
The Origin of Phthalates in Algae: Biosynthesis and Environmental Bioaccumulation


1
Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, 90128 Palermo, Italy
2
National Biodiversity Future Center (NBFC), 90133 Palermo, Italy
*
Author to whom correspondence should be addressed.
Environments 2024, 11(4), 78; https://doi.org/10.3390/environments11040078
Submission received: 21 February 2024
/
Revised: 8 April 2024
/
Accepted: 10 April 2024
/
Published: 11 April 2024
Abstract
:Phthalic acid esters (PAEs) are a class of ubiquitous and dangerous lipophilic chemicals widely used as additives in various products to improve their physical and chemical properties. Although they have been banned in many countries, their persistence in all environmental compartments is of particular concern. The aquatic environment is especially affected by these compounds because it is strongly influenced both by contamination of anthropic origin and natural contaminants including those produced biosynthetically by some organisms such as algae. In this context, algal organisms can be a source and remedy for phthalate pollution. Both the increase and decrease in uptake and production depend on the physicochemical characteristics of the environment. The dynamics of the natural processes are aimed at achieving an optimal environmental state for their competitiveness and balance of the cellular homeostasis. This review summarizes the studies dealing with biosynthesis and bioaccumulation of phthalates in algae and investigates the source of their origin by suggesting strategies to identify the process leading to their presence.
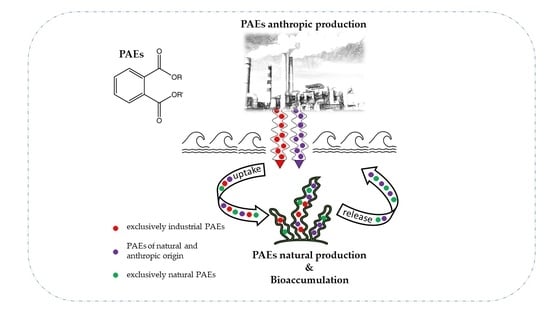
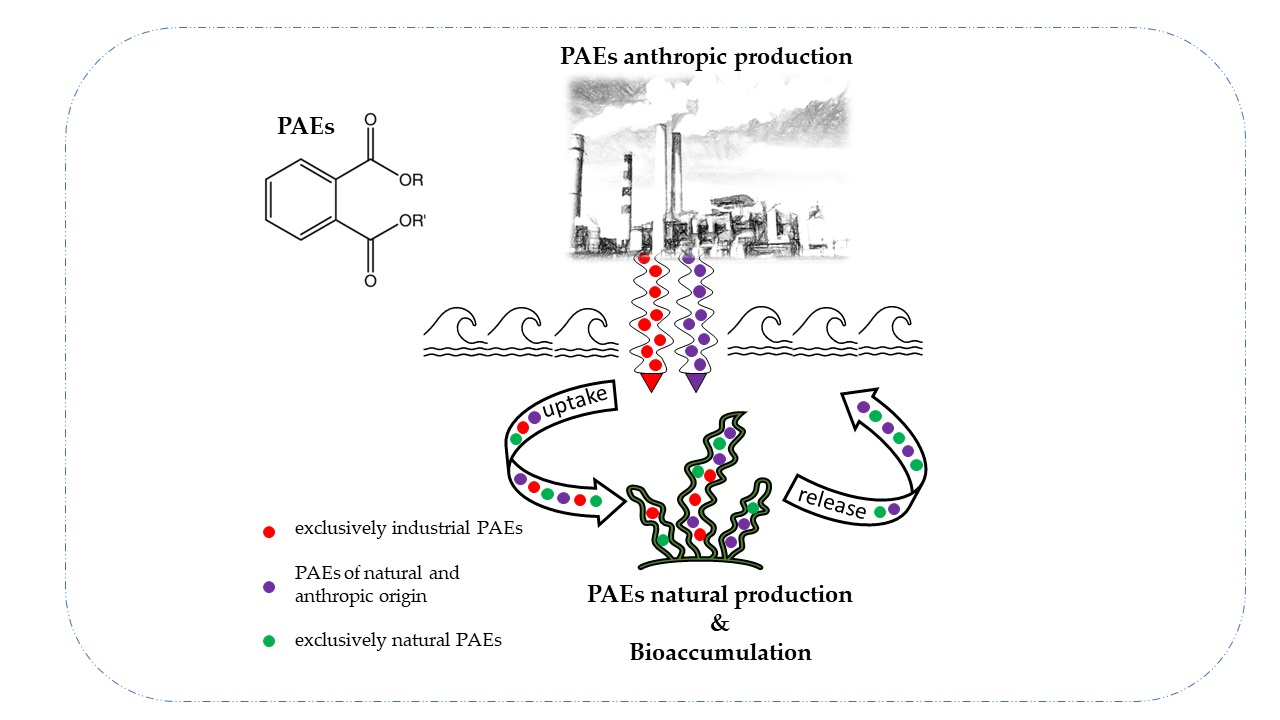
1. Introduction
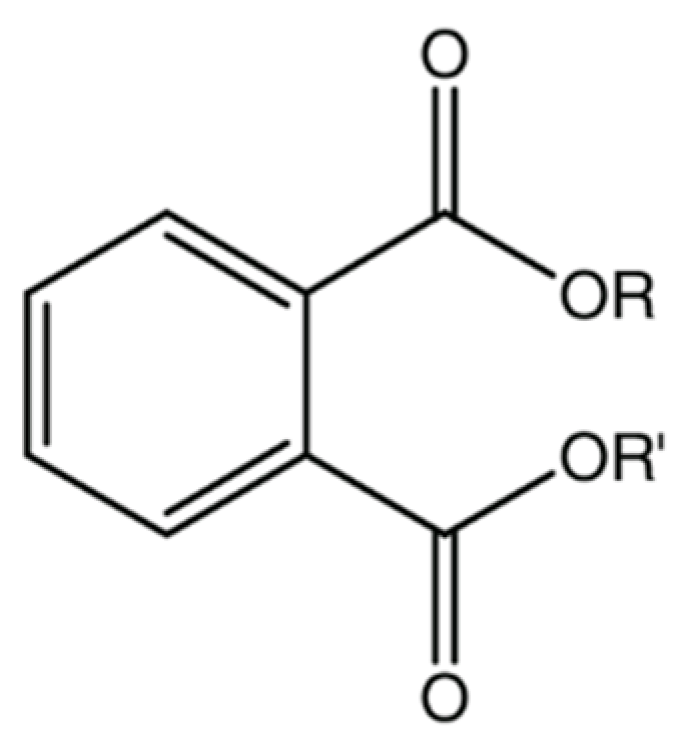
Phthalic acid esters (PAEs) are a class of lipophilic chemicals (with partition coefficient octanol/water, log Kow, ranging from 1.6 to 12) widely used since 1930s as additives in various products such as in polyvinyl chloride (PVC) products, personal care products, cosmetics, adhesives, paints, printing inks, pesticides, toys, and in many other applications from electronics to medications [1,2,3,4]. PAEs are therefore used both as plasticizers and as non-plasticizers: to improve different characteristics of various products (e.g., durability, resistance, extensibility, flexibility in polymer or persistence in perfumes, etc.) [3,4].
Figure 1 and Table 1 show general chemical structure and a short list of the most common phthalates.
PAEs are ubiquitous substances easily detected in all environmental compartments (atmosphere, water, soil, and sediments) [4].
Those shown in Table 1 are the most common phthalates. In particular, DEHP was the most used in PVC materials (up to 40% product weight and about 50% of all plasticizers used in PVC) [2]. Despite bans and restrictions on the use of phthalates, environmental and food contamination continues to persist. According to European Union (EU) law (Commission Regulation (EU) 2018/2005), the content of DEHP, DBP, BBP, and DIBP should not exceed 0.1% in toys, children’s products, and lamination products while DEHP, BBP, DBP, DIBP, DINP, DIDP, and DNOP are strictly limited in food material contacts (FCMs) and as cosmetic components [6]. In the United States, there is no formal ban on their use in cosmetics, but phthalates are mostly limited in FCMs. In China, the limit for DBP, BBP, DEHP, DNOP, DINP, and DIDP in plastic toys is 0.1% of the composition of the material; for cosmetics, DEHP, BBP, and DBP are prohibited. However, phthalates are still found in a variety of products and are a cause for concern [6,7].
The large amount of plastic pollution has led to the significant accumulation of these substances in aquatic environments. Once PAEs are released, they can be transferred through the food chain, causing detrimental effects to the health of exposed organisms and ultimately impacting the ecosystem’s functioning [4,8]. In fact, phthalates are known to be endocrine-active substances (EAS) and can cause different liver, reproductive, metabolic, and developmental diseases [4].
The sources of contamination are typically considered anthropogenic: once synthesized and added as additives, they are easily released into the environment (e.g., from plastic to the aquatic environment) [4]. When phthalates are used as monomers to produce polymers, they are covalently bonded with each other. However, when used as plasticizers, they are not covalently bonded to the polymer to which they are mixed and can be easily released [2].
In the aquatic environment, few studies have been conducted on algae, especially macroalgae. It is important to note that this lack of research is unique to algae when compared to other organisms.
Algae are the main producers of aquatic ecosystems, the starting point of the chain or trophic network, and contribute to about half of the oxygen present on the planet [9].
Macroalgae are multicellular, photosynthetic organisms that are mainly found in aquatic environments, particularly in the marine realm. They belong to the Eukarya domain and are classified into three groups: green algae, red algae, and brown algae (also known as Chlorophyta, Rhodophyta, and Phaeophyta, respectively).
Microalgae are a large group of eukaryotic, photoautotrophic organisms that include both protists and prokaryotic cyanobacteria. They are a significant component of phytoplankton. Microalgae are microscopic and can become highly visible during massive blooms, such as red tides and other harmful algal blooms. They come in a variety of sizes, structures, and shapes and are classified into more than a dozen groups based predominantly on pigment composition and ultrastructural characteristics [9,10,11].
Together with the natural presence of algae in all aquatic environments, global algae production has increased significantly today. The colonization of more than 291 species used for different purposes (e.g., food and nutraceuticals, paper, fertilizers, medicines, cosmetics, and industrial products) is increasing their abundance on the planet. In addition, various algal extracts have demonstrated numerous health benefits for humans because of the active substances they contain, which have various biological actions such as anti-obesity, anti-diabetes, anti-cancer, antimicrobial, antiviral, antioxidant, anti-inflammatory, and cardioprotective effects [12].
Due to extreme climatic and environmental stress, such as salinity, light, temperature, and marine chemical diversions, algae produce many new primary metabolites (proteins, fibers, vitamins, minerals, and polyunsaturated fatty acids) and secondary metabolites (sterols, phycoblins, terpenes, tocopherols, ficants, polyphenols, and polysaccharides) in addition to the necessary for normal growth [12].
Among these metabolites, phthalates are a cause for concern due to their frequent use in algal matrices for various purposes that involve direct exposure. At the same time, algal species can act as bioremediators in environmental bioremediation by accumulating or degrading a wide range of pollutants [8,13,14,15].
Therefore, the recorded presence of phthalates in algae could derive from an inner source of significant natural origin.
Indeed, while evidence highlights the ability of some species to adsorb or biodegrade PAEs, it is just as proven that some species of plants, algae, bacteria, fungi, etc., biosynthesize phthalates to improve their competitiveness in the environment [2,3].
Phthalates are typically biosynthesized for their ecologically relevant biological activities, such as allelopathic/phytotoxic, antimicrobial (antibacterial and antifungal), insecticidal, and antioxidant properties. They also act as inducers of apoptosis and cell cycle arrest, and have cytotoxic, antitumor, larvicidal, antifouling, chemotactic, antimelanogenic, and antiviral effects. They are generally used to defend against biotic and abiotic factors [3,16]. Several other reasons for the biosynthesis of PAEs have been observed. These reasons include the usefulness of the organism or the natural environment, such as the reduction of disease occurrence, the improvement of soil properties, and plant growth [3,16].
The undoubted importance of these organisms therefore lies both in the transfer of the biologically active substances present in them and in the ability of these organisms to mitigate or cause environmental pollution. To determine the origin of phthalates in algae and their contribution to or mitigation of phthalate pollution, further investigations are necessary. The aim of this work is to gather and analyze studies on the ability of algal organisms to biosynthesize, bioremediate the environment, and bioaccumulate phthalates. The most extensively studied phthalates are discussed in the literature, which is divided into hypotheses regarding their sources of occurrence. Strategies for identifying their origins are also suggested.
2. PAEs and Algal Matrices
Despite the possibility that phthalates in natural matrices could originate from biosynthetic pathways, many studies claim accumulation from the polluted environment as the sole source, while others leave the question unresolved. In the following section, an overview of the results and findings on the presence of PAEs in algae is provided, together with a critical analysis of the context of the study and comments on alternative hypotheses.
2.1. Principal PAEs Studies in Algae
As already mentioned, different works have found the presence of phthalates in biomonitoring studies and in chemical characterization of algal constituents but did not aim to verify their source.
For example, based on spectroscopic evidence, brown algae Stoechospermum marginatum showed the presence of DBP, DEHP, and probably DINP [17], while in brown algae Sargassum confusum were found DnBP, DINP, DEHP, and dicyclohexyl phthalate (DCHP) [18].
Most research has found PAEs in characterization studies of bioactive volatile compounds and their associated pharmacological properties, such as the study conducted on four species of macroalgae (Hydropuntia edulis, Halymenia venusta, Ulva lactuca, and Padina gymnospora) by Vijayaraj et al. [19], who reported the presence of DEP, DBP, and DIOP in one of them (Ulva lactuca). In another study, it was observed that BIBP, extracted from the rhizoids of Laminaria japonica, showed inhibitory activity against α-glucosidase suggesting its potential as an antidiabetic agent for type II diabetes therapies [20].
Ulva lactuca, Petalonia fascia, Gelidium spinosum, and Cladophora glomerata were investigated as natural sources of antibacterial and antimetabolic compounds.
In detail, the alcoholic extracts of Ulva lactuca and Cladophora glomerata showed the most significant biological activity due to the presence of DIOP and DBP respectively [21,22]. Additionally, MEHP found in acetone extract of Sargassum wightii showed an high antibacterial efficiency against different Gram-positive and Gram-negative bacterial strains [23].
In a study of characterization of volatile constituents in brown algae Padina pavonia and Hydroclathrus clathratus, different phthalates such as DnBP, DIBP, DEHP, and DnOP were found [24]. In this work, DEHP was identified as the main constituent, as well as in red algae Corallina officinalis [25]. In addition DEHP was present also in and green algae Caulerpa racemosa and Codium tomentosum [26]. In another study of essential oil composition of Nizamuddinia zanardinii, a brown alga collected from Oman Gulf, it was observed that phthalates (DEP and DBP) represented 5.9% of the chemical constituents of its volatile oil [27].
Similarly, in another study, no hypothesis was made about the PAEs’ origin; in fact, the source of DEHP, isolated from Ceramium rubrum, had not been clarified [28].
Another example is the presence of DnBP in the dichloromethane extract of green alga Ulva tepida, tested for its inductive effect on Ulva settlements and involved in the triggering of green tides [29].
DIOP was found in methanolic extracts of red algae Gelidium crinale, brown algae Sargassum hornschuchii, and green algae Ulva linza, which exhibited a high antioxidant effect [30].
Chloroform and ethanol extracts of the red algae Sarconema filiforme and Laurencia obtuse were found to contain several previously unreported PAEs, including phthalic acid, butyl tridecyl ester; phthalic acid, bis(7-methyloctyl) ester; didecan-2-yl phthalate; phthalic acid, hex-3-yl isobutyl ester; and butyl 8-chlorooctyl phthalate, as well as others already known (see Table 2). These extracts exhibited a moderate antiproliferative effect against human breast cancer cell lines (MCF-7 and MDA-MB231) as well as a human lung cancer cell line (A549) [12].
Some fractions of a dichloromethane and methanol extract of Iridea cordata containing mainly phthalates (see Table 2), showed high antileishmanial activity. In fact, these showed promising activity against amastigotes and a higher selectivity towards the parasite than towards the mammalian host cells, compared to the reference drug amphotericin B [31].
All phthalates presented in this review are ortho-phthalates and there are currently only a few papers presenting others; for example, one by Guven et al. [32] in which dimethyl terephthalate was obtained from three red algae (Phyllophora nervosa, Acanthophora delilei, and Hypnea musciformis) by extraction with different solvents.

{kind=link}
{kind=link}
Table 2.
Overview of the detection of phthalates in various species of algae in both natural environments (N.E.) and laboratory conditions (L.C.), as well as the hypothesis regarding their origin.
Table 2.
Overview of the detection of phthalates in various species of algae in both natural environments (N.E.) and laboratory conditions (L.C.), as well as the hypothesis regarding their origin.
| Species | PAEs (µg/g Dry Weight) | Context | Hypothesis of PAEs’ Origin and Reference | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DnBP | DEHP | MEHP | |||||||||||
| Bangia atropurpurea 8 | 62.14 | 34.74 | L.C. | Biosynthesis [33] | |||||||||
| Porphyra angusta 8 | 22.16 | 6.35 | L.C. | ||||||||||
| Porphyra angusta 8 | 11.67 * | 6.86 * | N.E. | ||||||||||
| Porphyra dentata 8 | 33.47 | 18.53 | L.C. | ||||||||||
| Ulva fasciata 8 | 17.56 * | 11.59 * | N.E. | ||||||||||
| Enteromorpha intestinalis 8 | 18.32 * | 15.77 * | N.E. | ||||||||||
| Cladophora fracta 7 | 77.77 ** | 354.20 | N.E. | Biosynthesis [34] | |||||||||
| Spirogyra sp. 7 | 147.83 | 339.59 ** | N.E. | ||||||||||
| Botryococcus braunii 7 | - | 191.97 | N.E. | ||||||||||
| Chlorella sp. 7 | 41.36 | 376.10 | N.E. | ||||||||||
| Hydrodictyon reticulatum 7 | 165.21 | - | N.E. | ||||||||||
| Ulva lactuca 1 | DEP, DMP, DBP, BBP, DEHP, and DnOP at concentration less than 1 µg/g | L.C. | Bioaccumulation [8] | ||||||||||
| PAEs (compositional %) | |||||||||||||
| DEP | DnBP | DIBP | DEHP | MEHP | DnOP | DIOP | |||||||
| Nizamuddinia zanardinii 9 | 0.70 | 5.10 | N.E. | Biosynthesis [27] | |||||||||
| Padina pavonica 9 | 0.92 | 1.26 | 19.75 | 0.38 | N.E. | Not assessed [24] | |||||||
| Hydroclanthrus clathratus 9 | 0.34 | 40.22 | 1.16 | N.E. | |||||||||
| Cladophora glomerata 6 | 27.70 | N.E. | Not assessed [21] | ||||||||||
| Gelidium crinale 6 | - | - | - | - | - | - | 0.86 | N.E. | Not assessed [30] | ||||
| Sargassum hornschuchii 6 | - | - | - | - | - | - | traces | N.E. | |||||
| Ulva linza 6 | - | - | - | - | - | - | 7.58 | N.E. | |||||
| Sarconema filiforme 3 | 3.67 | 15.67 | N.E. | Not assessed [12] | |||||||||
| Sarconema filiforme 2 | 41.62 | N.E. | |||||||||||
| Laurencia obtusa 3 | 16.12 | N.E. | |||||||||||
| Laurencia obtusa 2 | 54.25 | N.E. | |||||||||||
| Presence (P) or absence (A) of PAEs in algal surface (S), inner part (I), rhizoids (R) or whole (W) | |||||||||||||
| DEP | DnBP | DIBP | BIBP | DEHP | DMBP | MEHP | DnOP | DnDP | DINP | DIOP | |||
| Ulva lactuca 5 | A (W) | P (W) | P (W) | P (W) | N.E. | Bioaccumulation | |||||||
| Enteromorpha linza 5 | A (S) P (I) | P (W) | P (W) | P (W) | N.E. | [35] | |||||||
| Cystoseira barbata 5 | P (S) A (I) | P (W) | P (W) | P (W) | N.E. | ||||||||
| Pterocladia capillacea 5 | P (S) A (I) | P (W) | P (W) | P (W) | N.E. | ||||||||
| Ceramium rubrum 5 | A (W) | P (W) | P (W) | P (W) | N.E. | ||||||||
| Ulva rigida 5 | A (W) | A(W) | P (W) | P (W) | N.E. | Bioaccumulation [36] | |||||||
| Enteromorpha muscoides 5 | P (W) | P (W) | A (W) | A (W) | N.E. | ||||||||
| Enteromorpha linza 5 | A (W) | A(W) | P (W) | P (W) | N.E. | ||||||||
| Gelidium pulchellum 5 | A (W) | A(W) | P (W) | A (W) | N.E. | Bioaccumulation [37] *** | |||||||
| Polysiphonia elongata 5 | A (W) | A(W) | A (W) | P (W) | N.E. | ||||||||
| Coralligena elongata 5 | A (W) | A(W) | A (W) | A (W) | N.E. | ||||||||
| Codium fragile 5 | A (W) | A (W) | A (W) | A (W) | N.E. | ||||||||
| Peyssonnelia squamaria 5 | A (W) | A (W) | P (W) | A (W) | N.E. | ||||||||
| Rhodymenia corrallina 5 | A (W) | A (W) | A (W) | P (W) | N.E. | ||||||||
| Phyllophora nervosa 5 | A (W) | P (W) | P (W) | A (W) | N.E. | ||||||||
| Colpomenia peregrina 5 | A (W) | A (W) | P (W) | P (W) | N.E. | ||||||||
| Cystoseira barbata 5 | A (W) | P (W) | P (W) | A (W) | N.E. | ||||||||
| Zanardina prototypus 5 | A (W) | P (W) | P (W) | A (W) | N.E. | ||||||||
| Gracilaria verrucosa 5 | A (W) | P (W) | A (W) | A (W) | N.E. | ||||||||
| Gracilaria verrucosa 5 | P (W) | N.E. | |||||||||||
| Polysiphonia elongata 5 | P (W) | N.E. | |||||||||||
| Phyllophora nervosa 5 | P (W) | P (W) | N.E. | ||||||||||
| Rhodymenia corrallina 5 | P (W) | N.E. | |||||||||||
| Ceramium rubrum 5 | P (W) | P (W) | P (W) | N.E. | |||||||||
| Gelidium pulchellum 5 | P (W) | P (W) | P (W) | N.E. | |||||||||
| Cystoseira barbata 5 | P (W) | P (W) | P (W) | P (W) | N.E. | ||||||||
| Zanardina prototypus 5 | P (W) | P (W) | N.E. | ||||||||||
| Colpomenia peregrina 5 | P (W) | P (W) | N.E. | ||||||||||
| Ulva rigida 5 | P (W) | N.E. | |||||||||||
| Ulva lactuca 5 | P (W) | P (W) | N.E. | ||||||||||
| Enteromorpha muscoides 5 | P (W) | N.E. | |||||||||||
| Enteromorpha linza 5 | P (W) | P (W) | P (W) | P (W) | N.E. | ||||||||
| Gracilaria lemaneiformis | P (W) | N.E. | Biosynthesis [38] | ||||||||||
| Cladophora fracta | P (W) | P (W) | N.E. | Biosynthesis [39] | |||||||||
| Chaetomorpha basiretorsa | P (W) | P (W) | N.E. | Biosynthesis [40] | |||||||||
| Ulva tepida 5 | P (W) | N.E. | Uncertain [29] | ||||||||||
| Desmarestia anceps 2 | A (W) | A(W) | P (W) | A (W) | A(W) | N.E. | Not assessed [41] | ||||||
| Desmarestia anceps 3 | A (W) | A(W) | P (W) | A (W) | A(W) | N.E. | |||||||
| Pyropia endiviifolia 1 | A (W) | A(W) | P (W) | A (W) | P (W) | N.E. | |||||||
| Pyropia endiviifolia 2 | A (W) | P (W) | P (W) | A (W) | P (W) | P (W) | N.E. | ||||||
| Pyropia endiviifolia 3 | A (W) | P (W) | P (W) | A (W) | P (W) | P (W) | N.E. | ||||||
| Sargassum wightii 4 | P (W) | N.E. | Biosynthesis [23] | ||||||||||
| Caulerpa racemosa | P (W) | P (W) | P (W) | P (W) | N.E. | Uncertain [26] | |||||||
| Codium tomentosum | - | P (W) | P (W) | P (W) | N.E. | ||||||||
| Sargassum confusum 1,6 | P (W) | P (W) | P (W) | N.E. | Biosynthesis [18] | ||||||||
| Stoechospermum marginatum 10 | P (W) | N.E. | Bioaccumulation [17] | ||||||||||
| Sargassum wightii 2,6 | P (W) | N.E. | Biosynthesis [42] | ||||||||||
| Sargassum muticum | P (W) | P (W) | N.E. | Uncertain [43] | |||||||||
| Laminaria japonica 5,6 | P (W) | P (W) | L.C. | Biosynthesis [44] | |||||||||
| Ulva sp. 5,6 | P (W) | P (W) | L.C. | ||||||||||
| Undaria pinnatifida 5,6 | P (W) | P (W) | L.C. | ||||||||||
| Croisetta sp. | P (W) | N.E. | Uncertain [45] | ||||||||||
| Laminaria japonica 3 | P (R) | N.E. | Not assessed [20] | ||||||||||
| Iridea cordata 2,3 | P (W) | P (W) | P (W) | N.E. | Uncertain [31] | ||||||||
* Mean of three different samples: (1) harbor near the effluent from wastewater treatment plant; (2) intertidal zone 0.5 km off the coast; (3) 2 km off the coast. ** Mean of two different strain samples. *** In the study are presented only phthalates found, not those sought and not found. 1 Hexane extract; 2 chloroform extract; 3 ethanol extract; 4 acetone extract; 5 dichlorometane extract; 6 methanol extract; 7 diethyl ether extract; 8 supercritical fluid extraction with CO2; 9 hydro-distillation; 10 methylene chloride.
Most of the research related to these two processes has been conducted on microalgae. Microalgae show promise as bioremediation options for EAS in treatment plants [14] and important sources of phthalate biosynthesis [3,46].
Most research is carried out on microalgae because they have several advantages over macroalgae and higher plants. These advantages include the possibility of mass cultivation, the need for small amounts of microalgae for experiments, fast growth reproduction and production, minimal nutrient requirements, high resistance, ease of extraction of biological compounds, and simple maintenance management [47]. Microalgae are the primary producers in the aquatic food chain, playing a fundamental role in the processes of bioaccumulation and biomagnification [1,9]. They act as the entry point for contaminants into the food chain, posing a direct threat to higher trophic levels, including wildlife and human health. Phthalates can enter aquatic food chains through microalgal bioaccumulation. At the same time, direct microalgal degradation of pollutants is a critical process that affects the environmental fate of PAEs. In addition to this, organic pollutants are degraded by heterotrophic bacteria that are stimulated by the oxygen produced by microalgae [1].
2.2. Hypothesis of PAEs Origin
In general, studies in the literature with a prudential approach do not assess or clarify the origin of phthalates in algae (see Table 2). This is likely because the main objective of the literature articles is not to demonstrate the dual purpose associated with their presence in algal organisms. This, coupled with the ubiquity of phthalates, inevitably leads to analytical difficulties in identifying sources of origin [4]. In fact, a cross-contamination process can cause false positives (commonly DEHP and DnBP) if the necessary measurements are not taken. Thus, contamination of samples and associated blanks can occur in different sample preparation, extraction, and analysis processes. Since the hypothesis of the origin of PAEs in algae could be controversial, the correct application of the most appropriate analytical technique as a function of the context (e.g., sampling site, homogenized sample, interference, etc.) is crucial to develop a solid assessment. Therefore, particular attention must be paid to all laboratory tools or instrumental components that are used and that may contain constitutively or enable contamination. In fact, these can be contaminated by phthalates through both contact with materials that can transfer them, and be the object of deposition of vapors and particulates containing phthalates present in the working environment. In this context, all instruments used should be suitable (e.g., glass and ceramics) and phthalate-free and/or should be properly cleaned with different rinsing cycles using solvents and high temperatures. From an analytical point of view, appropriate testing assurance and quality protocols should be followed in order to correctly identify and quantify phthalates [4]. This problem of misidentification could mainly affect chemical characterization studies where appropriate measures may not be taken during analysis (e.g., use of plastic or inadequately decontaminated material for sample storage or handling).
In addition, the type of extraction solvent used is fundamental in the determination of a phthalate which may or may not be identified (see different extract fraction in Table 2). Most of the works in Table 2 use dichloromethane as an extraction solvent as it appears to have a greater affinity with the phthalates investigated.
In this context, there may be many more phthalates than those identified, particularly in chemical characterization analyses where PAEs may not be identified by type of solvent or analytical method, or in quantitative analyses where the focus is on the identification and quantification of a few compounds. All these factors, including the analysis of only a portion of the organism rather than the entire organism, contribute to an underestimation.
2.2.1. Biosynthesis
Although some macroalgae, such as Ulva sp., Sargassum sp., Gracialaria sp., or Pterocladia sp., have the remarkable capacity to adsorb lipophilic pollutants [13,30] and endocrine active substances [15], including phthalates [8,48], and can potentially be used for bioremediation approaches, studies have shown that micro- and macroalgae produce phthalates via biosynthetic pathways [33,34,44] probably through the shikimic acid pathway [3,49].
Using the isotopic labeling approach, it has been observed that di(n-butyl) phthalate and probably di(2-ethylhexyl) phthalate can be biosynthesized by various macroalgae algae such as Undaria pinnatifida, Laminaria japonica, and Ulva sp. [44].
The presence of DnOP and DEHP has been reported in antifouling active lipid fractions isolated from macroalgae Sargassum muticum [43]. DOP has also been isolated from other brown algae Sargassum wightii [42], Ishige okamurae [50], and Sargassum confusum showing inhibition of spore attachments [18]. The latter work suggests that this antibacterial compound was not derived from the environment.
Similarly, phthalate ester derivates that have shown extensive antimicrobial activity have been isolated from the green macroalgae Ulva fasciata [51].
In another work, it was observed that the content of DEHP and DBP in the filaments of Bangia atropurpurea cultivated in the natural medium were similar to those cultivated in the artificial medium [33]. In this last culture experiment, it was reported that the filaments of the red macroalgae B. atropurpurea synthesized de novo phthalic esters, probably to improve the flexibility of algal cells [33]. Similarly, investigation of 16 species of freshwater microalgae and cyanobacteria under stress conditions showed that some of them produced either DBP or mono(2-ethylhexyl) phthalate (MEHP) or both [34]. This observation indicates that organisms could be a significant source of phthalates in aquatic ecosystems.
As can be seen in Table 2, in the paper of Chen [33], DBP concentrations were found to be higher than those of DEHP, while in the work of Babu and Wu [34], MEHP concentrations were usually higher than those of its precursor (DEHP), probably because it was produced by its degradation.
As reported by Chen [33], PAE levels detected in various algal species cultivated in the same environment were significantly different, suggesting that it was due to the intrinsic nature of the algae. However, this difference may be due to the algal biosorption or biosynthesis capabilities of these substances or both and not exclusively from biosynthetic pathways. In this context, as reported by Sastry and Rao [42], the comparison of the results of the extracts from the analysis of different algal species collected from the same sample site allowed the researchers to discriminate whether the purified extracts and relative phthalates originated from impurities absorbed or from constitutive substances.
As we will see in the next sections on the efficiency of bioaccumulation and the biodegradation of phthalates, environmental parameters also play a fundamental role in the biosynthesis of phthalates. This was observed in cyanobacteria Anabaena circinalis, where factors such as light, temperature, and nutrients were found to affect the production of DEHP [52].
2.2.2. Bioaccumulation
Algae’s biosorption capacity is affected by physicochemical factors, in particular pH, temperature, the type and log Kow of PAEs, weight of algal biomass, and PAEs’ concentration [8,48,53].
Once algal biomass has absorbed phthalates, these pollutants can be removed from the organic matrix using green techniques, such as photoinduced degradation [8]. This matrix can be reused for the same bioremediation application [48], as well as for other applications, such as fertilizers in agriculture.
In fact, it has been observed that the desorption of DEHP from algal biosorbents does not significantly reduce the ability to reuse the same matrix as a biosorbent, suggesting the promising use of macroalgae for repeated bioremediation processes [48].
Along with molecular adsorption capacities, macrophytes such as macroalgae (Padina sp. and Sargassum ilicifolium) and aquatic plants (Cymodocea rotundata, Cymodocea serrulata and Thalassia hemprichii) have been observed to accumulate microplastics, which is likely to further increase phthalate concentrations in their tissues [54].
Most studies that explore new analytical techniques for determining PAEs or aim to monitor the environment assume that the presence of PAEs in aquatic biota is due to bioaccumulation phenomena, as observed in the leaves of the autochthonous seagrass Posidonia oceanica [55,56].
High bioaccumulation factors of phthalates are often found in freshwater and marine algae, as observed in a recent trophodynamics study by Liu et al. [57] in China. The enhanced bioaccumulation capacities in algae are probably related to their life habits and frequent contact with sediments as well as their metabolism [57]. In fact, unlike other organisms such as fish, algae have lower metabolic and excretion capacities, resulting in greater bioaccumulation.
A recent study by Berneira et al. [58] evaluated the volatile composition and fatty acid profile of seven Antarctic macroalgae and recorded the presence of DBP, DEP, and DMP. The study suggests that these substances are not associated with endogenous processes in macroalgae but rather result from environmental pollution. This finding is consistent with previous research on the green marine macroalgae Capsosiphon fulvescens, collected in South Korea [59].
It is widely acknowledged that phthalate esters significantly contribute to the contamination of aquatic organisms due to their lipophilic properties, which lead to bioaccumulation.
Some authors have questioned their origin [43], while others have concluded that phthalates are certainly taken from the aquatic environment, although the mechanisms related to their accumulation or genesis are not yet fully understood [17]. Phthalates found in the environment may originate from industrial waste, decomposition products of certain algae, degradation of humic substances by microorganisms, or dispersants (chemical mixtures used to disperse oil spills) [17].
Furthermore, sources of phthalate contamination may arise from external sources or laboratory materials used in the processes of extraction, isolation of natural products, and analysis. It can be challenging to identify the sources of PAE contamination.
Several studies [8,35,57] suggest that the presence of phthalates in algae is due to bioaccumulation phenomena, which reflect its environmental contamination. For example, Gezgin et al. [35] found parallels between phthalates (DEP, DnBP, DIBP, and DEHP) in macroalgae (Ulva lactuca, Enteromorpha linza, Cystoseria barbata, Pterocladia capillaceae, and Ceramium rubrum) and in seawater.
Based on the results presented by Gezgin et al. [35], it can be inferred that the presence of phthalates in both the internal and external parts of a system could be attributed to both biosynthesis and accumulation. However, if phthalates are only found on the surface, it may suggest that the predominant process is exogenous (environmental contamination) rather than endogenous (biosynthesis). In contrast, if phthalates are found only within the seaweed, it could be assumed that their presence is connected to biosynthetic pathways.
2.2.3. Critical Analysis of Undefined PAEs’ Origin
The chemical analysis of phthalates in the biological matrix alone raises questions about the presence of these substances in the environment and thus about the source of contamination. In the absence of data on environmental contamination, a dual source of contamination cannot be excluded. In this type of study, analysis of not only the organism but also the water and other environmental matrices is strongly recommended. Several studies do not have these objectives and therefore do not present or discuss the origin of phthalates but, based on the conditions of the study, it is possible to assume that the contamination is mainly related to environmental pollution or mainly due to endogenous factors.
As previously mentioned, some studies have confirmed that phthalates are produced by algae. These works aim to investigate the origin of phthalates [33,34,44] or consider phthalates as chemical constituents of algae, as observed in chemical characterization of algae [27,38,40] and in studies on the biological activity of algae extracts [18,39,42]. Some studies also suggest that phthalates are naturally present [23] or mention other studies that have reported that they are naturally produced [24]. Other papers suggest that the presence of phthalates in algae is due to environmental pollution, indicating that algae can be used as indicators of pollution in aquatic environments [8,17,18,36,37].
However, most of the work (see not assessed and uncertain in Table 2) does not define the origin of phthalates. Some researchers consider phthalates to be phytochemical constituents and/or bioactive components of algae [12,20,21,30,41], which could be considered biosynthetic.
Similarly, some works do not provide certainty on the occurrence of phthalates because they report works that consider phthalates coming both from external contamination and from natural biosynthesis origin [43], suggesting that the presence of phthalates may arise from one of the two sources [45], or more generally that the presence of these substances in algae could be related not only to exposure or environmental contamination during the separation process, but also their biosynthesis from algae [31].
2.3. Algae as the Future of PAEs’ Bioremediation
Although phthalates are produced by algae, exposure to high concentrations of phthalates has been shown to induce negative effects in both macro- [8] and microalgae [60,61,62,63,64]. At the same time, it has been observed that several microalgal species can degrade phthalates. Marine microalgae species (Cylindrotheca Closterium, Dunaliella salina, and Chaetoceros muelleri) have degraded DEP and DBP through intra- and extracellular esterase enzymes [1]. Production and degradation suggest that environmental conditions determine the activation or inhibition of specific pathways.
However, while algae can be negatively impacted by these contaminants, they also have the ability to bioaccumulate and biodegrade phthalic esters, as demonstrated in the case of the marine dinoflagellate Karenia brevis and the study of degradation pathways (esterification, demethylation, or transesterification) of DEP and dipropyl phthalate (DPrP) [65].
Several factors may determine an increase in bioaccumulation and/or biodegradation. For example, low temperatures have been observed to increase the half-life of PAE bioaccumulation and inhibition of their biodegradation by algae [66]. In this context, it has been observed that the rate of biodegradation is influenced by the presence of inorganic carbon, which leads to an increase in microalgal growth in Closterium lunula, as well as an increase in the biodegradation of DEP [67].
It is important to note that nitrogen and phosphorus, which are the main nutrients, play a crucial role in both bioconcentration and biodegradation. Studies have shown that adding these nutrients to lake water has an impact on the rate of biodegradation and bioconcentration of DBP and DEHP in the microalgae Chlorella vulgaris [68].
This study found that the addition of nitrogen had a more pronounced effect than the addition of phosphorous on degradation. However, the addition of both nutrients had a greater influence on degradation than the addition of a single nutrient [68].
The bioconcentration factor (BCF) was found to be strongly correlated with the amount of dissolved organic carbon (DOC). Specifically, the BCF decreased with an increase in algal exudate, as measured by DOC [68].
The same observations were made in a similar experiment conducted on Microcystis aeruginosa that showed high rates of bioconcentration and biodegradation of DBP and DEHP [66]. These results suggest that DOC plays an important role in the bioconcentration of DBP and DEHP [66,68].
It is important to highlight that algae biodegrade differently according to the concentration level of exposure, the type of phthalate, the physical and chemical parameters, and the species being considered.
In a study involving three marine microalgae (Chaetoceros muelleri, Cylindrotheca closterium, Dunaliella salina) exposed to DEP and DBP, it was observed that these three algae have degraded PAEs more rapidly at an environmentally relevant concentration (0.1 mg L−1).
In particular, the degradation was more efficient when PAEs were taken individually, while the mixture of the two phthalates showed inhibitory effects on the process [69]. Overall, all species demonstrated strong adaptability. However, C. closterium exhibited greater efficiency in degrading phthalates compared to the other two species [69].
To assess the effectiveness of bioremediation techniques using algae, it is important to deepen our understanding of the metabolic dynamics involved in the degradation and production of these substances. Therefore, analytical techniques and biomolecular studies should be adapted for both laboratory experiments and environmental monitoring.
2.4. Photodegradation of Phthalates
Sunlight is essential for the survival of photosynthetic organisms and at the same time can affect the stability of photosensitive organic compounds such as phthalates [8,9].
While biodegradation is the primary process that affects the fate of phthalates in water, other abiotic processes, such as photodegradation, can also impact their stability depending on environmental conditions [53]. Photoinduced degradation is primarily caused by the UV component of solar radiation, specifically UV-B and UV-A. Phthalates, in particular, exhibit maximum photodegradation efficiency at a wavelength of 254 nm, which involves reactions such as decarboxylation, hydroxylation, dealkylation, and splitting of C-O, C-C, and O-alkyl bonds [8]. However, this wavelength cannot penetrate the Earth’s surface [70].
These reactions seem to play a crucial role in the atmospheric fate of phthalate esters. The most significant photodegradation process for organic chemical pollutants in the atmosphere is their reaction with hydroxyl radicals. Under these environmental conditions, a half-life for DEHP of about one day was demonstrated [53].
For instance, when dry algal matrices (Ulva lactuca) were subjected to UV irradiation in laboratory conditions, a high removal efficiency of PAEs was observed. Specifically, degradation rates of DMP, DEP, DBP, BBP, DEHP, and DnOP ranged from 60% to 100% after 8 h of exposure to lamps emitting UV radiation (with a main wavelength of 254 nm) [8].
In an aqueous medium, photolysis occurs through the absorption of UV light from sunlight. Shorter wavelengths are attenuated by passage through the atmosphere and water column, while longer wavelengths lack sufficient energy to break covalent bonds. Photolysis can occur through direct or indirect mechanisms. Direct photolysis involves the absorption of UV radiation by the chemical, while indirect photolysis occurs when natural substances such as water absorb UV radiation, leading to the formation of activated species like singlet oxygen or hydroxy radicals that react with phthalate esters. It is important to note that the mechanism of photolysis can be mediated by either of these pathways [53].
According to Gledhill et al. [71], a 1 mg/L solution of BBP exposed to sunlight for 28 days underwent less than 5% degradation. The study concluded that BBP has an aqueous photolysis half-life of more than 100 days.
Photooxidation of phthalate esters in surface waters does not appear to be a significant transformation process. The half-life for aqueous photooxidation ranges from 0.12 to 1.5 years for DEHP, and from 2.4 to 12 years for DEP and DnBP [53].
Variation in pollutant degradation kinetics can be caused by different light sources, initial concentration, and the type of experimental medium, as well as its physical and chemical parameters [8,53,72,73].
Due to the benzene carboxyl group, phthalates have reduced sensitivity to wavelengths above 300 nm, resulting in decreased photosensitivity to natural sunlight. This effect is typically more pronounced at longer wavelengths, with λ radiation greater than 370 nm leading to increased photodegradation time [73]. In a photodegradation experiment, DMP and PDO were tested at a concentration of 0.20 mg/L using a solar simulator that emitted radiation similar to solar radiation in the UV region (λ: 280 nm to 400 nm) [73]. In this experiment, conducted in an aqueous medium, a half-life of 19.2 days for DMP and 3.99 days for PDO was observed [73].
In a photodegradation experiment conducted in natural sunlight, DBP, BBP, DEHP, and DINP showed longer degradation times at initial concentrations above 120 mg/L [72]. Specifically, DBP had a half-life of 360 days, DEHP had a half-life of 1600 days, and DINP had a half-life of 140 days. However, DBP and BBP had longer half-lives in darkness (1300 days and 1500 days, respectively) [72]. In contrast, a 254 nm photodegradation experiment in an aqueous solvent showed significantly lower half-lives for DMP (half-life = 26.3 h), DBP (half-life = 6.9 h), and DOP (half-life = 0.72 h) at initial concentrations of about 0.6 mg/L [74].
In this context, it is necessary to assess whether photodegradation produces toxic intermediate products or proceeds to complete phthalate mineralization [73].
In the natural aquatic environment, the solar photodegradation efficiency of phthalates would be lower than in the experimental environment due to factors such as water turbidity, depth, and other environmental characteristics, as well as chemical and physical characteristics that affect the penetration of radiation in water [53,70]. PAEs, being lipophilic organic chemicals, are easily adsorbed by organic matter in the environment. Understanding the photodegradation mechanism of PAEs in organic matter and the factors that affect degradation rates is complicated due to the organic phase’s complex nature [73].
It can be concluded that solar light contributes to the degradation of phthalates both directly, through photodegradation to a lesser extent, and indirectly to a greater extent, by stimulating the growth of photosynthetic organisms that biodegrade these organic compounds with high efficiency.
3. Future Directions
Ensuring molecular identity and concentration levels is crucial. This can be achieved through techniques such as nuclear magnetic resonance and chromatography associated with mass spectrometry.
Organism-derived phthalates differ chemically from synthetic ones (of petroleum origin) in terms of the abundance of 14C and bond structure [16]. However, further verification is required to confirm the origin of PAEs in algae.
In treatment plants, pollutants undergo various processes such as sedimentation, physical filtration, adsorption, biodegradation, volatilization, and plant uptake. Therefore, it is necessary to use mass balance models that consider incoming and outgoing loads and the evolution of the pollutant mass [75].
In controlled experiments, it is crucial to analyze all components of the system at the beginning and end to evaluate the evolution of pollutants. Therefore, it is essential to analyze all system components. Missing quantities may have been metabolized by the algae, while quantities higher than the initial ones would indicate production.
A comprehensive analysis of phthalates produced solely industrially or by organisms, such as dialkyl phthalates and bis(2-methylheptyl) phthalate, would yield valuable information on their origin. However, these phthalates are less prevalent and have been less studied [46]. Accurately defining algal species and their ability to biosynthesize phthalates requires phylogenetic and chemotaxonomic studies. These studies can determine whether the phthalates are unique to the species or produced by other sources. However, if no specific approaches, such as isotopic analysis, are used to identify the origin of a particular phthalate, evaluations should be conducted to form a preliminary hypothesis of its origin.
In a classic biomonitoring study, three scenarios may occur during the analysis of different individuals belonging to the same species:
- (1)
- Phthalate is present in all individuals;
- (2)
- Phthalate is not present in any individual;
- (3)
- Phthalate is present with variable frequency between individuals.
In the first case, the contamination may result from environmental bioaccumulation and/or biosynthetic processes.
In the second case, it could be thought that phthalate is not produced by biosynthetic means and that there is no significant environmental contamination such as to determine bioaccumulation. However, although rarely, it is possible that even if the environment is contaminated, no bioaccumulation occurs due to specific environmental conditions (physical chemical parameters, lower affinity of the substance for the algal matrix than other biological matrices, altered physiological state of the organism, etc.). In this scenario, it is also possible that there is no instrumentally detectable contamination or that species produced quantities of phthalates not detectable instrumentally.
In the third scenario, if some individuals are contaminated and others are not, it could be hypothesized that the predominant phenomenon of contamination is connected to environmental bioaccumulation, unless the species produces phthalates in undetectable quantities or is differently biosynthesized due to local environmental variables.
It is important to emphasize that bioaccumulated phthalates could be of both anthropogenic and natural origin.
The current challenge for research is to conduct further investigations to differentiate and quantify the two sources of PAEs, as well as the environmental conditions that lead to increased production and adsorption. Additionally, using marked phthalates to distinguish between analytes added to the system and those produced or removed would be advantageous. Constant monitoring of experimental parameters is essential in determining changes in phthalate concentration in algal matrices.
Although many countries have severely limited the production and use of phthalates, constant and increasing anthropic disturbance, including pollution from other substances, may still lead to an increase in phthalate concentrations. This increase could result in the production of metabolites by stressed organisms. Production and degradation suggest that environmental conditions determine the activation or inhibition of specific pathways. To better understand the origin of phthalates, it is necessary to conduct genetic and biochemical analyses on the biosynthesis pathways. This will help identify the specific genes and metabolic pathways involved in the process. Currently, due to the lack of research in this area, it is only possible to make hypotheses.
Phthalates could be synthesized in algae through the shikimic acid pathway, as observed in filamentous fungal strains for the production of dibutyl phthalate [76]. This compound is assembled from protocatechuic acid to phthalic acid, and finally with butyl alcohol through esterification [76].
Fungal spore density and incubation temperature [76] mainly influence this production, but it can also be regulated by other physical and nutritional factors [16].
The biosynthesis of phthalates can be influenced by various factors. Physiological differences and specific metabolic needs vary between algal and fungal organisms, as well as within the same realm.
The production of secondary metabolites is determined by the organism’s genetic makeup and is regulated by gene expression, which is influenced by limiting conditions. The biosynthetic pathways for these metabolites are complex and affected by both biotic and abiotic factors. Therefore, studies on different species are necessary to confirm the specific biomolecular pathways of phthalate production. Further investigation is necessary to fully understand these processes.
In this way, it will be informative to observe the direction in which the production/degradation equilibrium shifts. Evaluating the potential applications of production or inhibition of production and bioremediation to enhance environmental conditions would be worthwhile.
Additional research is necessary to determine which species produce phthalates and which do not, and whether the latter can be utilized as biosorbents for environmental remediation.
In conclusion, all these papers show that the natural production of PAEs as chemically active metabolites is a common phenomenon that contributes to the contamination of the aquatic environment. The work mentioned above makes it clear that the natural production of phthalates would further exacerbate the already widespread environmental contamination caused by the dispersion of materials containing these additives.
The contribution of phthalates to environmental contamination from algal biosynthesis appears insignificant compared to human activities. However, some authors attribute the greatest contribution of present phthalates in the environment to the plant world. It has been estimated that the production of phthalates of natural origin, excluding the unquantifiable contribution of bacteria, fungi, and algae, is about 36 times greater than the annual world production of 4.9 million tons (according to 2019 data) [49]. Therefore, considering the current restrictions on the use of phthalates and the estimates mentioned above, it can be concluded that the percentage of PAEs from natural sources is higher than that from industrial production sources, which are therefore insignificant in comparison.
In this context, it is undeniable that the risks associated with the consumption of algae or medicinal plants due to the presence of phthalates should be thoroughly assessed, despite their potential as a source of bioactive molecules for various applications.
Author Contributions
Conceptualization and methodology, D.S.; investigation, visualization, and data curation, D.S. and A.V.; writing—original draft preparation, D.S.; writing—review and editing, D.S., A.M., A.V. and A.P.; supervision, A.M., A.V. and A.P. All authors have read and agreed to the published version of the manuscript.
Funding
Project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4—Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union—NextGenerationEU. Award Number: Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP B73C22000790001, Project title “National Biodiversity Future Center—NBFC”.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gao, J.; Chi, J. Biodegradation of Phthalate Acid Esters by Different Marine Microalgal Species. Mar. Pollut. Bull. 2015, 99, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Das, M.T.; Kumar, S.S.; Ghosh, P.; Shah, G.; Malyan, S.K.; Bajar, S.; Thakur, I.S.; Singh, L. Remediation Strategies for Mitigation of Phthalate Pollution: Challenges and Future Perspectives. J. Hazard. Mater. 2021, 409, 124496. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhu, X.; Zhou, S.; Cheng, Z.; Shi, K.; Zhang, C.; Shao, H. Phthalic Acid Esters: Natural Sources and Biological Activities. Toxins 2021, 13, 495. [Google Scholar] [CrossRef]
- Savoca, D.; Barreca, S.; Lo Coco, R.; Punginelli, D.; Orecchio, S.; Maccotta, A. Environmental Aspect Concerning Phthalates Contamination: Analytical Approaches and Assessment of Biomonitoring in the Aquatic Environment. Environments 2023, 10, 99. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Fasano, M.; Palandri, L.; Righi, E. A review of European and international phthalates regulation: Focus on daily use products. Eur. J. Public Health 2022, 32, ckac131-226. [Google Scholar] [CrossRef]
- Zhao, E.; Xiong, X.; Hu, H.; Li, X.; Wu, C. Phthalates in Plastic Stationery in China and Their Exposure Risks to School-Aged Children. Chemosphere 2023, 339, 139763. [Google Scholar] [CrossRef] [PubMed]
- Savoca, D.; Lo Coco, R.; Melfi, R.; Pace, A. Uptake and Photoinduced Degradation of Phthalic Acid Esters (PAEs) in Ulva Lactuca Highlight Its Potential Application in Environmental Bioremediation. Environ. Sci. Pollut. Res. 2022, 29, 90887–90897. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.L. Algae: The World’s Most Important “Plants”—An Introduction. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 5–12. [Google Scholar] [CrossRef]
- Pereira, L. Macroalgae. Encyclopedia 2021, 1, 177–188. [Google Scholar] [CrossRef]
- Correa, I.; Drews, P.; Botelho, S.; De Souza, M.S.; Tavano, V.M. Deep Learning for Microalgae Classification. In Proceedings of the 2017 16th IEEE International Conference on Machine Learning and Applications (ICMLA), Cancun, Mexico, 18–21 December 2017; IEEE: New York, NY, USA; pp. 20–25. [Google Scholar]
- Teleb, W.K.; Tantawy, M.A.; Osman, N.A.H.K.; Abdel-Rahman, M.A.; Hussein, A.A. Structural and Cytotoxic Characterization of the Marine Red Algae Sarconema Filiforme and Laurencia Obtusa. Egypt. J. Aquat. Biol. Fish 2022, 26, 549–573. [Google Scholar] [CrossRef]
- Cheney, D.; Rajic, L.; Sly, E.; Meric, D.; Sheahan, T. Uptake of PCBs Contained in Marine Sediments by the Green Macroalga Ulva Rigida. Mar. Pollut. Bull. 2014, 88, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Roccuzzo, S.; Beckerman, A.P.; Trögl, J. New Perspectives on the Bioremediation of Endocrine Disrupting Compounds from Wastewater Using Algae-, Bacteria- and Fungi-Based Technologies. Int. J. Environ. Sci. Technol. 2021, 18, 89–106. [Google Scholar] [CrossRef]
- Astrahan, P.; Korzen, L.; Khanin, M.; Sharoni, Y.; Israel, Á. Seaweeds Fast EDC Bioremediation: Supporting Evidence of EE2 and BPA Degradation by the Red Seaweed Gracilaria Sp., and a Proposed Model for the Remedy of Marine-Borne Phenol Pollutants. Environ. Pollut. 2021, 278, 116853. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.N. Bioactive Natural Derivatives of Phthalate Ester. Crit. Rev. Biotechnol. 2020, 40, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Wahidulla, S.; De Souza, L. Phthalate Esters from the Brown Alga Stoechospermum Marginatum (C. Agardh) Kuetzing. Bot. Mar. 1995, 38, 333–334. [Google Scholar] [CrossRef]
- Ganti, V.S.; Kim, K.H.; Bhattarai, H.D.; Shin, H.W. Isolation and Characterisation of Some Antifouling Agents from the Brown Alga Sargassum Confusum. J. Asian Nat. Prod. Res. 2006, 8, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraj, R.; Altaff, K.; Kumaran, N.S. Volatile Bioactive Compounds from Marine Macro-Algae and Their Pharmacological Properties. Indian J. Geo-Marine Sci. 2022, 51, 832–842. [Google Scholar] [CrossRef]
- Bu, T.; Liu, M.; Zheng, L.; Guo, Y.; Lin, X. A-glucosidase Inhibition and the in Vivo Hypoglycemic Effect of Butyl-isobutyl-phthalate Derived from the Laminaria Japonica Rhizoid. Phytother. Res. 2010, 24, 1588–1591. [Google Scholar] [CrossRef] [PubMed]
- Usha, N.S.; Sabari Anand, J.V. Mangaiyarkarasi Preliminary Phytochemical Screening and GC-MS Analysis of Cladophora Glomerata: Green Marine Algae. Int. J. Basic Clin. Pharmacol. 2019, 8, 732. [Google Scholar] [CrossRef]
- El Shouny, W.A.; Gaafar, R.M.; Ismail, G.A.; Elzanaty, M.M. Antibacterial Activity of Some Seaweed Extracts against Multidrug Resistant Urinary Tract Bacteria and Analysis of Their Virulence Genes. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 2569–2586. [Google Scholar] [CrossRef]
- Rosaline, D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Detection of Antibacterial Activity and Its Characterization from the Marine Macro-Algae Sargassum Wightii (Greville Ex J. Agardh 1848). Indian J. Mar. Sci. 2016, 45, 1365–1371. [Google Scholar]
- Awad, N.E.; Motawe, H.M.; Selim, M.A.; Matloub, A.A. Volatile Constituents of the Brown Algae Padina Pavonia (L.) Gaill. and Hydroclathrus Clathratus (C. Agardh) Howe and Their Antimicrobial Activity. Med. Aromat. Plant Sci. Biotechnol. 2009, 3, 12–15. [Google Scholar]
- Awad, N.E.; Selim, M.A.; Saleh, M.M.; Matloub, A.A. Seasonal variation of the lipoidal matters and hypolipidaemic activity of the red alga Corallina officinalis L. Phytother. Res. 2003, 17, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Awad, N.E. Volatile constituents, carbohydrate compositions and topical anti-inflammatory of the two green algae Caulerpa racemosa (O. Dargent) and Codium tomentosum (Stackhouse). Bull. Fac. Pharm. 2002, 40, 233–243. [Google Scholar]
- Firouzi, J.; Gohari, A.R.; Rustaiyan, A.; Larijani, K.; Saeidnia, S. Composition of the Essential Oil of Nizamuddinia Zanardinii, a Brown Alga Collected from Oman Gulf. J. Essent. Oil Bear. Plants 2013, 16, 689–692. [Google Scholar] [CrossRef]
- Noguchi, T.; Ikawa, M.; Uebel, J.J.; Andersen, K.K. Lipid constituents of the red algae Ceramium rubrum. A search for antimicrobial and chemical defense substances. In Marine Algae in Pharmaceutical Science; Hoppe, H.A., Levring, T., Tanaka, Y., Eds.; Walter de Gruyter & Co: New York, NY, USA, 1979; pp. 711–718. [Google Scholar]
- Agusman; Qi, Y.; Wu, Z.; He, J.; Rittschof, D.; Su, P.; Ke, C.; Feng, D. Conspecific Cues That Induce Spore Settlement in the Biofouling and Green Tide-Forming Alga Ulva Tepida Provide a Potential Aggregation Mechanism. Int. Biodeterior. Biodegrad. 2019, 145, 104807. [Google Scholar] [CrossRef]
- Mohy El-Din, S.M.; Alagawany, N.I. Phytochemical Constituents and Anticoagulation Property of Marine Algae Gelidium Crinale, Sargassum Hornschuchii and Ulva Linza. Thalassas 2019, 35, 381–397. [Google Scholar] [CrossRef]
- Rangel, K.C.; Debonsi, H.M.; Clementino, L.C.; Graminha, M.A.S.; Vilela, L.Z.; Colepicolo, P.; Gaspar, L.R. Antileishmanial Activity of the Antarctic Red Algae Iridaea Cordata (Gigartinaceae; Rhodophyta). J. Appl. Phycol. 2019, 31, 825–834. [Google Scholar] [CrossRef]
- Gu¨ven, K.C.; Reisch, J.; Kizil, Z.; Gu¨vener, B.; Cevher, E. Dimethyl Terephthalate Pollution in Red Algae. Phytochemistry 1990, 29, 3115. [Google Scholar] [CrossRef]
- Chen, C.Y. Biosynthesis of Di-(2-Ethylhexyl) Phthalate (DEHP) and Di-n-Butyl Phthalate (DBP) from Red Alga—Bangia Atropurpurea. Water Res. 2004, 38, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Babu, B.; Wu, J.-T. Production of Phthalate Esters by Nuisance Freshwater Algae and Cyanobacteria. Sci. Total Environ. 2010, 408, 4969–4975. [Google Scholar] [CrossRef] [PubMed]
- Gezgin, T.; Güven, K.C.; Akçin, G. Phthalate esters in marine algae. Turk. J. Mar. Sci. 2001, 7, 119–130. [Google Scholar]
- Erakin, S.; Güven, K.C. The volatile petroleum hydrocarbons in marine algae around Turkish Coasts. Acta Pharm. Sci. 2008, 50, 167–182. [Google Scholar]
- Erakin, S.; Binark, N.; Güven, K.C.; Coban, B.; Erduğan, H. Phthalates pollution in algae of Turkish coast. J Black Sea/Medit. 2014, 20, 122–126. [Google Scholar]
- Lu, H.; Xie, H.; Yang, Y.; Wei, X. Chemical Constituents from the Macroalga Gracilaria lemaneiformis. J. Trop. Subtrop. Bot. 2011, 19, 166–170. [Google Scholar]
- Dong, S.J.; Bi, X.D.; Wang, N.; Song, L.; Dai, W.; Zhang, S.L. Algicidal activities of Cladophora fracta on red tide-forming microalgae Heterosigma akashiwo and Gymnodinium breve. Allelopath. J. 2016, 37, 231–240. [Google Scholar]
- Shi, D.; Han, L.; Sun, J.; Wang, Y.; Yang, Y.; Shi, J.; Fan, X. Chemical constituents from marine alga Chaetomorpha basiretorsa. China J. Chin. Mater. Med. 2005, 30, 347–350. [Google Scholar]
- De Souza, P.O.; Silva, F.A.; Frozza, C.O.D.S.; Frassini, R.; Roesch-Ely, M.; Dos Santos, M.A.Z.; Freitag, R.A.; Colepicolo, P.; Pereira, C.M.P.; Braganhol, E. Bioprospecting of New Antarctic Seaweed Selective Antitumor Molecules: Chemical Characterization and in Vitro Analysis. Phytomed. Plus 2022, 2, 100246. [Google Scholar] [CrossRef]
- Sastry, V.M.V.S.; Rao, G.R.K. Dioctyl Phthalate, and Antibacterial Compound from the Marine Brown Alga? Sargassum Wightii. J. Appl. Phycol. 1995, 7, 185–186. [Google Scholar] [CrossRef]
- Bazes, A.; Silkina, A.; Douzenel, P.; Faÿ, F.; Kervarec, N.; Morin, D.; Berge, J.-P.; Bourgougnon, N. Investigation of the Antifouling Constituents from the Brown Alga Sargassum Muticum (Yendo) Fensholt. J. Appl. Phycol. 2009, 21, 395–403. [Google Scholar] [CrossRef]
- Namikoshi, M.; Fujiwara, T.; Nishikawa, T.; Ukai, K. Natural Abundance 14C Content of Dibutyl Phthalate (DBP) from Three Marine Algae. Mar. Drugs 2006, 4, 290–297. [Google Scholar] [CrossRef]
- Kasanah, N.; Seto, D.S.; Ulfah, M.; Peng, Z. Triyanto First Report on Chemistry of a Red Seaweed Croisettea Sp. from the Coastal Area of Yogyakarta, Indonesia. Mar. Biol. Res. 2022, 18, 215–221. [Google Scholar] [CrossRef]
- Thiemann, T. Isolation of Phthalates and Terephthalates from Plant Material—Natural Products or Contaminants? Open Chem. J. 2021, 8, 1–36. [Google Scholar] [CrossRef]
- Masojídek, J.; Ranglová, K.; Lakatos, G.E.; Silva Benavides, A.M.; Torzillo, G. Variables Governing Photosynthesis and Growth in Microalgae Mass Cultures. Processes 2021, 9, 820. [Google Scholar] [CrossRef]
- Chan, H.W.; Lau, T.C.; Ang, P.O.; Wu, M.; Wong, P.K. Biosorption of Di(2-Ethylhexyl)Phthalate by Seaweed Biomass. J. Appl. Phycol. 2004, 16, 263–274. [Google Scholar] [CrossRef]
- Semenov, A.A.; Enikeev, A.G.; Babenko, T.A.; Shafikova, T.N.; Gorshkov, A.G. Phthalates—A Strange Delusion of Ecologists. Theor. Appl. Ecol. 2021, 1, 16–21. [Google Scholar] [CrossRef]
- Cho, J.-Y.; Choi, J.-S.; Kang, S.-E.; Kim, J.-K.; Shin, H.-W.; Hong, Y.-K. Isolation of Antifouling Active Pyroglutamic Acid, Triethyl Citrate and Di-n-Octylphthalate from the Brown Seaweed Ishige Okamurae. J. Appl. Phycol. 2005, 17, 431–435. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Aboshady, A.M.; Elshobary, M.E. Production and Characterization of Antimicrobial Active Substance from Some Macroalgae Collected from Abu-Qir Bay Alexandria, Egypt. Afr. J. Biotechnol. 2013, 12, 6847–6858. [Google Scholar]
- Sarkar, A.; Bagavananthem Andavan, G.S. Computational Optimization of in Vitro Parameters for Di-(2-ethylhexyl) Phthalate Production from Anabaena Circinalis. J. Cell. Biochem. 2019, 120, 790–798. [Google Scholar] [CrossRef]
- Staples, C.A.; Peterson, D.R.; Parkerton, T.F.; Adams, W.J. The Environmental Fate of Phthalate Esters: A Literature Review. Chemosphere 1997, 35, 667–749. [Google Scholar] [CrossRef]
- Seng, N.; Lai, S.; Fong, J.; Saleh, M.F.; Cheng, C.; Cheok, Z.Y.; Todd, P.A. Early evidence of microplastics on seagrass and macroalgae. Mar. Freshw. 2020, 71, 922–928. [Google Scholar] [CrossRef]
- Castro-Jiménez, J.; Ratola, N. An Innovative Approach for the Simultaneous Quantitative Screening of Organic Plastic Additives in Complex Matrices in Marine Coastal Areas. Environ. Sci. Pollut. Res. 2020, 27, 11450–11457. [Google Scholar] [CrossRef] [PubMed]
- Jebara, A.; Albergamo, A.; Rando, R.; Potortì, A.G.; Lo Turco, V.; Mansour, H.B.; Di Bella, G. Phthalates and Non-Phthalate Plasticizers in Tunisian Marine Samples: Occurrence, Spatial Distribution and Seasonal Variation. Mar. Pollut. Bull. 2021, 163, 111967. [Google Scholar] [CrossRef]
- Liu, B.; Lv, L.; Ding, L.; Gao, L.; Li, J.; Ma, X.; Yu, Y. Comparison of Phthalate Esters (PAEs) in Freshwater and Marine Food Webs: Occurrence, Bioaccumulation, and Trophodynamics. J. Hazard. Mater. 2024, 466, 133534. [Google Scholar] [CrossRef] [PubMed]
- Berneira, L.; Da Silva, C.; Poletti, T.; Ritter, M.; Dos Santos, M.; Colepicolo, P.; De Pereira, C.M.P. Evaluation of the Volatile Composition and Fatty Acid Profile of Seven Antarctic Macroalgae. J. Appl. Phycol. 2020, 32, 3319–3329. [Google Scholar] [CrossRef]
- Sun, S.-M.; Chung, G.-H.; Shin, T.-S. Volatile Compounds of the Green Alga, Capsosiphon Fulvescens. J. Appl. Phycol. 2012, 24, 1003–1013. [Google Scholar] [CrossRef]
- Li, F.; Wu, M.; Yao, Y.; Zheng, X.; Zhao, J.; Wang, Z.; Xing, B. Inhibitory Effects and Oxidative Target Site of Dibutyl Phthalate on Karenia Brevis. Chemosphere 2015, 132, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yi, X.; Zhou, H.; Chi, T.; Li, W.; Yang, K. Combined Effect of Polystyrene Microplastics and Dibutyl Phthalate on the Microalgae Chlorella pyrenoidosa. Environ. Pollut. 2020, 257, 113604. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.J.; Zhao, W.Y.; Cheng, S.P. Toxicity of Dibutyl Phthalate to Algae. Bull. Environ. Contam. Toxicol. 2003, 71, 602–608. [Google Scholar] [CrossRef]
- Melin, C.; Egnéus, H. Effects of Di- n -butyl Phthalate on Growth and Photosynthesis in Algae and on Isolated Organelles from Higher Plants. Physiol. Plant. 1983, 59, 461–466. [Google Scholar] [CrossRef]
- Gu, S.; Zheng, H.; Xu, Q.; Sun, C.; Shi, M.; Wang, Z.; Li, F. Comparative Toxicity of the Plasticizer Dibutyl Phthalate to Two Freshwater Algae. Aquat. Toxicol. 2017, 191, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, G.; Zheng, H.; Liu, N.; Shi, M.; Luo, X.; Chen, L.; Li, F.; Hu, S. Fate of Four Phthalate Esters with Presence of Karenia Brevis: Uptake and Biodegradation. Aquat. Toxicol. 2019, 206, 81–90. [Google Scholar] [CrossRef]
- Chi, J.; Liu, H.; Li, B.; Huang, G.-L. Accumulation and Biodegradation of Dibutyl Phthalate in Chlorella vulgaris. Bull. Environ. Contam. Toxicol. 2006, 77, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Pan, G. Increase in Biodegradation of Dimethyl Phthalate by Closterium Lunula Using Inorganic Carbon. Chemosphere 2004, 55, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Li, B.; Wang, Q.Y.; Liu, H. Influence of Nutrient Level on Biodegradation and Bioconcentration of Phthalate Acid Esters in Chlorella vulgaris. J. Environ. Sci. Health Part A 2007, 42, 179–183. [Google Scholar] [CrossRef]
- Chi, J.; Li, Y.; Gao, J. Interaction between Three Marine Microalgae and Two Phthalate Acid Esters. Ecotoxicol. Environ. Saf. 2019, 170, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, C.J. Ultraviolet radiation and phytoplankton photosynthesis 1. Limnol. Oceanogr. 1979, 24, 1117–1120. [Google Scholar] [CrossRef]
- Gledhill, W.E.; Kaley, R.G.; Adams, W.J.; Hicks, O.; Michael, P.R.; Saeger, V.W.; LeBlanc, G.A. An environmental safety assessment of butyl benzyl phthalate. Environ. Sci. Technol. 1980, 14, 301–305. [Google Scholar] [CrossRef]
- Lertsirisopon, R.; Soda, S.; Sei, K.; Ike, M. Abiotic degradation of four phthalic acid esters in aqueous phase under natural sunlight irradiation. J. Environ. Sci. 2009, 21, 285–290. [Google Scholar] [CrossRef]
- Feng, J.R.; Deng, Q.X.; Ni, H.G. Photodegradation of phthalic acid esters under simulated sunlight: Mechanism, kinetics, and toxicity change. Chemosphere 2022, 299, 134475. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Li, X.; Feng, L. Behavior of stable carbon isotope of phthalate acid esters during photolysis under ultraviolet irradiation. Chemosphere 2013, 92, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, K.; Branchu, P.; Boudahmane, L.; Caupos, E.; Demare, D.; Deshayes, S.; Dubois, P.; Kajeiou, M.; Meffray, L.; Partibane, C.; et al. Mass Balance of Micropollutants over the First Year of Operation in a Stormwater Biofilter: A Field Approach Integrating Water and Soil. 2019. Available online: https://enpc.hal.science/hal-02190391/document (accessed on 20 February 2024).
- Tian, C.; Ni, J.; Chang, F.; Liu, S.; Xu, N.; Sun, W.; Xie, Y.; Guo, Y.; Ma, Y.; Yang, Z.; et al. Bio-Source of di-n-butyl Phthalate Production by Filamentous Fungi. Sci. Rep. 2016, 6, 19791. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
General chemical structure of phthalate esters.

Table 1.
Most common phthalates with acronyms, molecular formulas, CAS, R, and R′ chains and their log Kow.
Table 1.
Most common phthalates with acronyms, molecular formulas, CAS, R, and R′ chains and their log Kow.
| PAEs | Acronym | Molecular Formula | R | R′ | Log Kow |
|---|---|---|---|---|---|
| dimethyl phthalate | DMP | C10H10O4 | CH3 | CH3 | 1.60 |
| diethyl phthalate | DEP | C12H14O4 | CH2CH3 | CH2CH3 | 2.47 |
| mono(2-ethylhexyl) phthalate | MEHP | C16H22O4 | CH(CH2)5(CH3)2 | H | 4 * |
| dibutyl phthalate | DnBP | C16H22O4 | CH2CH2CH2CH3 | CH2CH2CH2CH3 | 4.50 |
| diisobutyl phthalate | DIBP | C16H22O4 | CH2CH(CH3)2 | CH2CH(CH3)2 | 4.11 |
| butyl isobutyl phthalate | BIBP | C16H22O4 | CH2CH(CH3)2 | CH2CH2CH2CH3 | 4.8 * |
| di(2-methylbutyl) phthalate | DMBP | C18H26O4 | CH(CH2)2(CH3)2 | CH(CH2)2(CH3)2 | - |
| benzyl butyl phthalate | BBzP | C19H20O4 | CH2C6H5 | CH2C6H5 | 4.73 |
| di(2-ethylhexyl) phthalate | DEHP | C24H38O4 | CH(CH2)5(CH3)2 | CH(CH2)5(CH3)2 | 7.60 |
| di-n-octyl phthalate | DnOP | C24H38O4 | (CH2)7CH3 | (CH2)7CH3 | 8.10 |
| diisooctyl phthalate | DIOP | C24H38O4 | CH(CH2)5(CH3)2 | CH(CH2)5(CH3)2 | 8.5 * |
| di-isononyl phthalate | DINP | C26H42O4 | CH(CH2)6(CH3)2 | CH(CH2)6(CH3)2 | 9.6 * |
| di-n-decyl phthalate | DnDP | C28H46O4 | (CH2)8CH3 | (CH2)8CH3 | 11.2 * |
Log Kow values were obtained from PubChem [5]. * Computed by XLogP3 3.0 (PubChem release 14 October 2021).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pace, A.; Vaglica, A.; Maccotta, A.; Savoca, D. The Origin of Phthalates in Algae: Biosynthesis and Environmental Bioaccumulation. Environments 2024, 11, 78. https://doi.org/10.3390/environments11040078
AMA Style
Pace A, Vaglica A, Maccotta A, Savoca D. The Origin of Phthalates in Algae: Biosynthesis and Environmental Bioaccumulation. Environments. 2024; 11(4):78. https://doi.org/10.3390/environments11040078
Chicago/Turabian StylePace, Andrea, Alessandro Vaglica, Antonella Maccotta, and Dario Savoca. 2024. "The Origin of Phthalates in Algae: Biosynthesis and Environmental Bioaccumulation" Environments 11, no. 4: 78. https://doi.org/10.3390/environments11040078
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.