Blocking Effect of Chaga Mushroom (Inonotus oliquus) Extract for Immune Checkpoint CTLA-4/CD80 Interaction



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extraction
2.3. Instruments
2.4. Preparation of Standard and Sample Solutions
2.5. Chemicals and Antibodies
2.6. Competitive ELISA
2.7. CTLA-4 Blockade Bioassay
2.8. HPLC Analytical Methods
2.9. Cell Viability in CTLA-4 Jurkat T Cells and CD80/aAPC CHO-K1 Cells
3. Results
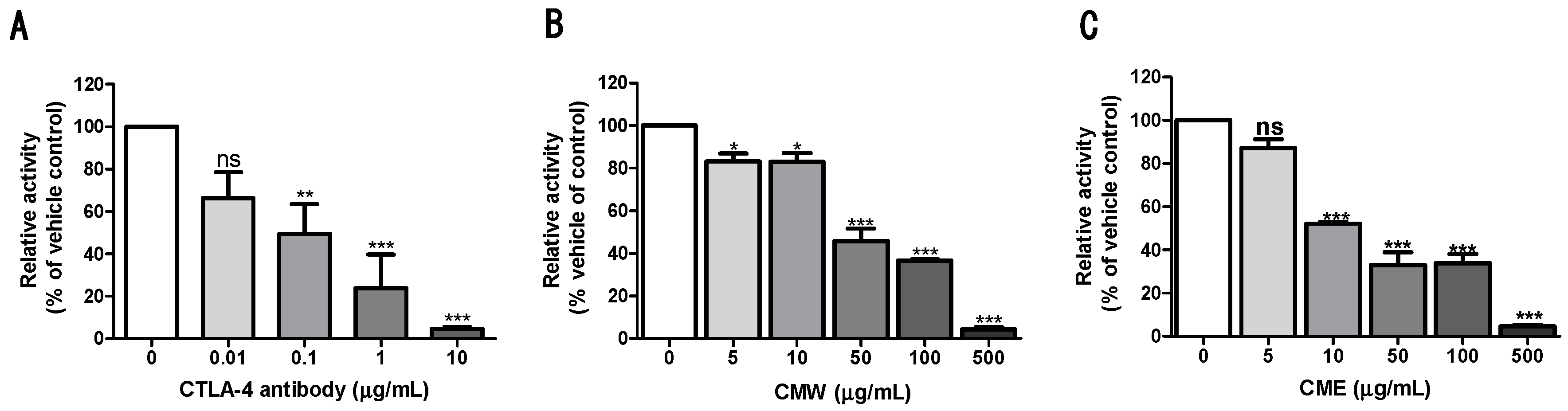
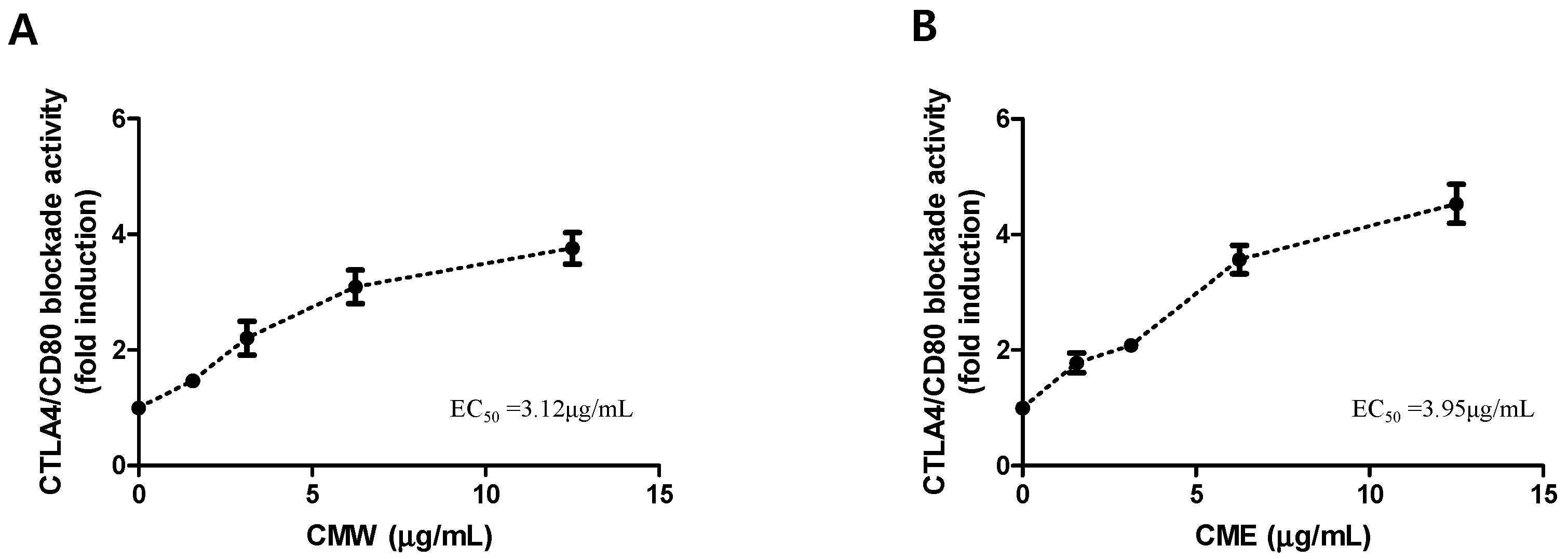
3.1. Chaga Mushroom Extract-Blocked CTLA-4/CD80 Interaction
3.2. NFAT Reporter Activity of the Chaga Mushroom Extract
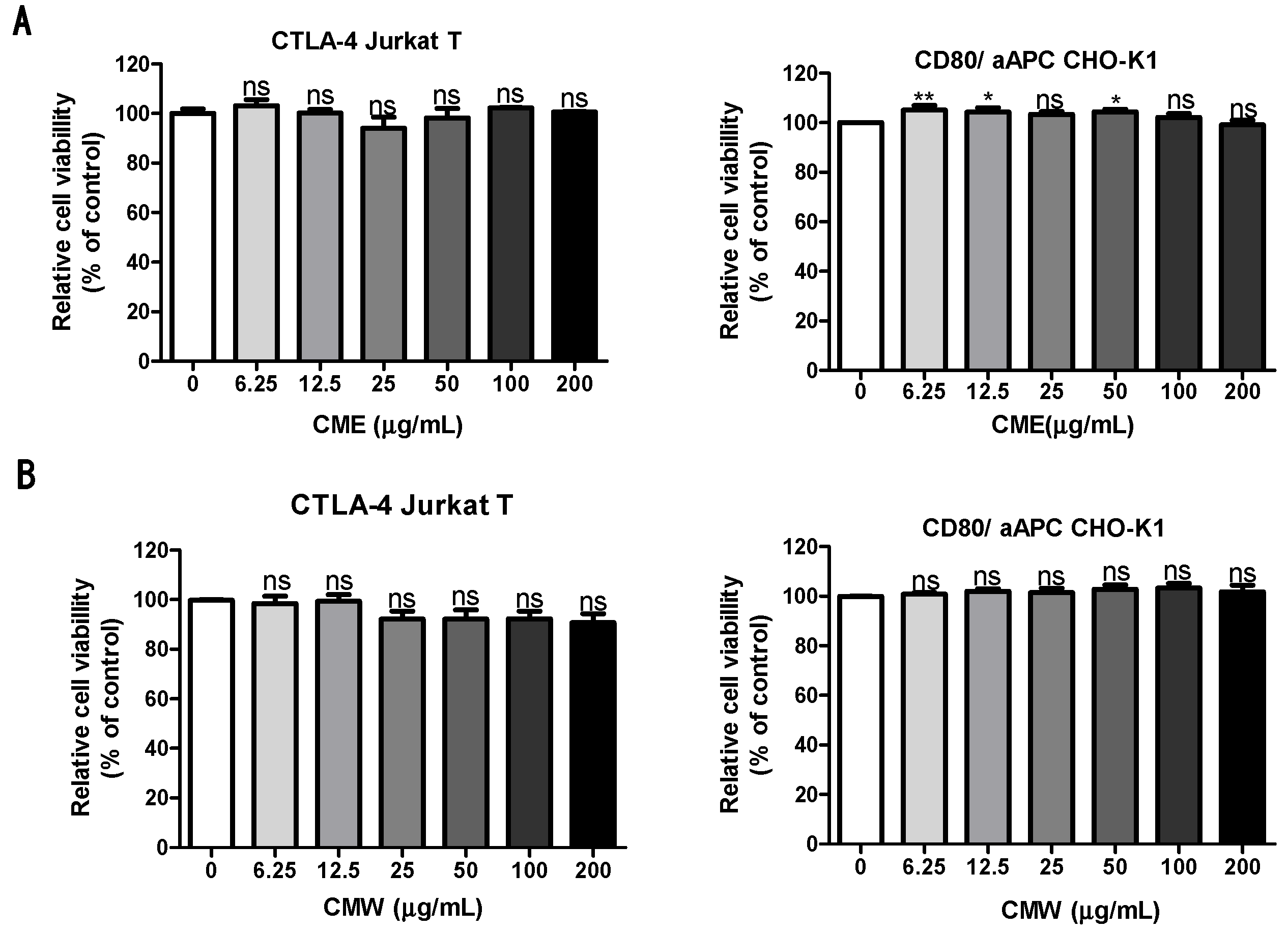
3.3. Evaluation of Cytotoxicity of the Chaga Mushroom Extract in CTLA-4 Jurkat T and CD80/aAPC CHO-K1 Cells
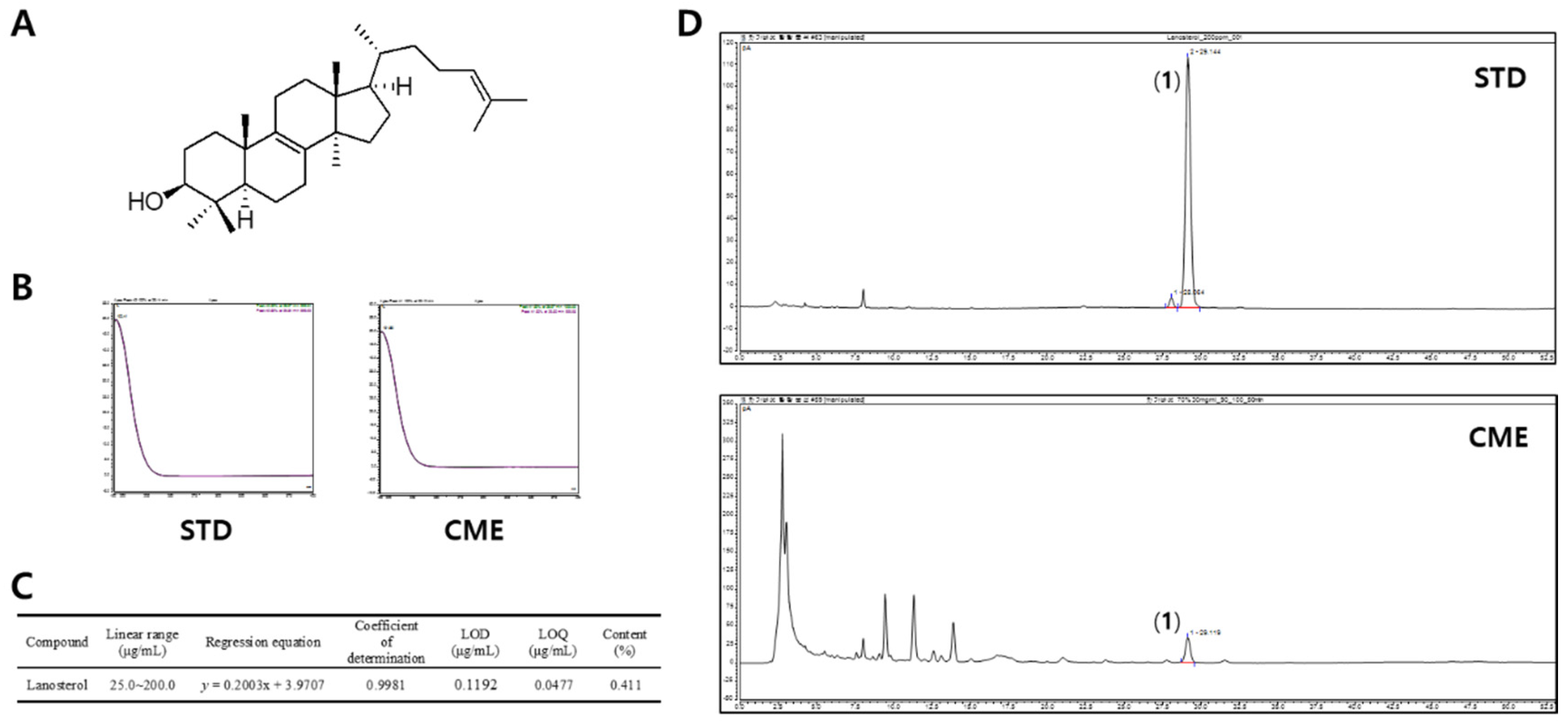
3.4. Content Analysis of Lanosterol (1)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Won, J.H.; Kim, Y.H.; Choi, J.W.; Park, H.J.; Lee, K.T. In vivo and in vitro anti-inflammatory and anti-nociceptive effects of the methanol extract of Inonotus obliquus. J. Ethnopharmacol. 2005, 101, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, E.J.; Kim, S.H. Ethanol extract of Innotus obliquus (Chaga mushroom) induces G1 cell cycle arrest in HT-29 human colon cancer cells. Nutr Res. Pract. 2015, 9, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Roh, H.S.; Baek, K.H.; Lee, S.; Lee, S.; Song, S.S.; Kim, K.H. Bioactivity-based analysis and chemical characterization of cytotoxic constituents from Chaga mushroom (Inonotus obliquus) that induce apoptosis in human lung adenocarcinoma cells. J. Ethnopharmacol. 2018, 224, 63–75. [Google Scholar] [CrossRef]
- He, J.; Feng, X.Z.; Lu, Y.; Zhao, B. Three new triterpenoids from Fuscoporia obliqua. J. Asian Natl. Prod. Res. 2001, 3, 55–61. [Google Scholar] [CrossRef]
- Glamoclija, J.; Ciric, A.; Nikolic, M.; Fernandes, A.; Barros, L.; Calhelha, R.C.; Ferreira, I.C.; Sokovic, M.; van Griensven, L.J. Chemical characterization and biological activity of Chaga (Inonotus obliquus), a medicinal “mushroom”. J. Ethnopharmacol. 2015, 162, 323–332. [Google Scholar] [CrossRef]
- Song, Y.; Hui, J.; Kou, W.; Xin, R.; Jia, F.; Wang, N.; Hu, F.; Zhang, H.; Liu, H. Identification of Inonotus obliquus and analysis of antioxidation and antitumor activities of polysaccharides. Curr. Microbiol. 2008, 57, 454–462. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, J.; Min, B.S.; Shim, S.H. Chemical Constituents from the Sclerotia of Inonotus obliquus. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 287–294. [Google Scholar] [CrossRef]
- Li, W.; Kim, T.I.; Kim, J.H.; Chung, H.S. Immune Checkpoint PD-1/PD-L1 CTLA-4/CD80 are Blocked by Rhus verniciflua Stokes and its Active Compounds. Molecules 2019, 24, 4062. [Google Scholar] [CrossRef]
- June, C.H.; Warshauer, J.T.; Bluestone, J.A. Is autoimmunity the Achilles’ heel of cancer immunotherapy? Nat. Med. 2017, 23, 540–547. [Google Scholar] [CrossRef]
- Sasikumar, P.G.; Ramachandra, M. Small-Molecule Immune Checkpoint Inhibitors Targeting PD-1/PD-L1 and Other Emerging Checkpoint Pathways. BioDrugs 2018, 32, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Kourie, H.R.; Klastersky, J. Immune checkpoint inhibitors side effects and management. Immunotherapy 2016, 8, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Linsley, P.S.; Brady, W.; Urnes, M.; Grosmaire, L.S.; Damle, N.K.; Ledbetter, J.A. CTLA-4 is a second receptor for the B cell activation antigen B7. J. Exp. Med. 1991, 174, 561–569. [Google Scholar] [CrossRef]
- Hurwitz, A.A.; Foster, B.A.; Kwon, E.D.; Truong, T.; Choi, E.M.; Greenberg, N.M.; Burg, M.B.; Allison, J.P. Combination immunotherapy of primary prostate cancer in a transgenic mouse model using CTLA-4 blockade. Cancer Res. 2000, 60, 2444–2448. [Google Scholar] [PubMed]
- Youn, M.J.; Kim, J.K.; Park, S.Y.; Kim, Y.; Park, C.; Kim, E.S.; Park, K.I.; So, H.S.; Park, R. Potential anticancer properties of the water extract of Inonotus obliquus by induction of apoptosis in melanoma B16-F10 cells. J. Ethnopharmacol. 2009, 121, 221–228. [Google Scholar] [CrossRef]
- Lemieszek, M.K.; Langner, E.; Kaczor, J.; Kandefer-Szerszen, M.; Sanecka, B.; Mazurkiewicz, W.; Rzeski, W. Anticancer effects of fraction isolated from fruiting bodies of Chaga medicinal mushroom, Inonotus obliquus (Pers.:Fr.) Pilat (Aphyllophoromycetideae): In vitro studies. Int J. Med. Mushrooms 2011, 13, 131–143. [Google Scholar] [CrossRef]
- Youn, M.J.; Kim, J.K.; Park, S.Y.; Kim, Y.; Kim, S.J.; Lee, J.S.; Chai, K.Y.; Kim, H.J.; Cui, M.X.; So, H.S.; et al. Chaga mushroom (Inonotus obliquus) induces G0/G1 arrest and apoptosis in human hepatoma HepG2 cells. World J. Gastroenterol. 2008, 14, 511–517. [Google Scholar] [CrossRef]
- Nguyet, T.M.N.; Lomunova, M.; Le, B.V.; Lee, J.S.; Park, S.K.; Kang, J.S.; Kim, Y.H.; Hwang, I. The mast cell stabilizing activity of Chaga mushroom critical for its therapeutic effect on food allergy is derived from inotodiol. Int. Immunopharmacol. 2018, 54, 286–295. [Google Scholar] [CrossRef]
- Niu, H.; Song, D.; Mu, H.; Zhang, W.; Sun, F.; Duan, J. Investigation of three lignin complexes with antioxidant and immunological capacities from Inonotus obliquus. Int. J. Biol. Macromol. 2016, 86, 587–593. [Google Scholar] [CrossRef]
- Won, D.P.; Lee, J.S.; Kwon, D.S.; Lee, K.E.; Shin, W.C.; Hong, E.K. Immunostimulating activity by polysaccharides isolated from fruiting body of Inonotus obliquus. Mol. Cells 2011, 31, 165–173. [Google Scholar] [CrossRef]
- Rios, J. Effects of triterpenes on the immune system. J. Ethnopharmacol. 2010, 128, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Mai, Q.; Ma, J.; Xu, M.; Wang, X.; Cui, T.; Qiu, F.; Han, G. Triterpenoids from Inonotus obliquus and their antitumor activities. Fitoterapia 2015, 101, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, S.A.; Habibrakhmanova, V.R.; Sysoeva, M.A. Chemical Composition and Biological Activity of Triterpenes and Steroids of Chaga Mushroom. Biomed. Khim 2016, 10, 63–69. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Seta, K.; Ogawa, Y.; Takayama, T.; Nagata, M.; Taguchi, T.; Yahata, K. Chaga mushroom-induced oxalate nephropathy. Clin. Nephrol. 2014, 81, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Hyun, K.W.; Jeong, S.C.; Lee, D.H.; Park, J.S.; Lee, J.S. Isolation and characterization of a novel platelet aggregation inhibitory peptide from the medicinal mushroom, Inonotus obliquus. Peptides 2006, 27, 1173–1178. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.I.; Choi, J.-G.; Kim, J.H.; Li, W.; Chung, H.-S. Blocking Effect of Chaga Mushroom (Inonotus oliquus) Extract for Immune Checkpoint CTLA-4/CD80 Interaction. Appl. Sci. 2020, 10, 5774. https://doi.org/10.3390/app10175774
Kim TI, Choi J-G, Kim JH, Li W, Chung H-S. Blocking Effect of Chaga Mushroom (Inonotus oliquus) Extract for Immune Checkpoint CTLA-4/CD80 Interaction. Applied Sciences. 2020; 10(17):5774. https://doi.org/10.3390/app10175774
Chicago/Turabian StyleKim, Tae In, Jang-Gi Choi, Ji Hye Kim, Wei Li, and Hwan-Suck Chung. 2020. "Blocking Effect of Chaga Mushroom (Inonotus oliquus) Extract for Immune Checkpoint CTLA-4/CD80 Interaction" Applied Sciences 10, no. 17: 5774. https://doi.org/10.3390/app10175774
APA StyleKim, T. I., Choi, J.-G., Kim, J. H., Li, W., & Chung, H.-S. (2020). Blocking Effect of Chaga Mushroom (Inonotus oliquus) Extract for Immune Checkpoint CTLA-4/CD80 Interaction. Applied Sciences, 10(17), 5774. https://doi.org/10.3390/app10175774