni, M.V.; Alrasheed, H.; Damiani, G.R.; Montagnani, M.; De Salvia, M.A.; De Pergola, G.; Tartagni, M.; Loverro, G. Intermittent Low-Dose Finasteride Administration Is Effective for Treatment of Hirsutism in Adolescent Girls: A Pilot Study. J. Pediatr. Adolesc. Gynecol. 2014, 27, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Lakryc, E.M.; Motta, E.L.A.; Soares, J.M.; Haidar, M.A.; De Lima, G.R.; Baracat, E.C.; Soares, J.M., Jr. The benefits of finasteride for hirsute women with polycystic ovary syndrome or idiopathic hirsutism. Gynecol. Endocrinol. 2003, 17, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.A.; Allan, C.M.; Handelsman, D.J. Rodent Models for Human Polycystic Ovary Syndrome. Biol. Reprod. 2012, 86, 149. [Google Scholar] [CrossRef]
- Walters, K.A. Role of androgens in normal and pathological ovarian function. Reproduction 2015, 149, R193–R218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, V.; Veiga-Lopez, A. Sheep models of polycystic ovary syndrome phenotype. Mol. Cell. Endocrinol. 2013, 373, 8–20. [Google Scholar] [CrossRef]
- Abbott, D.H.; Nicol, L.E.; Levine, J.E.; Xu, N.; Goodarzi, M.O.; Dumesic, D.A. Nonhuman primate models of polycystic ovary syndrome. Mol. Cell. Endocrinol. 2013, 373, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Walters, K.; Bertoldo, M.; Handelsman, D. Evidence from animal models on the pathogenesis of PCOS. Best Pr. Res. Clin. Endocrinol. Metab. 2018, 32, 271–281. [Google Scholar] [CrossRef]
- Moore, A.M.; Prescott, M.; Campbell, R. Estradiol Negative and Positive Feedback in a Prenatal Androgen-Induced Mouse Model of Polycystic Ovarian Syndrome. Endocrinology 2013, 154, 796–806. [Google Scholar] [CrossRef]
- Xita, N.; Tsatsoulis, A. Fetal Programming of Polycystic Ovary Syndrome by Androgen Excess: Evidence from Experimental, Clinical, and Genetic Association Studies. J. Clin. Endocrinol. Metab. 2006, 91, 1660–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippou, P.; Homburg, R. Is foetal hyperexposure to androgens a cause of PCOS? Hum. Reprod. Updat. 2017, 23, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Nohara, K.; Laque, A.; Allard, C.; Münzberg, H.; Mauvais-Jarvis, F. Central mechanisms of adiposity in adult female mice with androgen excess. Obesity 2014, 22, 1477–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, A.S.L.; Middleton, L.J.; Jimenez, M.; Desai, R.; McMahon, A.C.; Allan, C.M.; Handelsman, D.J.; Walters, K.A. Characterization of Reproductive, Metabolic, and Endocrine Features of Polycystic Ovary Syndrome in Female Hyperandrogenic Mouse Models. Endocrinology 2014, 155, 3146–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, A.S.L.; Edwards, M.C.; Desai, R.; Jimenez, M.; Gilchrist, R.B.; Handelsman, D.J.; Walters, K.A. Neuroendocrine androgen action is a key extraovarian mediator in the development of polycystic ovary syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, E3334–E3343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, A.S.; Thackray, V.G.; Ryan, G.E.; Tolson, K.P.; Glidewell-Kenney, C.A.; Semaan, S.J.; Poling, M.C.; Iwata, N.; Breen, K.M.; Duleba, A.J.; et al. A Novel Letrozole Model Recapitulates Both the Reproductive and Metabolic Phenotypes of Polycystic Ovary Syndrome in Female Mice1. Biol. Reprod. 2015, 93, 69. [Google Scholar] [CrossRef]
- Leonie, E.; Kramer, P.; McLuskey, A.; Karels, B.; Themmen, A.P.N.; Visser, J.A. Reproductive and Metabolic Phenotype of a Mouse Model of PCOS. Endocrinology 2012, 153, 2861–2869. [Google Scholar] [Green Version]
- Keller, E.; Chazenbalk, G.D.; Aguilera, P.; Madrigal, V.; Grogan, T.; Elashoff, D.; Dumesic, D.A.; Abbott, D.H. Impaired Preadipocyte Differentiation into Adipocytes in Subcutaneous Abdominal Adipose of PCOS-Like Female Rhesus Monkeys. Endocrinology 2014, 155, 2696–2703. [Google Scholar] [CrossRef]
- Padmanabhan, V.; Veiga-Lopez, A.; Herkimer, C.; Salloum, B.A.; Moeller, J.; Beckett, E.; Sreedharan, R. Developmental Programming: Prenatal and Postnatal Androgen Antagonist and Insulin Sensitizer Interventions Prevent Advancement of Puberty and Improve LH Surge Dynamics in Prenatal Testosterone-Treated Sheep. Endocrinology 2015, 156, 2678–2692. [Google Scholar] [CrossRef]
- E Ryan, G.; Malik, S.; Mellon, P.L. Antiandrogen Treatment Ameliorates Reproductive and Metabolic Phenotypes in the Letrozole-Induced Mouse Model of PCOS. Endocrinology 2018, 159, 1734–1747. [Google Scholar] [CrossRef]
- Sullivan, S.D.; Moenter, S.M. Prenatal androgens alter GABAergic drive to gonadotropin-releasing hormone neurons: Implications for a common fertility disorder. Proc. Natl. Acad. Sci. USA 2004, 101, 7129–7134. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, K.M.; Padmanabhan, V.; Coolen, L.M.; Lehman, M.N. Prenatal Programming by Testosterone of Hypothalamic Metabolic Control Neurones in the Ewe. J. Neuroendocr. 2011, 23, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, A.; Eid, S.; Kay, C.; Jimenez, M.; McMahon, A.; Desai, R.; Allan, C.; Smith, J.; Handelsman, D.; Walters, K.A. Haplosufficient Genomic Androgen Receptor Signaling Is Adequate to Protect Female Mice from Induction of Polycystic Ovary Syndrome Features by Prenatal Hyperandrogenization. Endocrinology 2015, 156, 1441–1452. [Google Scholar] [CrossRef]
- Ma, Y.; Andrisse, S.; Chen, Y.; Childress, S.; Xue, P.; Wang, Z.; Jones, D.; Ko, C.M.; Divall, S.; Wu, S. Androgen Receptor in the Ovary Theca Cells Plays a Critical Role in Androgen-Induced Reproductive Dysfunction. Endocrinology 2017, 158, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.H. Neuronal androgen receptor: Molecular gateway to polycystic ovary syndrome? Proc. Natl. Acad. Sci. USA 2017, 114, 4045–4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.-Y.; Li, Z.-L.; Wu, C.-Y.; Liu, Y.-M.; Lin, H.; Wang, S.-H.; Xiao, W.-F. Endocrine traits of polycystic ovary syndrome in prenatally androgenized female Sprague-Dawley rats. Endocr. J. 2010, 57, 201–209. [Google Scholar] [CrossRef]
- Moore, A.M.; Prescott, M.; Marshall, C.J.; Yip, S.H.; Campbell, R.E. Enhancement of a robust arcuate GABAergic input to gonadotropin-releasing hormone neurons in a model of polycystic ovarian syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, 596–601. [Google Scholar] [CrossRef]
- Sarma, H.N.; Manikkam, M.; Herkimer, C.; Dell’Orco, J.; Welch, K.B.; Foster, D.L.; Padmanabhan, V. Fetal Programming: Excess Prenatal Testosterone Reduces Postnatal Luteinizing Hormone, But Not Follicle-Stimulating Hormone Responsiveness, to Estradiol Negative Feedback in the Female. Endocrinology 2005, 146, 4281–4291. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Harlan, R.E. Absence of androgen receptors in LHRH immunoreactive neurons. Brain Res. 1993, 624, 309–311. [Google Scholar] [CrossRef]
- Smith, J.T. Sex Steroid Regulation of Kisspeptin Circuits. Results Probl. Cell Differ. 2013, 784, 275–295. [Google Scholar]
- Navarro, V.M.; Gottsch, M.L.; Chavkin, C.; Okamura, H.; Clifton, D.K.; Steiner, R.A. Regulation of gonadotropin-releasing hormone secretion by kisspeptin/dynorphin/neurokinin B neurons in the arcuate nucleus of the mouse. J. Neurosci. 2009, 29, 11859–11866. [Google Scholar] [CrossRef] [PubMed]
- Skorupskaite, K.; George, J.T.; A Anderson, R. The kisspeptin-GnRH pathway in human reproductive health and disease. Hum. Reprod. Updat. 2014, 20, 485–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, K.A.; Edwards, M.C.; Tesic, D.; Caldwell, A.S.; Jimenez, M.; Smith, J.T.; Handelsman, D.J. The Role of Central Androgen Receptor Actions in Regulating the Hypothalamic-Pituitary-Ovarian Axis. Neuroendocrinology 2018, 106, 389–400. [Google Scholar] [CrossRef] [PubMed]
- AlBalawi, F.S.; Daghestani, M.H.; Daghestani, M.H.; Eldali, A.; Warsy, A.S. rs4889 polymorphism in KISS1 gene, its effect on polycystic ovary syndrome development and anthropometric and hormonal parameters in Saudi women. J. Biomed. Sci. 2018, 25, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umayal, B.; Jayakody, S.; Chandrasekharan, N.; Wijesundera, W.S.; Wijeyaratne, C. Polycystic ovary syndrome (PCOS) and kisspeptin—A Sri Lankan study. J. Postgrad. Med. 2019, 65, 18–23. [Google Scholar]
- Katulski, K.; Podfigurna, A.; Czyzyk, A.; Meczekalski, B.; Genazzani, A.D. Kisspeptin and LH pulsatile temporal coupling in PCOS patients. Endocrine 2018, 61, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Osuka, S.; Iwase, A.; Nakahara, T.; Kondo, M.; Saito, A.; Nakamura, T.; Takikawa, S.; Goto, M.; Kotani, T.; Kikkawa, F. Kisspeptin in the hypothalamus of 2 rat models of polycystic ovary syndrome. Endocrinology 2017, 158, 367–377. [Google Scholar] [CrossRef]
- Brown, R.E.; Wilkinson, D.A.; Imran, S.A.; Caraty, A.; Wilkinson, M. Hypothalamic kiss1 mRNA and kisspeptin immunoreactivity are reduced in a rat model of polycystic ovary syndrome (PCOS). Brain Res. 2012, 1467, 1–9. [Google Scholar] [CrossRef]
- Cernea, M.; Padmanabhan, V.; Goodman, R.L.; Coolen, L.M.; Lehman, M.N. Prenatal Testosterone Treatment Leads to Changes in the Morphology of KNDy Neurons, Their Inputs, and Projections to GnRH Cells in Female Sheep. Endocrinology 2015, 156, 3277–3291. [Google Scholar] [CrossRef] [Green Version]
- George, J.T.; Kakkar, R.; Marshall, J.; Scott, M.L.; Finkelman, R.D.; Ho, T.W.; Veldhuis, J.; Skorupskaite, K.; A Anderson, R.; McIntosh, S.; et al. Neurokinin B Receptor Antagonism in Women with Polycystic Ovary Syndrome: A Randomized, Placebo-Controlled Trial. J. Clin. Endocrinol. Metab. 2016, 101, 4313–4321. [Google Scholar] [CrossRef]
- Villanueva, E.C.; Myers, M.G., Jr. Leptin receptor signaling and the regulation of mammalian physiology. Int. J. Obes. 2008, 32, S8–S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumesic, D.A.; Akopians, A.L.; Madrigal, V.K.; Ramirez, E.; Margolis, D.J.; Sarma, M.K.; Thomas, A.M.; Grogan, T.R.; Haykal, R.; Schooler, T.A.; et al. Hyperandrogenism Accompanies Increased Intra-Abdominal Fat Storage in Normal Weight Polycystic Ovary Syndrome Women. J. Clin. Endocrinol. Metab. 2016, 101, 4178–4188. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.C.; Veiga-Lopez, A.; Moeller, J.; Beckett, E.; Pease, A.; Keller, E.; Madrigal, V.; Chazenbalk, G.; Dumesic, D.; Padmanabhan, V. Developmental Programming: Impact of Gestational Steroid and Metabolic Milieus on Adiposity and Insulin Sensitivity in Prenatal Testosterone-Treated Female Sheep. Endocrinology 2015, 157, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Mannerås-Holm, L.; Leonhardt, H.; Kullberg, J.; Jennische, E.; Odén, A.; Holm, G.; Hellström, M.; Lönn, L.; Olivecrona, G.; Stener-Victorin, E.; et al. Adipose Tissue Has Aberrant Morphology and Function in PCOS: Enlarged Adipocytes and Low Serum Adiponectin, but not Circulating Sex Steroids, Are Strongly Associated with Insulin Resistance. Endocrinology 2011, 152, 332. [Google Scholar] [CrossRef]
- Singh, A.; Bora, P.; Krishna, A. Systemic adiponectin treatment reverses polycystic ovary syndrome-like features in an animal model. Reprod. Fertil. Dev. 2018, 30, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Benrick, A.; Chanclón, B.; Micallef, P.; Wu, Y.; Hadi, L.; Shelton, J.M.; Stener-Victorin, E.; Asterholm, I.W. Adiponectin protects against development of metabolic disturbances in a PCOS mouse model. Proc. Natl. Acad. Sci. USA 2017, 114, E7187–E7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Hu, T.; Zhao, H.; Huang, Y.; Ye, R.; Lin, J.; Zhang, C.; Zhang, H.; Wei, G.; Zhou, H.; et al. Brown adipose tissue transplantation ameliorates polycystic ovary syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, 2708–2713. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Indran, I.R.; Tan, H.M.; Li, Y.; Zhang, Z.; Li, J.; Yong, E.-L. A Dietary Medium-Chain Fatty Acid, Decanoic Acid, Inhibits Recruitment of Nur77 to the HSD3B2 Promoter In Vitro and Reverses Endocrine and Metabolic Abnormalities in a Rat Model of Polycystic Ovary Syndrome. Endocrinology 2016, 157, 382–394. [Google Scholar] [CrossRef]
- Marti, N.; Bouchoucha, N.; Sauter, K.-S.; Flück, C.E. Resveratrol inhibits androgen production of human adrenocortical H295R cells by lowering CYP17 and CYP21 expression and activities. PLoS ONE 2017, 12, e0174224. [Google Scholar] [CrossRef]
- Ergenoglu, M.; Yildirim, N.; Yildirim, A.G.S.; Yeniel, O.; Erbas, O.; Yavasoglu, A.; Taskiran, D.; Karadadas, N. Effects of Resveratrol on Ovarian Morphology, Plasma Anti-Mullerian Hormone, IGF-1 Levels, and Oxidative Stress Parameters in a Rat Model of Polycystic Ovary Syndrome. Reprod. Sci. 2015, 22, 942–947. [Google Scholar] [CrossRef]
- Banaszewska, B.; Wrotyńska-Barczyńska, J.; Spaczynski, R.Z.; Pawelczyk, L.; Duleba, A.J. Effects of Resveratrol on Polycystic Ovary Syndrome: A Double-blind, Randomized, Placebo-controlled Trial. J. Clin. Endocrinol. Metab. 2016, 101, 4322–4328. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez Paris, V.; Bertoldo, M.J. The Mechanism of Androgen Actions in PCOS Etiology. Med. Sci. 2019, 7, 89. https://doi.org/10.3390/medsci7090089
Rodriguez Paris V, Bertoldo MJ. The Mechanism of Androgen Actions in PCOS Etiology. Medical Sciences. 2019; 7(9):89. https://doi.org/10.3390/medsci7090089
Chicago/Turabian StyleRodriguez Paris, Valentina, and Michael J. Bertoldo. 2019. "The Mechanism of Androgen Actions in PCOS Etiology" Medical Sciences 7, no. 9: 89. https://doi.org/10.3390/medsci7090089
APA StyleRodriguez Paris, V., & Bertoldo, M. J. (2019). The Mechanism of Androgen Actions in PCOS Etiology. Medical Sciences, 7(9), 89. https://doi.org/10.3390/medsci7090089