A Novel Evolution Strategy of Level Set Method for the Segmentation of Overlapping Cervical Cells

 ,
,  , and
, and
Abstract
1. Introduction
- (1)
- We designed a morphological scaling-based topology filter (MSTF) to filter out the false positive fragments caused by improper allocation of the initial contour points of touching cells (i.e., misallocation). In MSTF, we constructed the signed distance function (SDF) as the LSF of the initialized cytoplasm contour based on the linear time Euclidean distance transform (LTEDT) algorithm [62], which is denoted by LTEDT-SDF.
- (2)
- We theoretically derived a new mathematical toolbox about vector calculus for evolution of the LSF as the supplementary for the codimension two-level set method (CTLSM) [63], aiming to keep the initialized contours of the nonoverlapping region fixed. Our proposed evolution method of partially fixed contour is called the 2D codimension two-object level set method (DCTLSM), which can alleviate the accuracy loss of a MSTF.
- (3)
- We proposed a novel evolution strategy of LSF inspired by the watershed method [64]. In this strategy, we provided an effective guidance mechanism for attracting and repelling the LSF to converge towards its actual cell boundary.
- (4)
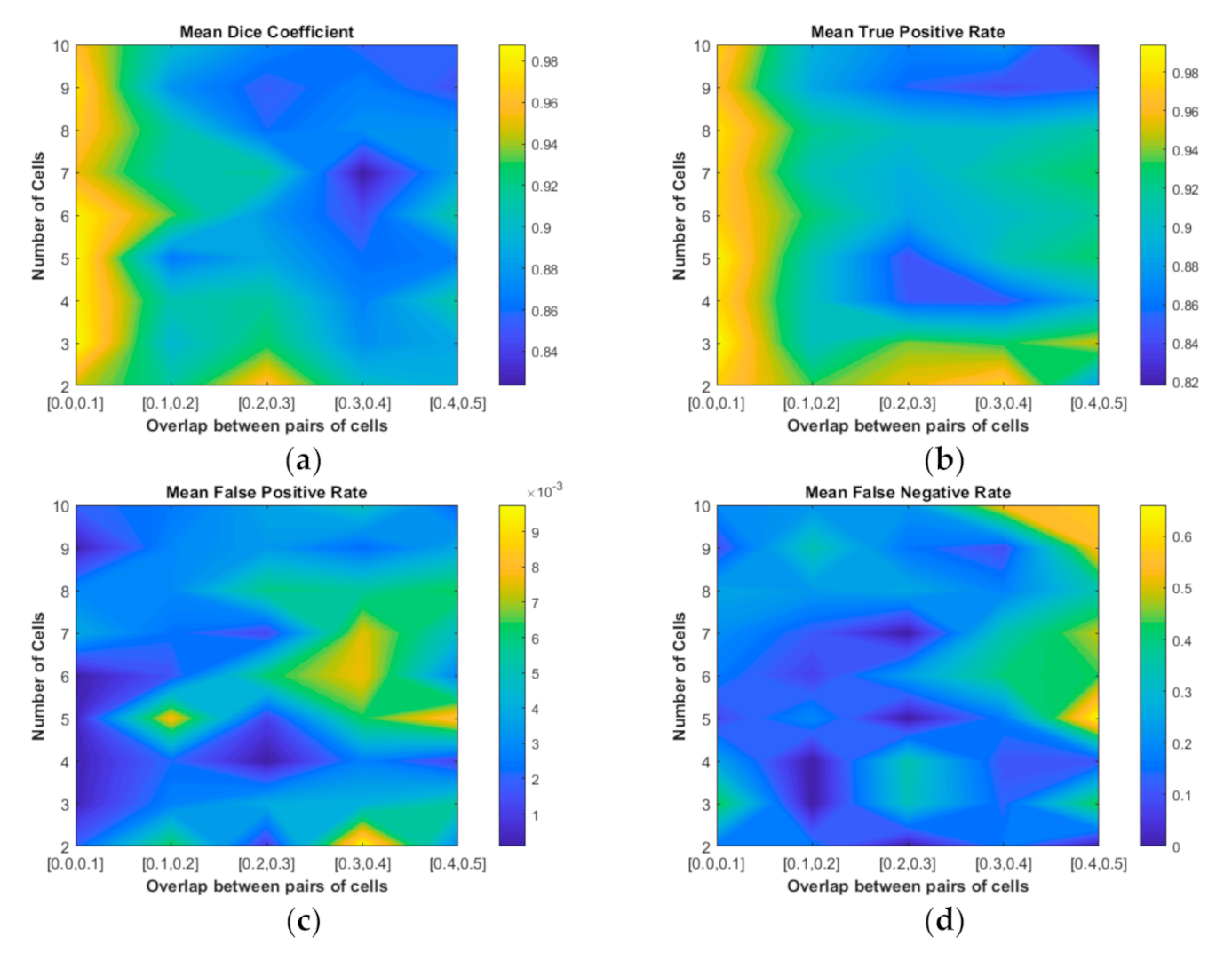
- We used the dataset published by the First Overlapping Cervical Cytology Image Segmentation Challenge held in conjunction with the IEEE International Symposium on Biomedical Imaging (ISBI-2014 challenge) to evaluate our proposed method. The experimental results showed that cellular clumps consisting of two to 10 cells under an overlap ratio less than 0.2 can be accurately segmented. Furthermore, the segmentation of cellular clumps consisting of two to four cells can be effectively segmented with an overlap ratio less than 0.5. By qualitive and quantitative comparisons, our method outperformed the other segmentation methods.
2. Methodology
2.1. Cellular Component Segmentation
2.2. Touching Cell Spliting
2.2.1. Morphological Scaling-Based Topology Filter
2.2.2. 2D Codimension Two-Object Level Set Method
2.3. Overlapping Cell Segmentation
2.3.1. Cutting Line Detection
2.3.2. Contour Scanning Strategy for Segmentation
3. Experiments
3.1. Image Datasets
3.2. Evaluation Metrics
4. Results and Discussion
4.1. The Determination of Morphological Scaling Threshold for MSTF and DCTLSM
4.2. Quantitative Evaluation of Our Segmentation Results
4.2.1. Quantitative Comparison with Baseline Method
4.2.2. Quantitative Comparison with The-State-of-The-Art Methods
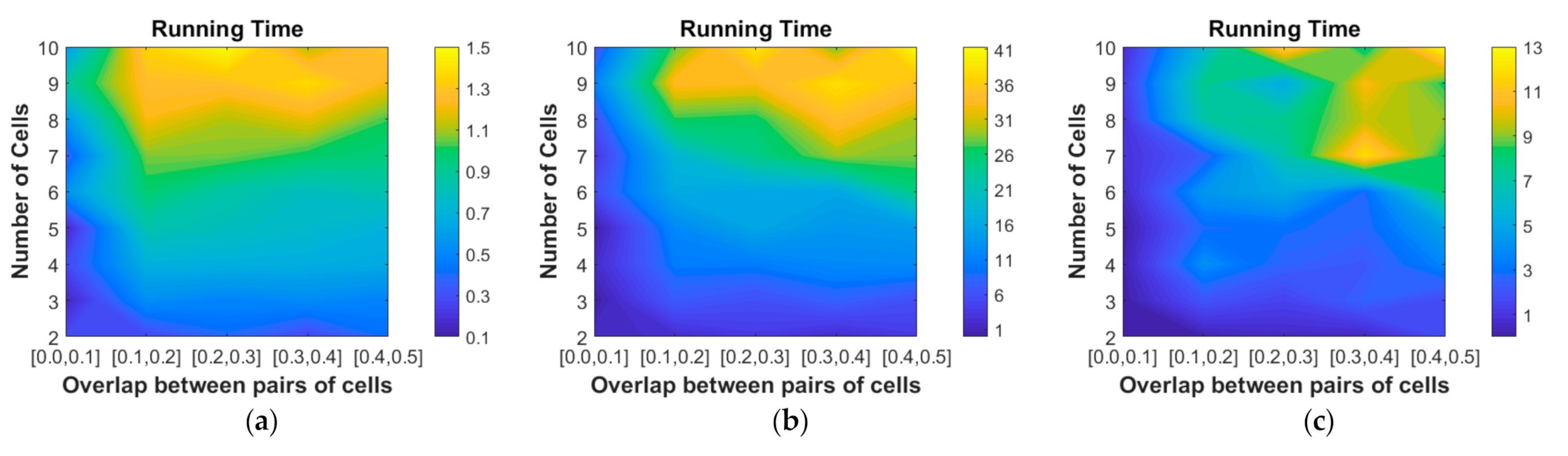
4.2.3. Computational Complexity
4.3. Qualitative Evaluation of Our Segmentation Results
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Forman, D. Global cancer statistics. Cancer J. Clin. 2011, 6, 169–190. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2012, 136, E359–E386. [Google Scholar] [CrossRef]
- Bray, F.; Jianlogong, R.; Masuyer, E.; Ferlay, J. Global estimates of cancer prevalence for 27 sites in the adult population in 2008. Int. J. Cancer 2013, 132, 1133–1145. [Google Scholar] [CrossRef]
- Papanicolaou, G.N. A new procedure for staining vaginal smears. Science 1942, 95, 438–439. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer Prevention & Early Detection Facts & Figures 2015–2016; American Cancer Society: Atlanta, GA, USA, 2015. [Google Scholar]
- Pan, X.P.; Li, L.Q.; Yang, H.H.; Liu, Z.B.; Yang, J.X.; Zhao, L.L.; Fan, Y.X. Accurate segmentation of nuclei in pathological images via sparse reconstruction and deep convolutional networks. Neurocomputing 2017, 229, 88–99. [Google Scholar] [CrossRef]
- Fan, Y.; Bradley, A.P. A method for quantitative analysis of clump thickness in cervical cytology slides. Micron 2016, 80, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Win, K.P.; Kitjaidure, Y.; Hamamoto, K.; Myo Aung, T. Computer-Assisted Screening for Cervical Cancer Using Digital Image Processing of Pap Smear Images. Appl. Sci. 2020, 10, 1800. [Google Scholar] [CrossRef]
- Zhang, L.; Kong, H.; Chin, C.T.; Liu, S.; Chen, Z.; Wang, T.; Chen, S. Segmentation of cytoplasm and nuclei of abnormal cells in cervical cytology using global and local graph cuts. Comput. Med. Imaging Graph. 2014, 38, 369–380. [Google Scholar] [CrossRef]
- Plissiti, M.E.; Nikou, C. A review of automated techniques for cervical cell image analysis and classification. In Biomedical Imaging and Computational Modeling in Biomechanics; Springer-Verlag: Amsterdam, The Netherlands, 2013; Volume 4, pp. 1–18. [Google Scholar]
- Béliz-Osorio, N.; Crespo, J.M.; García-Rojo, A.M.; Azpiazu, J. Cytology Imaging Segmentation Using the Locally Constrained Watershed Transform. In Proceedings of the 10th International Symposium on Mathematical Morphology and Its Applications to Image and Signal Processing (ISMM 2011), Verbania-Intra, Italy, 6–8 July 2011; Springer-Verlag: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bamford, P.; Lovell, B. Unsupervised cell nucleus segmentation with active contours. Signal Process. 1998, 71, 203–213. [Google Scholar] [CrossRef]
- Jung, C.; Kim, C. Segmenting clustered nuclei using H-minima transform-based marker extraction and contour parameterization. IEEE Trans. Biomed. Eng. 2010, 57, 2600–2604. [Google Scholar] [CrossRef]
- Plissiti, M.E.; Nikou, C.; Charchanti, A. Watershed-based segmentation of cell nuclei boundaries in Pap smear images. In Proceedings of the IEEE International Conference on Information Technology & Applications in Biomedicine, Corfu, Greece, 3–5 November 2010. [Google Scholar]
- Jung, C.; Kim, C.; Chae, S.W.; Oh, S. Unsupervised segmentation of overlapped nuclei using Bayesian classification. IEEE Trans. Biomed. Eng. 2010, 57, 2825–2832. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Ping, X.; Ding, Y. Automated cell nucleus segmentation using improved snake. Proc. Int. Conf. Image Process. (ICIP) 2004, 4, 2737–2740. [Google Scholar]
- Plissiti, M.E.; Nikou, C.; Charchanti, A. Automated detection of cell nuclei in pap smear images using morphological reconstruction and clustering. IEEE Trans. Inf. Technol. Biomed. 2011, 15, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Plissiti, M.; Charchanti, A.; Krikoni, O.; Fotiadis, D. Automated segmentation of cell nuclei in Pap smear images. In Proceedings of the IEEE International Special Topic Conference on Information Technology in Biomedicine, Ioannina, Greece, 26–28 October 2006; pp. 26–28. [Google Scholar]
- Lin, C.H.; Chan, Y.K.; Chen, C.C. Detection and segmentation of cervical cell cytoplast and nucleus. Int. J. Imaging Syst. Technol. 2010, 19, 260–270. [Google Scholar] [CrossRef]
- Yang-Mao, S.F.; Chan, Y.K.; Chu, Y.P. Edge Enhancement Nucleus and Cytoplast Contour Detector of Cervical Smear Images. IEEE Trans. Syst. Man Cybern. 2008, 38, 353–366. [Google Scholar] [CrossRef]
- Yang, X.; Gao, X.; Tao, D.; Li, X. Improving level set method for fast auroral oval segmentation. IEEE Trans. Image Process. 2014, 23, 2854–2865. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, L.; Chen, S.; Ni, D.; Lei, B.; Wang, T. Accurate segmentation of cervical cytoplasm and nuclei based on multiscale convolutional network and graph partitioning. IEEE Trans. Biomed. Eng. 2015, 62, 2421–2433. [Google Scholar] [CrossRef]
- Ushizima, D.; Bianchi, A.; Carneiro, C. Segmentation of Subcellular Compartments Combining Superpixel Representation with Voronoi Diagrams; Tech. Rep. LBNL-6892E; Ernest Orlando Lawrence Berkeley Nat. Lab.: Berkeley, CA, USA, 2015. [Google Scholar]
- Zhang, L.; Kong, H.; Chin, C.T.; Liu, S.; Fan, T.; Wang, T.; Chen, S. Automation-assisted cervical cancer screening in manual liquid-based cytology with hematoxylin and eosin staining. Cytometry A 2014, 85, 214–230. [Google Scholar] [CrossRef]
- Ramalho, G.L.; Ferreira, D.S.; Bianchi, A.G.; Carneiro, C.M.; Medeiros, F.N.; Ushizima, D.M. Cell reconstruction under Voronoi and enclosing ellipses from 3D microscopy. In Proceedings of the IEEE International Symposium on Biomedical Imaging (ISBI), Brooklyn Bridge, NY, USA, 16–19 April 2015; pp. 1–2. [Google Scholar]
- Lee, H.; Kim, J. Segmentation of overlapping cervical cells in microscopic images with superpixel partitioning and cell-wise contour refinement. In Proceedings of the 2016 IEEE Conference on Computer Vision and Pattern Recognition Workshops (CVPRW), Las Vegas, NV, USA, 26 June–1 July 2016; pp. 63–69. [Google Scholar]
- Kaur, S.; Sahambi, J. Curvelet initialized level set cell segmentation for touching cells in low contrast images. Comput. Med. Imaging Graph. 2016, 49, 46–57. [Google Scholar] [CrossRef]
- Tareef, A.; Song, Y.; Cai, W.; Feng, D.D.; Chen, M. Automated three-stage nucleus and cytoplasm segmentation of overlapping cells. In Proceedings of the 2014 13th International Conference on Control Automation Robotics & Vision (ICARCV), Singapore, 10–12 December 2014. [Google Scholar]
- Tareef, A.; Song, Y.; Lee, M.Z.; Feng, D.D.; Cai, W. Morphological Filtering and Hierarchical Deformation for Partially Overlapping Cell Segmentation. In Proceedings of the 2015 International Conference on Digital Image Computing: Techniques and Applications (DICTA), Adelaide, Australia, 23–25 November 2015. [Google Scholar]
- Nosrati, M.; Hamarneh, G. A variational approach for overlapping cell segmentation. Proc. IEEE ISBI Overlapping Cerv. Cytol. Image Segm. Chall. 2014, 1, 1–2. [Google Scholar]
- Nosrati, M.; Hamarneh, G. Segmentation of overlapping cervical cells: A variational method with star-shape prior. Proc. ISBI 2015, 186–189. [Google Scholar] [CrossRef]
- Lu, Z.; Carneiro, G.; Bradley, A.P. Automated nucleus and cytoplasm segmentation of overlapping cervical cells. In Medical Image Computing and Computer-Assisted Intervention; Springer: Berlin, Germany, 2013; pp. 452–460. [Google Scholar]
- Lu, Z.; Carneiro, G.; Bradley, A. An improved joint optimization of multiple level set functions for the segmentation of overlapping cervical cells. IEEE Trans. Image Process. 2015, 24, 261–272. [Google Scholar]
- Osher, S.; Sethian, J.A. Fronts propagating with curvature dependent speed: Algorithms based on Hamilton–Jacobi formulations. J. Comput. Phys. 1988, 79, 12–49. [Google Scholar] [CrossRef]
- Kass, M.; Witkin, A.; Terzopoulos, D. Snakes: Active contour models. Int. J. Comput. Vis. 1988, 1, 321–331. [Google Scholar] [CrossRef]
- Caselles, V.; Catt´e, F.; Coll, T.; Dibos, F. A Geometric Model for Active Contours in Image Processing. Numer. Math. 1993, 66, 1–31. [Google Scholar]
- Malladi, R.; Sethian, J.A.; Vemuri, B.C. Shape Modeling with Front Propagation: A Level Set Approach. IEEE Trans. Pami. 1995, 17, 158–175. [Google Scholar] [CrossRef]
- Mumford, D.; Shah, J. Optimal approximations by piecewise smooth functions and associated variational problems. Commun. Pure Appl. Math. 1989, 42, 577–685. [Google Scholar] [CrossRef]
- Zhao, H.K.; Chan, T.; Merriman, B.; Osher, S. A Variational Level Set Approach to Multiphase Motion. J. Comput. Phys. 1996, 127, 179–195. [Google Scholar] [CrossRef]
- Chan, T.; Vese, L. An Active Contour Model without Edges. In International Conference on Scale-Space Theories in Computer Vision; Springer-Verlag: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Caselles, V.; Kimmel, R.; Sapiro, G. Geodesic active contours. Int. J. Comput. Vis. 1997, 22, 61–79. [Google Scholar] [CrossRef]
- Chen, Y.; Thiruvenkadam, H.; Tagare, H.; Huang, F.; Wilson, D. On the Incorporation of Shape Priors int Geometric Active Contours. In Proceedings of the IEEE Workshop in Variational and Level Set Methods, Vancouver, BC, Canada, 1 February 2001; pp. 145–152. [Google Scholar]
- Li, C.M.; Xu, C.Y.; Gui, C.F.; Martin, D.F. Distance regularized level set evolution and its application to image segmentation. IEEE Trans. Image Process. 2010, 19, 3243–3254. [Google Scholar]
- Canny, J. A Computational Approach to Edge Detection. IEEE Trans. Pattern Anal. Mach. Intell. PAMI 1986, 679–698. [Google Scholar] [CrossRef]
- Smereka, P. Spiral Crystal Growth. Phys. D Nonlinear Phenom. 2000, 138, 282–301. [Google Scholar] [CrossRef]
- Burchard, P.; Cheng, L.T.; Merriman, B.; Osher, S. Motion of curves in three spatial dimensions using a level set approach. J. Comput. Phys. 2001, 170, 720–741. [Google Scholar] [CrossRef]
- Gao, X.; Wang, B.; Tao, D.; Li, X. A relay level set method for automatic image segmentation. IEEE Trans. Syst. Man Cybern. 2011, 41, 518. [Google Scholar]
- Wang, B.; Gao, X.; Tao, D.; Li, X. A Unified Tensor Level Set for Image Segmentation. IEEE Trans. Syst. Man Cybern. 2010, 40, 857–867. [Google Scholar] [CrossRef]
- Yang, X.; Gao, X.; Li, J.; Han, B. A shape-initialized and intensity-adaptive level set method for auroral oval segmentation. Inf. Sci. 2014, 277, 794–807. [Google Scholar] [CrossRef]
- Tareef, A.; Song, Y.; Huang, H.; Wang, Y.; Feng, D.; Chen, M.; Cai, W. Optimizing the cervix cytological examination based on deep learning and dynamic shape modeling. Neurocomputing 2017, 248, 28–40. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, H.; Wang, P.; Dong, D. Segmentation of Overlapping Cervical Smear Cells Based on U-Net and Improved Level Set. In Proceedings of the 2019 IEEE International Conference on Systems, Man and Cybernetics (SMC), Bari, Italy, 6–9 October 2019. [Google Scholar]
- Tareef, A.; Song, Y.; Cai, W.; Huang, H.; Chen, M. Learning Shape-Driven Segmentation Based on Neural Network and Sparse Reconstruction toward Automated Cell Analysis of Cervical Smears. Int. Conf. Neural Inf. Process. 2015, 1, 390–400. [Google Scholar]
- Tareef, A.; Song, Y.; Cai, W.; Huang, H.; Chang, H.; Wang, Y.; Fulham, M.; Feng, D.; Chen, M. Automatic segmentation of overlapping cervical smear cells based on local distinctive features and guided shape deformation. Neurocomputing 2017, 221, 94–107. [Google Scholar] [CrossRef]
- Song, Y.Y.; Tan, E.; Jiang, X.D.; Cheng, J.Z.; Ni, D.; Chen, S.P.; Lei, B.; Wang, T. Accurate Cervical Cell Segmentation from Overlapping Clumps in Pap Smear Images. IEEE Trans. Med. Imaging 2017, 36, 288–300. [Google Scholar] [CrossRef]
- Litjens, G.; Kooi, T.; Bejnordi, B.E.; Setio, A.A.A.; Ciompi, F.; Ghafoorian, M. A survey on deep learning in medical image analysis. Med. Image Anal. 2017, 42, 60–88. [Google Scholar] [CrossRef]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net. Convolutional Networks for Biomedical Image Segmentation. In Proceedings of the International Conference on Medical Image Computing and Computer-Assisted Intervention, Munich, Germany, 5–9 October 2015. [Google Scholar]
- He, K.; Gkioxari, G.; Dollar, P.; Girshick, R. Mask R-CNN. IEEE Trans. Pattern Anal. Mach. Intell. 2018, 42, 386–397. [Google Scholar] [CrossRef]
- Kirillov, A.; Girshick, R.B.; He, K.; Dollár, P. Panoptic Feature Pyramid Networks. arXiv 2018, arXiv:1901.02446. [Google Scholar]
- Wan, T.; Xu, S.; Sang, C.; Jin, Y.; Qin, Z. Accurate segmentation of overlapping cells in cervical cytology with deep convolutional neural networks. Neurocomputing 2019, 365, 157–170. [Google Scholar] [CrossRef]
- Li, K.; Lu, Z.; Liu, W.; Yin, J. Cytoplasm and nucleus segmentation in cervical smear images using radiating GVF snake. Pattern Recognit. 2012, 45, 1255–1264. [Google Scholar] [CrossRef]
- Kale, A.; Aksoy, S. Segmentation of cervical cell images. In Proceedings of the 2010 20th International Conference on Pattern Recognition, Istanbul, Turkey, 23–26 August 2010; pp. 2399–2402. [Google Scholar]
- Maurer, C.R.; Qi, R.; Raghavan, V. A linear time algorithm for computing exact euclidean distance transforms of binary images in arbitrary dimensions. IEEE Trans. Pattern Anal. Mach. Intell. 2003, 25, 265–270. [Google Scholar] [CrossRef]
- Osher, S.; Fedkiw, R. Level Set Methods and Dynamic Implicit Surfaces; Springer: New York, NY, USA, 2003. [Google Scholar]
- Meyer, F. Topographic distance and watershed lines. Signal Process. 1994, 38, 113–125. [Google Scholar] [CrossRef]
- Vedaldi, A.; Soatto, S. Quick shift and kernel methods for mode seeking. Comput. Vis. 2008, 705–718. [Google Scholar] [CrossRef]
- Dillencourt, M.B.; Samet, H.; Tamminen, M. A general approach to connected-component labeling for arbitrary image representations. J. ACM 1992, 39, 253–280. [Google Scholar] [CrossRef]
- Matas, J.; Chum, O.; Urban, M.; Pajdla, T. Robust wide baseline stereo from maximally stable extremal regions. In Proceedings of the British Machine Vision Conference 2002, Cardiff, UK, 2–5 September 2002; pp. 384–396. [Google Scholar]
- Osher, S. A Level Set Formulation for the Solution of the Dirichlet Problem for Hamilton–Jacobi Equations. SIAM J. Math. Anal. 1993, 24, 1145–1152. [Google Scholar] [CrossRef]
- Tsitsiklis, J. Efficient Algorithms for Globally Optimal Trajectories. IEEE Trans. Autom. Control 1995, 40, 1528–1538. [Google Scholar] [CrossRef]
- Guangqi, L. Mathmatical-Toolbox-for-DCTLSM. Available online: https://github.com/liuchee/Mathmatical-Toolbox-for-DCTLSM (accessed on 1 September 2020).
- Overlapping Cervical Cytology Image Segmentation Challenge-ISBI. Available online: https://cs.adelaide.edu.au/~carneiro/isbi14_challenge/index.html (accessed on 6 September 2020).
- Bradley, A.; Bamford, P. A one-pass extended depth of field algorithm based on the over-complete discrete wavelet transform. Image Vis. Comput. N. Z. (IVCNZ) 2004, 1, 279–284. [Google Scholar]
- Lu, Z.; Carneiro, G.; Bradley, A.; Ushizima, D.; Nosrati, M.; Bianchi, A.G.C.; Carneiro, C.M.; Hamarneh, G. Evaluation of Three Algorithms for the Segmentation of Overlapping Cervical Cells. IEEE J. Biomed. Health Inform. 2017, 21, 441–450. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TH | 2 Cells | 3 Cells | 4 Cells | 5 Cells | 6 Cells | 7 Cells | 8 Cells | 9 Cells | 10 Cells |
|---|---|---|---|---|---|---|---|---|---|
| ISBI-14 Train-45 Dataset (MSTF) | |||||||||
| 2 | ΔFPP = −0.38 ΔFNP = +1.19 | ΔFPP = −0.05 ΔFNP = +0.67 | ΔFPP = −0.09 ΔFNP = +0.42 | ΔFPP = −0.21 ΔFNP = +0.66 | ΔFPP = −0.06 ΔFNP = +0.46 | ΔFPP = −0.15 ΔFNP = +0.39 | ΔFPP = −0.21 ΔFNP = +0.75 | ΔFPP = −0.19 ΔFNP = +0.61 | ΔFPP = −0.14 ΔFNP = +0.98 |
| 5 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.06 ΔFNP = +0.70 | ΔFPP = −0.09 ΔFNP = +0.43 | ΔFPP = −0.26 ΔFNP = +0.70 | ΔFPP = −0.06 ΔFNP = +0.47 | ΔFPP = −0.17 ΔFNP = +0.43 | ΔFPP = −0.24 ΔFNP = +0.78 | ΔFPP = −0.24 ΔFNP = +0.63 | ΔFPP = −0.14 ΔFNP = +1.02 |
| 10 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.06 ΔFNP = +0.77 | ΔFPP = −0.13 ΔFNP = +0.47 | ΔFPP = −0.36 ΔFNP = +0.84 | ΔFPP = −0.08 ΔFNP = +0.56 | ΔFPP = −0.21 ΔFNP = +0.48 | ΔFPP = −0.29 ΔFNP = +0.86 | ΔFPP = −0.28 ΔFNP = +0.70 | ΔFPP = −0.16 ΔFNP = +1.10 |
| 15 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.07 ΔFNP = +0.90 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.39 ΔFNP = +1.00 | ΔFPP = −0.11 ΔFNP = +0.74 | ΔFPP = −0.27 ΔFNP = +0.56 | ΔFPP = −0.40 ΔFNP = +0.97 | ΔFPP = −0.34 ΔFNP = +0.82 | ΔFPP = −0.19 ΔFNP = +1.34 |
| 20 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.08 ΔFNP = +1.11 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.39 ΔFNP = +1.11 | ΔFPP = −0.19 ΔFNP = +0.91 | ΔFPP = −0.33 ΔFNP = +0.64 | ΔFPP = −0.47 ΔFNP = +1.09 | ΔFPP = −0.38 ΔFNP = +0.95 | ΔFPP = −0.27 ΔFNP = +1.73 |
| 25 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.09 ΔFNP = +1.32 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.28 ΔFNP = +1.11 | ΔFPP = −0.56 ΔFNP = +0.72 | ΔFPP = −0.53 ΔFNP = +1.21 | ΔFPP = −0.43 ΔFNP = +1.14 | ΔFPP = −0.39 ΔFNP = +2.13 |
| 30 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.14 ΔFNP = +1.86 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.35 ΔFNP = +1.27 | ΔFPP = −0.56 ΔFNP = +0.76 | ΔFPP = −0.57 ΔFNP = +1.30 | ΔFPP = −0.44 ΔFNP = +1.32 | ΔFPP = −0.72 ΔFNP = +2.36 |
| 35 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.22 ΔFNP = +2.49 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.39 ΔFNP = +1.43 | ΔFPP = −0.56 ΔFNP = +0.85 | ΔFPP = −0.57 ΔFNP = +1.37 | ΔFPP = −0.44 ΔFNP = +1.32 | ΔFPP = −0.72 ΔFNP = +2.36 |
| 50 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.70 ΔFNP = +2.99 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.56 ΔFNP = +1.65 | ΔFPP = −0.57 ΔFNP = +1.01 | ΔFPP = −0.57 ΔFNP = +1.37 | ΔFPP = −0.44 ΔFNP = +1.32 | ΔFPP = −0.72 ΔFNP = +2.36 |
| 70 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.70 ΔFNP = +2.99 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.56 ΔFNP = +1.65 | ΔFPP = −0.57 ΔFNP = +1.01 | ΔFPP = −0.57 ΔFNP = +1.37 | ΔFPP = −0.44 ΔFNP = +1.32 | ΔFPP = −0.72 ΔFNP = +2.36 |
| 100 | ΔFPP = −0.84 ΔFNP = +1.22 | ΔFPP = −0.70 ΔFNP = +2.99 | ΔFPP = −0.70 ΔFNP = +0.57 | ΔFPP = −0.40 ΔFNP = +1.19 | ΔFPP = −0.56 ΔFNP = +1.65 | ΔFPP = −0.57 ΔFNP = +1.01 | ΔFPP = −0.57 ΔFNP = +1.37 | ΔFPP = −0.44 ΔFNP = +1.32 | ΔFPP = −0.72 ΔFNP = +2.36 |
| ISBI-14 Train-45 Dataset (DCTLSM) | |||||||||
| 15 | ΔFPP = −0.54 ΔFNP = −0.08 | ΔFPP = −0.06 ΔFNP = −0.04 | ΔFPP = −0.67 ΔFNP = −0.03 | ΔFPP = −0.30 ΔFNP = −0.02 | ΔFPP = +0.00 ΔFNP = −0.02 | ΔFPP = −0.14 ΔFNP = +0.02 | ΔFPP = −0.18 ΔFNP = −0.00 | ΔFPP = −0.23 ΔFNP = −0.01 | ΔFPP = −0.10 ΔFNP = +0.05 |
| TH | 2 Cells | 3 Cells | 4 Cells | 5 Cells | 6 Cells | 7 Cells | 8 Cells | 9 Cells | 10 Cells |
|---|---|---|---|---|---|---|---|---|---|
| ISBI-14 Test-90 Dataset (MSTF) | |||||||||
| 15 | ΔFPP = −0.74 ΔFNP = +0.83 | ΔFPP = 0.00 ΔFNP = 0.00 | ΔFPP = −0.36 ΔFNP = +0.97 | ΔFPP = 0.00 ΔFNP = 0.00 | ΔFPP = −0.25 ΔFNP = +0.64 | ΔFPP = −0.31 ΔFNP = +0.72 | ΔFPP = −0.04 ΔFNP = +0.90 | ΔFPP = −0.37 ΔFNP = +0.91 | ΔFPP = −0.14 ΔFNP = +0.79 |
| ISBI-14 Test-90 Dataset (DCTLSM) | |||||||||
| 15 | ΔFPP = −0.74 ΔFNP = −0.02 | ΔFPP = 0.00 ΔFNP = 0.00 | ΔFPP = −0.36 ΔFNP = +0.02 | ΔFPP = 0.00 ΔFNP = 0.00 | ΔFPP = −0.23 ΔFNP = −0.06 | ΔFPP = −0.21 ΔFNP = −0.02 | ΔFPP = −0.01 ΔFNP = +0.04 | ΔFPP = −0.32 ΔFNP = −0.02 | ΔFPP = −0.10 ΔFNP = −0.06 |
| Methods | DC > 0.6 | DC > 0.7 | DC > 0.8 | DC > 0.9 |
|---|---|---|---|---|
| ISBI-14 Train-45 Dataset | ||||
| Lu [33] | DC = 0.905(0.078), FNO = 0.126(0.181) TPP = 0.917(0.087), FPP = 0.006(0.009) | DC = 0.912(0.066), FNO = 0.148(0.206) TPP = 0.920(0.080), FPP = 0.005(0.007) | DC = 0.924(0.049), FNO = 0.211(0.241) TPP = 0.927(0.066), FPP = 0.004(0.005) | DC = 0.951(0.027), FNO = 0.441(0.309) TPP = 0.943(0.043), FPP = 0.002(0.003) |
| Ours | DC = 0.917(0.068), FNO = 0.078(0.098) TPP = 0.930(0.069), FPP = 0.005(0.007) | DC = 0.923(0.059), FNO = 0.104(0.126) TPP = 0.931(0.065), FPP = 0.004(0.006) | DC = 0.935(0.045), FNO = 0.200(0.205) TPP = 0.936(0.056), FPP = 0.004(0.005) | DC = 0.960(0.023), FNO = 0.456(0.317) TPP = 0.947(0.039), FPP = 0.001(0.002) |
| ISBI-14 Test-90 Dataset | ||||
| Lu [33] | DC = 0.871(0.102), FNO = 0.254(0.268) TPP = 0.892(0.106), FPP = 0.005(0.007) | DC = 0.891(0.082), FNO = 0.317(0.284) TPP = 0.895(0.102), FPP = 0.003(0.006) | DC = 0.920(0.058), FNO = 0.439(0.304) TPP = 0.913(0.082), FPP = 0.002(0.003) | DC = 0.956(0.029), FNO = 0.630(0.301) TPP = 0.947(0.045), FPP = 0.001(0.002) |
| Ours | DC = 0.882(0.095), FNO = 0.178(0.177) TPP = 0.906(0.091), FPP = 0.005(0.007) | DC = 0.904(0.073), FNO = 0.281(0.226) TPP = 0.911(0.085), FPP = 0.003(0.005) | DC = 0.931(0.051), FNO = 0.450(0.269) TPP = 0.925(0.071), FPP = 0.002(0.004) | DC = 0.961(0.027), FNO = 0.663(0.292) TPP = 0.949(0.044), FPP = 0.001(0.001) |
| Methods | FNO | TPP | FPP | DC |
|---|---|---|---|---|
| ISBI-14 Train-45 Dataset | ||||
| Ushizima [23] | 0.267(0.278) | 0.841(0.130) | 0.002(0.003) | 0.872(0.082) |
| Nosrati [30] | 0.111(0.166) | 0.875(0.086) | 0.004(0.004) | 0.871(0.075) |
| Tareef [28] | 0.296(0.277) | 0.948(0.059) | 0.005(0.007) | 0.914(0.075) |
| Lu [33] | 0.148(0.206) | 0.920(0.080) | 0.005(0.007) | 0.912(0.066) |
| Lee [26] | 0.137(0.194) | 0.882(0.097) | 0.002(0.003) | 0.897(0.075) |
| Ours | 0.104(0.126) | 0.931(0.065) | 0.004(0.006) | 0.923(0.059) |
| Methods | FNO | TPP | FPP | DC |
|---|---|---|---|---|
| ISBI-14 Test-90 Dataset | ||||
| Ushizima [23] | 0.174(0.210) | 0.826(0.130) | 0.001(0.002) | 0.867(0.083) |
| Nosrati [30] | 0.140(0.170) | 0.900(0.090) | 0.005(0.004) | 0.870(0.080) |
| Nosrati [31] | 0.110(0.170) | 0.930(0.090) | 0.005(0.004) | 0.880(0.080) |
| Lu [33] | 0.317(0.284) | 0.895(0.102) | 0.003(0.006) | 0.891(0.082) |
| Tareef [52] | 0.163(0.223) | 0.939(0.064) | 0.005(0.005) | 0.888(0.076) |
| Tareef [53] | 0.274(0.277) | 0.907(0.088) | 0.004(0.005) | 0.889(0.073) |
| Tareef [50] | 0.222(0.240) | 0.945(0.071) | 0.005(0.005) | 0.897(0.077) |
| Huang [51] | 0.100(0.130) | 0.940(0.090) | 0.004(0.003) | 0.890(0.070) |
| Ours | 0.281(0.226) | 0.911(0.085) | 0.003(0.005) | 0.904(0.073) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Ding, Q.; Luo, H.; Ju, M.; Jin, T.; He, M.; Dong, G. A Novel Evolution Strategy of Level Set Method for the Segmentation of Overlapping Cervical Cells. Appl. Sci. 2021, 11, 443. https://doi.org/10.3390/app11010443
Liu G, Ding Q, Luo H, Ju M, Jin T, He M, Dong G. A Novel Evolution Strategy of Level Set Method for the Segmentation of Overlapping Cervical Cells. Applied Sciences. 2021; 11(1):443. https://doi.org/10.3390/app11010443
Chicago/Turabian StyleLiu, Guangqi, Qinghai Ding, Haibo Luo, Moran Ju, Tianming Jin, Miao He, and Gang Dong. 2021. "A Novel Evolution Strategy of Level Set Method for the Segmentation of Overlapping Cervical Cells" Applied Sciences 11, no. 1: 443. https://doi.org/10.3390/app11010443
APA StyleLiu, G., Ding, Q., Luo, H., Ju, M., Jin, T., He, M., & Dong, G. (2021). A Novel Evolution Strategy of Level Set Method for the Segmentation of Overlapping Cervical Cells. Applied Sciences, 11(1), 443. https://doi.org/10.3390/app11010443