Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus mutans Biofilm in Dental Plaque

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Bacteria Strain
2.2. Preparation of Protein Structure, Ligand Structure and Active Site Prediction
2.3. Molecular Docking
2.4. Anti-microbial Activity of Mangiferin and Time-Kill Assay
2.5. Biofilm Susceptibility Assay
2.6. The Assay of Glucansucrase Enzyme Inhibition
2.7. Scanning Electron Microscopic Analysis
2.8. Analysis of Bacterial Adhesion and Exopolysaccharide (EPS) Synthesis
2.9. RNA Isolation, Reverse Transcription, and Quantitative Real-Time PCR
2.10. Hemo Toxicity
2.11. Oral cell Toxicity
MTT Assay and Cytotoxicity Analysis
2.12. Data Analysis
3. Results
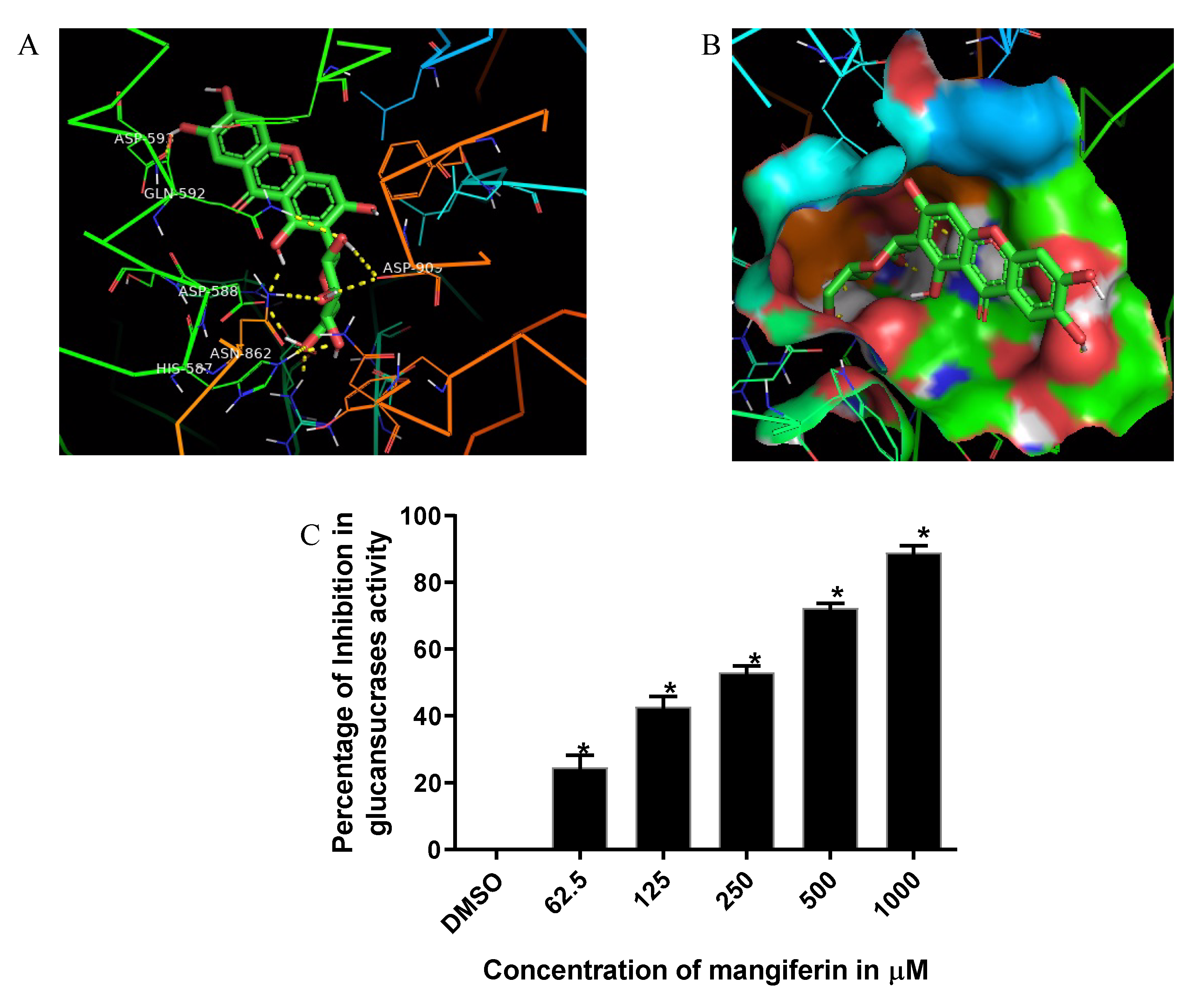
3.1. Computational Analysis of S. mutans Glucansucrase Binding Potential of Mangiferin
3.2. Mangiferin Inhibition of S. mutans Glucansucrase Activity
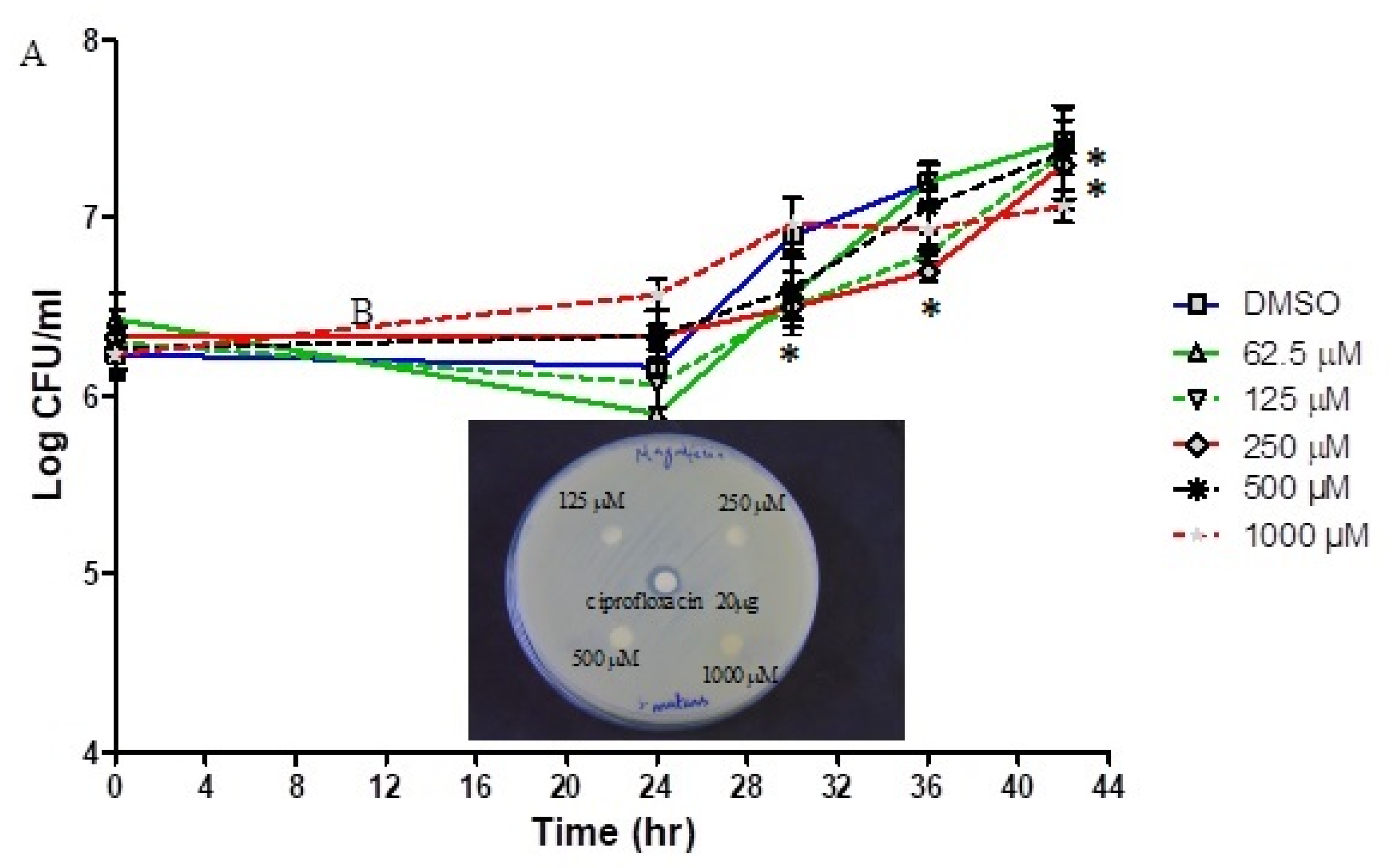
3.3. Antibacterial Activity of Mangiferin against Planktonic S. mutans
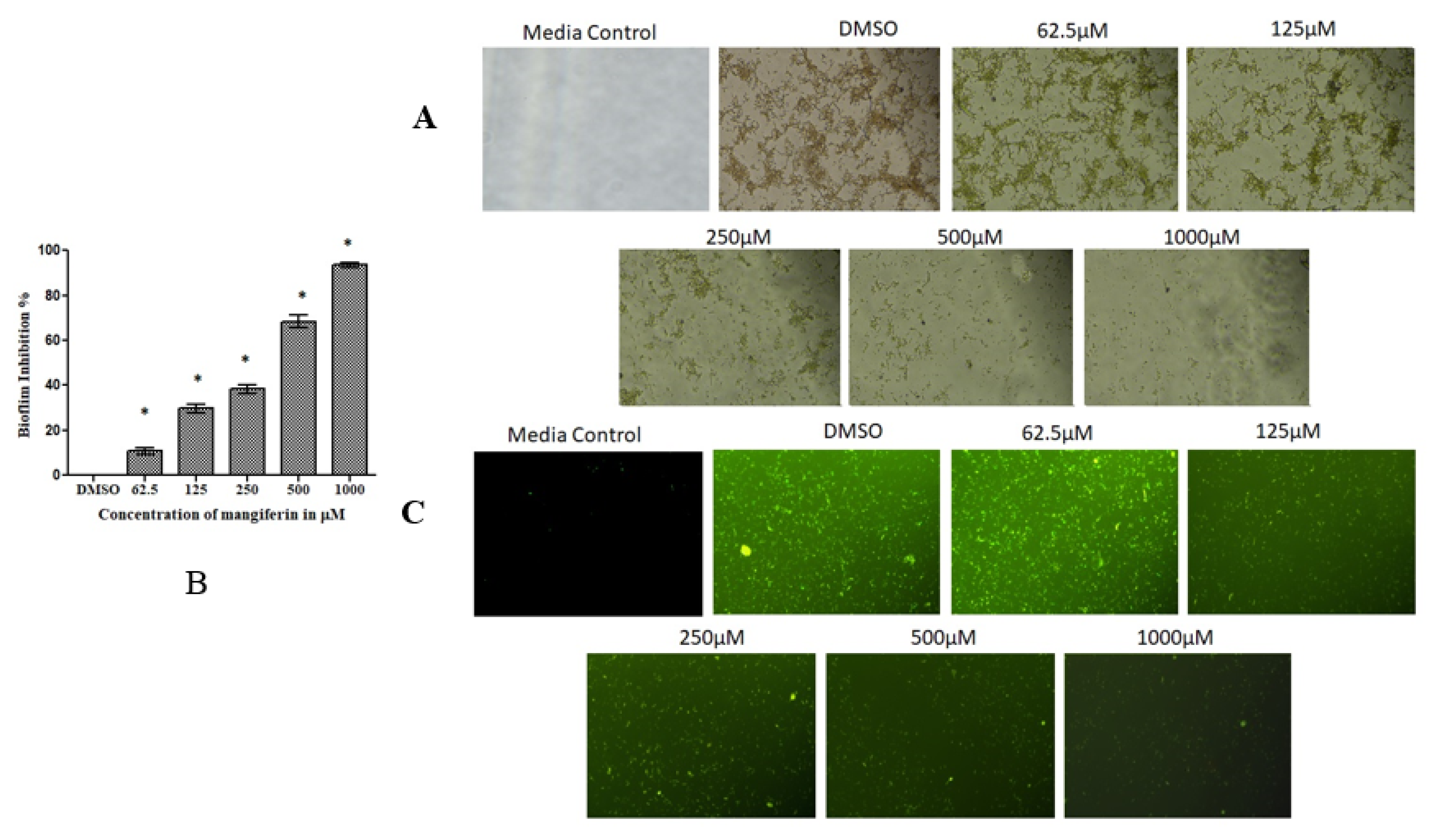
3.4. Mangiferin Suppressed S. mutans Biofilm
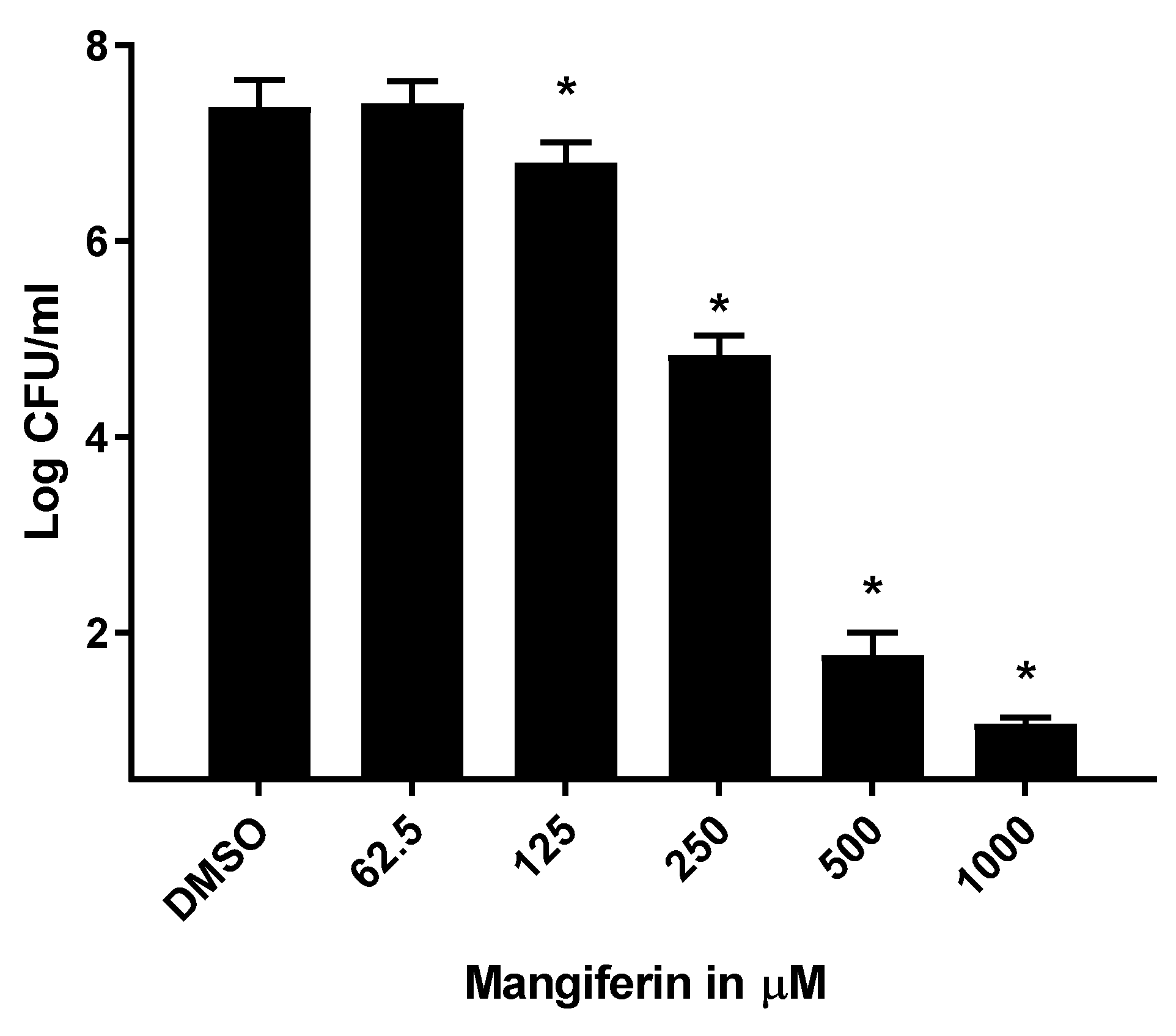
3.5. Mangiferin Suppressed S. mutans Biofilm on Dental Surface
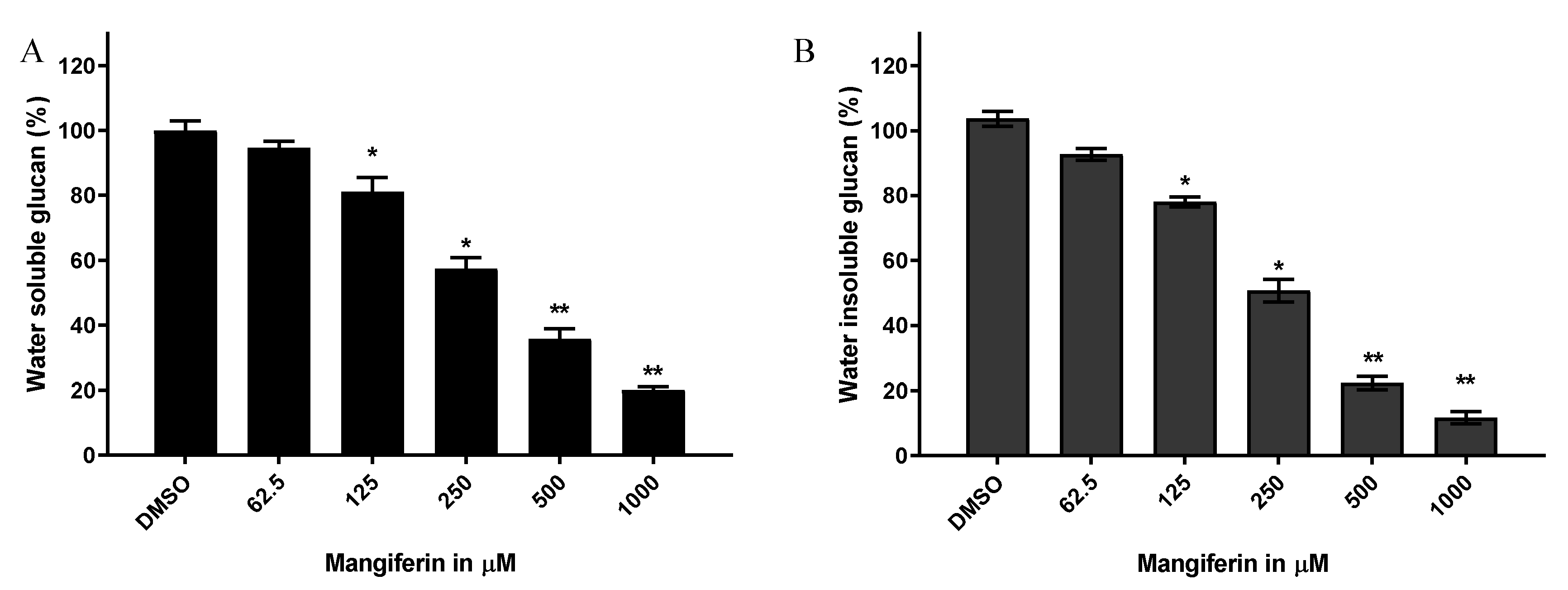
3.6. Mangiferin Inhibition of Glucan Synthesis in S. mutans Biofilm
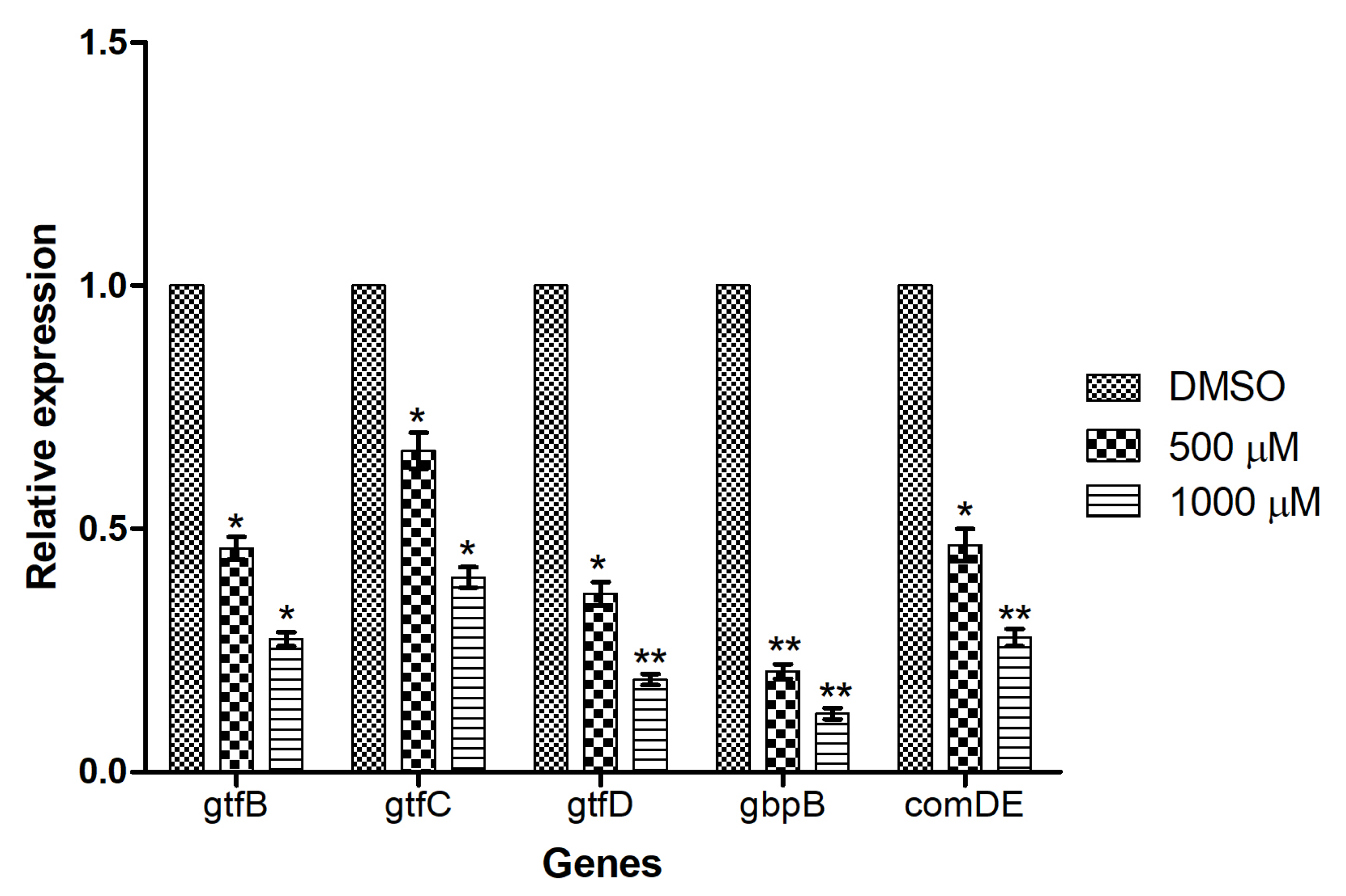
3.7. Mangiferin Down-Regulated the Expression of Genes Involved in Biofilm Formation
3.8. Scanning Electron Microscopic Observation of Biofilm
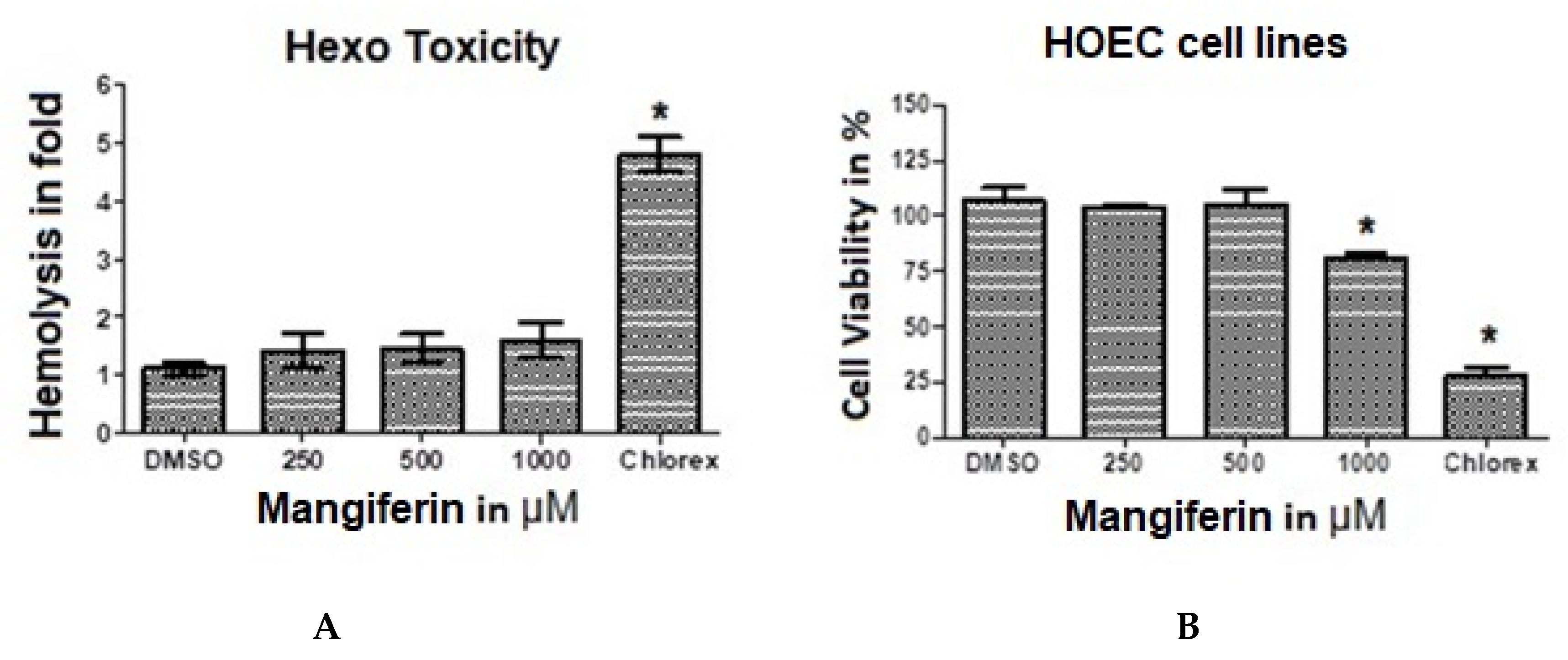
3.9. Hemo Toxicity
3.10. MTT Assay and Cytotoxicity Analysis
4. Discussion
5. Conclusions
6. Study Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Oliveira, J.S.; Pinto, M.E.; Santana, L.A.; Pinto, A.S.; Di Lenardo, D.; Vasconcelos, D.F. Biological effects of medicinal plants on induced periodontitis: A systematic review. Int. J. Dent. 2016, 3719879. [Google Scholar] [CrossRef]
- Lin, N.J. Biofilm over teeth and restorations: What do we need to know? Dent. Mater. 2017, 33, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Mira, A.; Simon-Soro, A.; Curtis, M.A. Role of microbial communities in the pathogenesis of periodontal diseases and caries. J. Clin. Periodontol. 2017, 44, S23–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Burden of Disease (GBD) 2015. Disease and injury incidence and prevalence collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aziz, M.M.; Emam, T.M.; Raafat, M.M. Hindering of cariogenic Streptococcus mutans biofilm by fatty acid array derived from an endophytic Arthrographis kalrae strain. Biomolecules 2020, 10, 811. [Google Scholar] [CrossRef]
- Moynihan, P. Sugars and dental caries: Evidence for setting a recommended threshold for intake. Adv. Nutr. 2016, 7, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Kuang, X.; Zhou, Y.; Peng, X.; Guo, Q.; Yang, T.; Zhou, X.; Luo, Y.; Xu, X. A Novel small molecule, ZY354, inhibits dental caries-associated oral biofilms. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Marsh, P.D. Microbiology of dental plaque biofilms and their role in oral health and caries. Dent. Clin. N. Am. 2010, 54, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, M.; Wang, Y.; Zhou, X.; Peng, X.; Ren, B.; Li, M.; Cheng, L. Effect of Veillonella parvula on the physiological activity of Streptococcus mutans. Arch. Oral. Biol. 2020, 109, 104578. [Google Scholar] [CrossRef]
- Chen, L.; Ren, Z.; Zhou, X.; Zeng, J.; Zou, J.; Li, Y. Inhibition of Streptococcus mutans biofilm formation, extracellular polysaccharide production, and virulence by an oxazole derivative. Appl. Microbiol. Biotechnol. 2016, 100, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; McLean, J.S.; Lux, R.; He, X.; Shi, W. The well-coordinated linkage between acidogenicity and aciduricity via insoluble glucans on the surface of Streptococcus mutans. Sci Rep. 2015, 10, 18015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Danishuddin, M.; Khan, A.U. Inhibitory effect of Zingiber officinale towards Streptococcus mutans virulence and caries development: In vitro and in vivo studies. BMC Microbiol. 2015, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Krzyściak, W.; Jurczak, A.; Kościelniak, D.; Bystrowska, B.; Skalniak, A. The virulence of Streptococcus mutans and the ability to form biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 499–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Balamurugan, P.; Princy, S.A. Inhibition of the Quorum Sensing System (ComDE Pathway) by Aromatic 1,3-di-m-tolylurea (DMTU): Cariostatic Effect with Fluoride in Wistar Rats. Front. Cell. Infect. Microbiol. 2017, 7, 313. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; van der Mei, H.C.; Busscher, H.J.; Ren, Y. Streptococcus mutans adhesion force sensing in multi-species oral biofilms. NPJ Biofilms Microbiomes 2020, 6, 25. [Google Scholar] [CrossRef]
- Walsh, T.; Oliveira-Neto, J.M.; Moore, D. Chlorhexidine treatment for the prevention of dental caries in children and adolescents. Cochrane database. Syst. Rev. 2015, 4, CD008457. [Google Scholar] [CrossRef]
- Cabral, C.T.; Fernandes, M.H. In vitro comparison of chlorhexidine and povidone–iodine on the long-term proliferation and functional activity of human alveolar bone cells. Clin. Oral Investig. 2007, 11, 155–164. [Google Scholar] [CrossRef]
- Nijampatnam, B.; Zhang, H.; Cai, X.; Michalek, S.M.; Wu, H.; Velu, S.E. Inhibition of Streptococcus mutans Biofilms by the natural stilbene piceatannol through the inhibition of glucosyltransferases. ACS Omega 2018, 3, 8378–8385. [Google Scholar] [CrossRef] [Green Version]
- Telang, M.; Dhulap, S.; Mandhare, A.; Hirwani, R. Therapeutic and cosmetic applications of mangiferin: A patent review. Expert. Opin. Ther. Pat. 2013, 23, 1561–1580. [Google Scholar] [CrossRef]
- Jangra, A.; Arora, M.K.; Kisku, A. The multifaceted role of mangiferin in health and diseases: A review. Adv. Tradit. Med. 2020. [Google Scholar] [CrossRef]
- Carvalho, R.R.; Pellizzon, C.H.; Justulin, L., Jr.; Felisbino, S.L.; Vilegas, W.; Bruni, F.; Lopes-Ferreira, M.; Hiruma-Lima, C.A. Effect of mangiferin on the development of periodontal disease: Involvement of lipoxin A4, anti-chemotaxic action in leukocyte rolling. Chem. Biol. Interact. 2009, 179, 344–350. [Google Scholar] [PubMed]
- Wang, R.R.; Gao, Y.D.; Ma, C.H.; Zhang, X.J.; Huang, C.G.; Huang, J.F. Mangiferin, an Anti-HIV-1 Agent Targeting Protease and Effective against Resistant Strains. Molecules 2011, 16, 4264–4277. [Google Scholar] [PubMed]
- Nordini, W.H.W.I.; Fathilah, A.R.; Rahim, Z.H.A. Plant extracts of Psidium guajava, Mangifera sp. and Mentha sp. inhibit the growth of the population of single-species oral biofilm. Altern. Complement. Ther. 2013, 2, 102–106. [Google Scholar] [CrossRef]
- Abdul, R.Z.H.; Shaikh, S.; Hasnor, W.I.W.N.; Wan Harun, W.H.A.; Razak, F.A. The effect of selected plant extracts on the development of single-species dental biofilms. J. Coll. Physicians Surg. Pak. 2014, 24, 796–801. [Google Scholar]
- Isola, G.; Polizzi, A.; Patini, R.; Ferlito, S.; Alibrandi, A.; Palazzo, G. Association among serum and salivary A. actinomycetemcomitans specific immunoglobulin antibodies and periodontitis. BMC Oral Health 2020, 20, 283. [Google Scholar] [CrossRef]
- Quang, N.V.; Dang, V.L.; Do, T.X.; Quach, M.T.; Nguyen, T.T.; Nguyen, Q.T.; Thanh, T.T.; Nguyen, T.M.P. Anticaries activity of mangiferin isolated from Mangifera indica leaves in Vietnam. Vietnam J. Chem. 2017, 55, 623–626. [Google Scholar] [CrossRef]
- Isola, G.; Polizzi, A.; Iorio-Siciliano, V.; Alibrandi, A.; Ramaglia, L.; Leonardi, R. Effectiveness of a nutraceutical agent in the non-surgical periodontal therapy: A randomized, controlled clinical trial. Clin. Oral Investig. 2020. [Google Scholar] [CrossRef]
- Hairul-Islam, M.I.; Arokiyaraj, S.; Kuralarasan, M.; Senthil Kumar, V.; Harikrishnan, P.; Saravanan, S.; Ashok, G.; Chellappandian, M.; Bharanidharan, R.; Muralidaran, S.; et al. Inhibitory potential of EGCG on Streptococcus mutans biofilm: A new approach to prevent Cariogenesis. 2020, 143, 104129. [Google Scholar]
- Bai, L.; Takagi, S.; Ando, T.; Yoneyama, H.; Ito, K.; Mizugai, H.; Isogai, E. Antimicrobial activity of tea catechin against canine oral bacteria and the functional mechanisms. J. Vet. Med. Sci. 2016, 78, 1439–1445. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhou, X.D.; Wu, C.D. The Tea Catechin Epigallocatechin Gallate Suppresses Cariogenic Virulence Factors of Streptococcus mutans. Antimicrob. Agents Chemother. 2011, 55, 1229–1236. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Goyal, A. 16S rRNA-based identification of a glucan-hyperproducing Weissella confusa. Enzym. Res. 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Yeon, M.J.; Choi, N.S.; Chang, Y.H.; Jung, M.Y.; Song, J.J.; Kim, J.S. Purification and characterization of a novel glucansucrase from leuconostoc lactis EG001. Microbiol. Res. 2010, 165, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Laosuwan, K.; Epasinghe, D.J.; Wu, Z.; Leung, W.K.; Green, D.W.; Jung, H.S. Comparison of biofilm formation and migration of Streptococcus mutans on tooth roots and titanium miniscrews. Clin. Exp. Dent. Res. 2018, 4, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Kim, J.E.; Jung, K.H.; Chang, K.W.; Jeon, J.G. Effect of sodium fluoride on the virulence factors and composition of Streptococcus mutans biofilms. Arch. Oral Biol. 2011, 56, 643–649. [Google Scholar] [CrossRef]
- Koo, H.; Schobel, B.; Scott-Anne, K.; Watson, G.; Bowen, W.H.; Cury, J.A.; Rosalen, P.L.; Park, Y.K. Apigenin and tt -Farnesol with Fluoride Effects on S. mutans Biofilms and Dental Caries. J. Dent. Res. 2005, 84, 1016–1020. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shafiei, Z.; Haji, A.R.Z.; Philip, K.; Thurairajah, N. Antibacterial and anti-adherence effects of a plant extract mixture (PEM) and its individual constituent extracts (Psidium sp., Mangifera sp., and Mentha sp.) on single- and dual-species biofilms. PEERJ 2016, 4, e2519. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Huang, Z.; Jiang, W.; Zhou, W. Antimicrobial Activity of Cinnamaldehyde on Streptococcus mutans Biofilms. Front Microbiol. 2019, 10, 2241. [Google Scholar] [CrossRef] [Green Version]
- Lemos, J.A.; Palmer, S.R.; Zeng, L.; Wen, Z.T.; Kajfasz, J.K.; Freires, I.A.; Abranches, J.; Brady, L.J. The biology of Streptococcus mutans. Microbiol Spectr. 2019, 7, 435–448. [Google Scholar] [CrossRef]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three-dimensional structures, reactions, mechanism, α-glucan analysis and their implications in biotechnology and food applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Gangoiti, J.; Bai, Y.; Pijning, T.; Van Leeuwen, S.S.; Dijkhuizen, L. Structure–function relationships of family GH70 glucansucrase and 4,6-α-glucanotransferase enzymes, and their evolutionary relationships with family GH13 enzymes. Cell. Mol. Life Sci. 2016, 73, 2681–2706. [Google Scholar] [PubMed] [Green Version]
- Ito, K.; Ito, S.; Shimamura, T.; Weyand, S.; Kawarasaki, Y.; Misaka, T.; Abe, K.; Kobayashi, T.; Cameron, A.D.; Iwata, S. Crystal structure of glucansucrase from the dental caries pathogen Streptococcus mutans. J. Mol. Biol. 2011, 408, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, L.; Mendoza, F.; Alderete, J.B.; Jiménez, V.A.; Jaña, G.A. The role of con-served arginine in the GH70 family: A computational study of the structural features and their implications on the catalytic mechanism of GTF-SI from Streptoccocus mutans. Org. Biomol. Chem. 2019, 17, 6269–6276. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Chen, L.; Li, J.; Li, Y. Inhibition of Streptococcus mutans polysaccharide synthesis by molecules targeting glycosyltransferase activity. J. Oral Microbiol. 2016, 8, 31095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.; Xiao, J.; Klein, M.I.; Jeon, J.G. Exopolysaccharides produced by Streptococcus mutans glucosyltransferases modulate the establishment of micro colonies within multispecies biofilms. J. Bacteriol. 2010, 192, 302432. [Google Scholar]
- Banavar, R.S.; Nirupad, S.; Chippagiri, P.; Pandurangappa, R. Antibacterial Effects of Natural Herbal Extracts on Streptococcus mutans: Can They Be Potential Additives in Dentifrices? Int. J. Dent. 2017, 4921614. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Donadu, M.G.; Ho, D.V.; Doan, T.Q.; Le, A.T.; Raal, A.; Usai, D.; Sanna, G.; Marchetti, M.; Usai, M.; et al. Biological activities of essential oil extracted from leaves of Atalantia sessiflora Guillauminin Vietnam. J. Infect. Dev. Ctries. 2020, 30, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Ács, K.; Balázs, V.L.; Kocsis, B.; Bencsik, T.; Böszörményi, A.; Horváth, G. Antibacterial activity evaluation of selected essential oils in liquid and vapor phase on respiratory tract pathogens. BMC Complement. Altern. Med. 2018, 27, 227. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi, B.; Rashidipour, M.; Marzban, A.; Soroush, S.; Azadpour, M.; Delfani, S.; Ramak, P. Mentha piperita essential oils loaded in a chitosan nanogel with inhibitory effect on biofilm formation against S. mutans on the dental surface. Carbohydr. Polym. 2019, 212, 142–149. [Google Scholar] [CrossRef]
- Isola, G.; Alibrandi, A.; Currò, M.; Matarese, M.; Ricca, S.; Matarese, G.; Ientile, R.; Kocher, T. Evaluation of salivary and serum ADMA levels in patients with periodontal and cardiovascular disease as subclinical marker of cardiovascular risk. J. Periodontol. 2020, 91, 1076–1084. [Google Scholar] [CrossRef]
- Isola, G.; Polizzi, A.; Alibrandi, A.; Indelicato, F.; Ferlito, S. Analysis of Endothelin-1 concentrations in individuals with periodontitis. Sci. Rep. 2020, 10, 1652. [Google Scholar] [PubMed]
- Gabe, V.; Kacergius, T.; Abu-Lafi, S.; Kalesinskas, P.; Masalha, M.; Falah, M.; Abu-Farich, B.; Melninkaitis, A.; Zeidan, M.; Rayan, A. Inhibitory effects of ethyl gallate on Streptococcus mutans biofilm formation by optical profilometry and gene expression analysis. Molecules 2019, 24, 529. [Google Scholar] [CrossRef] [Green Version]
- Duque, C.; Stipp, R.N.; Wang, B.; Smith, D.J.; Höfling, J.F.; Kuramitsu, H.K. Downregulation of Gbp B, a component of the Vic RK regulon, affects biofilm formation and cell surface characteristics of Streptococcus mutans. Infect. Immun. 2011, 79, 786–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedoya-Correa, C.M.; Rincón Rodríguez, R.J.; Parada-Sanchez, M.T. Genomic and phenotypic diversity of Streptococcus mutans. J. Oral Biosci. 2019, 61, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, T.; Peng, W.; Zhu, Y. Effects of resveratrol on cariogenic virulence properties of Streptococcus mutans. BMC Microbiol. 2020, 20, 99. [Google Scholar]
- Chakraborty, B.; Burne, R.A. Effects of Arginine on Streptococcus mutans Growth, virulence gene expression, and stress tolerance. Appl. Environ. Microbiol. 2017, 83, e00496. [Google Scholar] [CrossRef] [Green Version]
- Lei, Z.C.; Hu, H.; Hu, L.; Zhao, M.; Yang, Y.; Chuai, Y.; Ni, J.; Cai, J. Mangiferin aglycone attenuates radiation-induced damage on human intestinal epithelial cells. J. Cell. Biochem. 2012, 113, 2633–2642. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Amplicon Size (bp) | Role in Biofilm Formation |
|---|---|---|---|
| gtfB | FP: AGCAATGCAGCCAATCTACAAAT RP: ACGAACTTTGCCGTTATTGTCA | 95 | Water insoluble glucan production. |
| gtfC | FP: GTGCGCTACACCAATGACAGAG RP: GCCTACTGGAACCCAAACACCTA | 107 | Water soluble and insoluble glucan production. |
| gtfD | FP: TGGCACCGCAATATGTCTCTTC RP: CAATCCGCAATAACCTGAATACCG | 183 | Water soluble glucan production. |
| gbpB | FP: ATGGCGGTTATGGACACGTT RP: TTTGGCCACCTTGAACACCT | 50 | Glucan binding protein. |
| comDE | FP: ACAATTCCTTGAGTTCCATCCAAG RP: TGGTCTGCTGCCTGTTGC | 80 | Competence-stimulating peptide involved in quorum sensing mechanism |
| 16S | FP: CCTACGGGAGGCAGCAGTAG RP: CAACAGAGCTTTACGATCCGAAA | 100 |
| SL.No | Ligand | Binding Energy | Ligand Efficiency | Intermole Energy | Ligand Atoms (Ring) | Docked Amino Acid Residue (Bond Length) |
|---|---|---|---|---|---|---|
| 1. | Mangiferin | −7.35 | −0.25 | −8.66 | C7-OH (B) C1-OH (A) C2′-O C2′-OH C3′-OH C3′-O C4′-O C6′-O C6′-O C6′-OH | ASP‘593/OD1 (2.2 Å) ASP‘588/OD2 (2.0 Å) GLN‘592/1HE2 (2.4 Å) ASP‘909/OD1 (2.5 Å) ASP‘909/OD1 (2.1 Å) ASN‘862/1HD2 (2.5 Å) HIS‘587/HE2 (2.0 Å) HIS‘587/HE2 (2.4 Å) ARG‘475/1HH2 (2.0 Å) ASP‘588/OD2 (2.0 Å) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emeka, P.M.; Badger-Emeka, L.I.; Ibrahim, H.-I.M.; Thirugnanasambantham, K.; Hussen, J. Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus mutans Biofilm in Dental Plaque. Appl. Sci. 2020, 10, 8297. https://doi.org/10.3390/app10228297
Emeka PM, Badger-Emeka LI, Ibrahim H-IM, Thirugnanasambantham K, Hussen J. Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus mutans Biofilm in Dental Plaque. Applied Sciences. 2020; 10(22):8297. https://doi.org/10.3390/app10228297
Chicago/Turabian StyleEmeka, Promise M., Lorina I. Badger-Emeka, Hairul-Islam M. Ibrahim, Krishnaraj Thirugnanasambantham, and Jamal Hussen. 2020. "Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus mutans Biofilm in Dental Plaque" Applied Sciences 10, no. 22: 8297. https://doi.org/10.3390/app10228297
APA StyleEmeka, P. M., Badger-Emeka, L. I., Ibrahim, H.-I. M., Thirugnanasambantham, K., & Hussen, J. (2020). Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus mutans Biofilm in Dental Plaque. Applied Sciences, 10(22), 8297. https://doi.org/10.3390/app10228297