During the first phase of 19 days of this trial, conducted under the HRT of 6.7 d and OLR of 4.5 g COD/L·d, biogas production ranged between 0.4 and 1.2 L/L·d, mainly composed of methane in concentrations of 63–78% (
Figure 2). A biogas production rate of 0.996 L/L·d and 0.928 L/L·d, containing 65% methane, was registered after the digestion of pre-treated manure (140 °C, 40 min) and without pre-treatment, respectively [
12]. The values for biogas and methane content are of the same order of magnitude than those obtained in this work (1.1–1.2 L/L·d), however they were performed under identical OLR (2.5 g VS/L·d versus 2.2 g VS/ L·d) but advantageous operating conditions. They resulted from a higher digestion process temperature and longer HRT than the conditions in this work (55 °C, 18 d HRT versus 37 °C, 6.7 d HRT) [
12]. Thermophilic conditions of temperature, such as 55 °C, promote higher biogas productions than the mesophilic ones (37 °C) as shown by Kasinski [
53]. On the other hand, the increase in HRT allows for a better conversion of organic matter and, consequently, greater volumes of biogas will be expected as concluded by Shi et al. [
54]. Bearing these considerations in mind, the data obtained in the present work are comparatively of relevant interest because they indicate that it is possible to obtain a similar production in biogas (energy) and treat a higher volume of effluent in the same period and, at lower process temperature, conditions that will certainly provide an energy balance more attractive for the operating unit. Considering the co-digestion of Pennisetum hybrid and pig manure (50:50) under a mesophilic range of temperature (35 ± 1 °C) [
55], the results obtained are comparable to those of this work, although with a lower methane content: biogas production of 1.16 and 2.07 L/L·d with 77.8 and 79.5% CH
4 (OLR 2.22 and 4.96 g VS/L·d) versus 0.87 and 1.92 L/L·d with 56.7 and 60.3% CH
4 (OLR 2.0 and 5.0 g VS/ L·d [
55]) were registered, respectively.
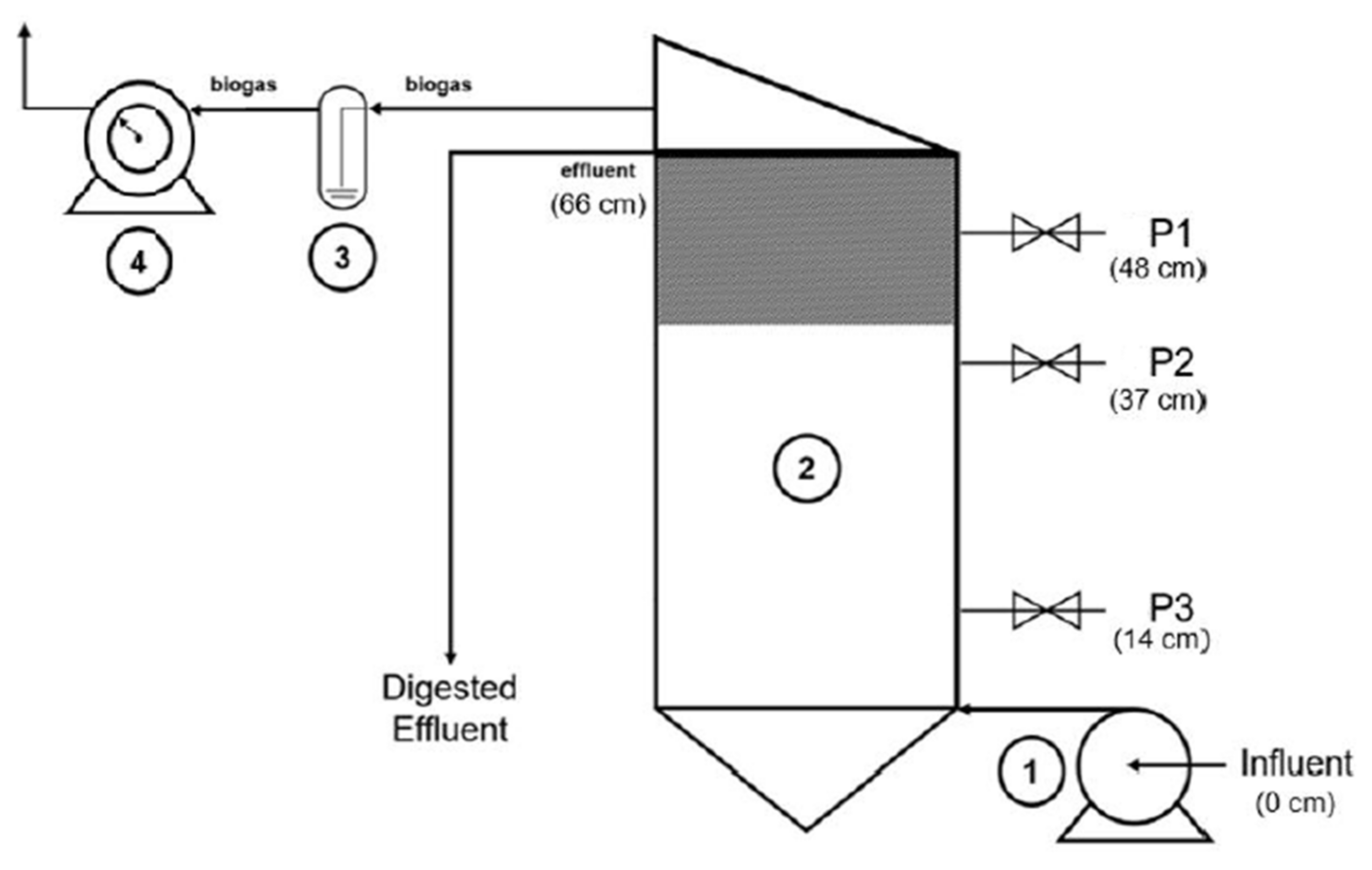
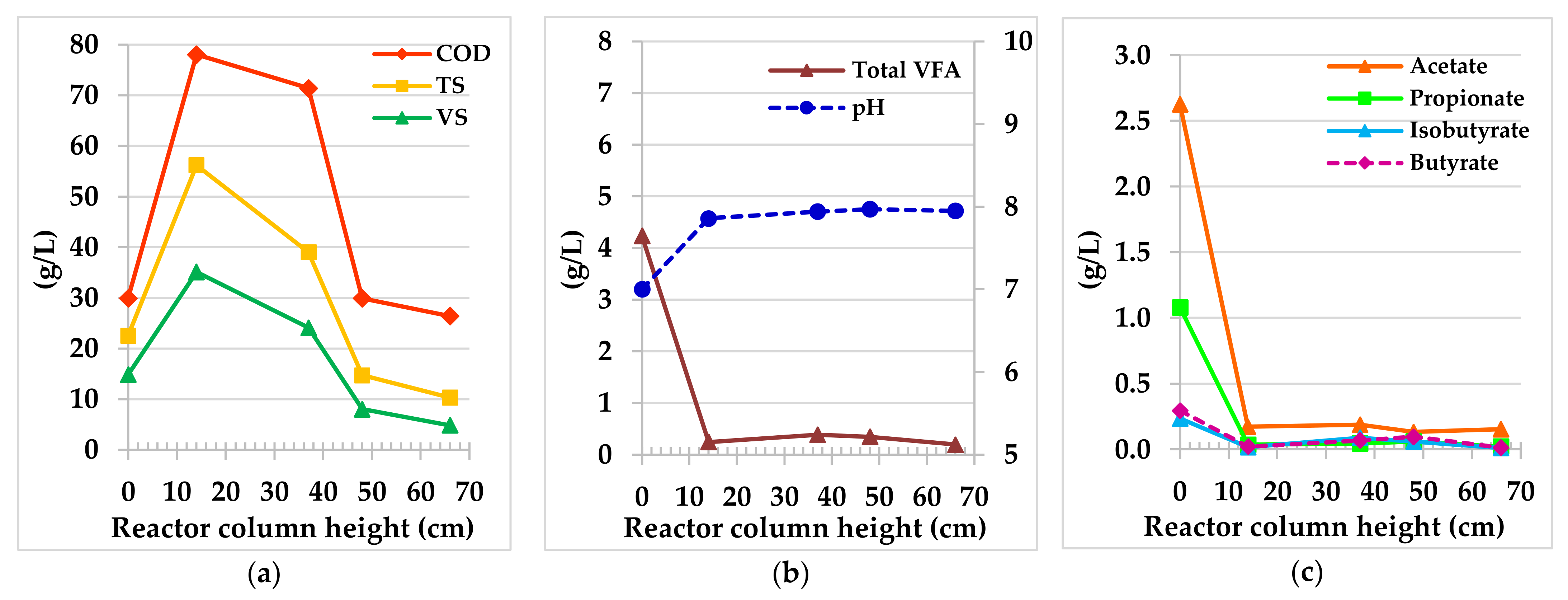
Gas production is a result of the organic matter conversion, and the partial removal of influent contents is consequently achieved. According to the parameters in
Table 3, the decrease in substrate concentration, shown in each process stage, indicates that the mixture digestion took place properly under the tested experimental conditions, as expected. However, regarding the data obtained, COD removal decreased from 52 to 12% with the reduction in the HRT from 6.7 d to 3.0 d, contradicting the increase in biogas production (
Figure 2). A possible explanation for this would be the presence of additional material in the effluent, such as suspended biomass that reached the top of the hybrid, and was counted as COD content, when the sample was taken for analysis. Regarding solids, the greatest removal amounts of 54% in TS, and 68% in VS, were obtained precisely at HRT 3 d, suggesting that microbial populations were able to convert the majority of the organic matter into biogas. Values of this order of magnitude (55–65% TS and 59–68% VS) were also reported by Demirer and Chen [
58] when digesting dairy manure in an anaerobic hybrid reactor, equipped with a floating support for biomass immobilization, yet these values were obtained at a higher HRT of 15 d. In another experiment, Lo et al. [
57] achieved COD removal of 57% using a hybrid UASB reactor to test the swine manure treatment.
Regarding nitrogen compounds, despite the similarity between the input and output of TKN values during the initial stage (HRT 6.7 d), the decrease in concentration (21%) achieved after the regimen at HRT 3 d suggests protein degradation by microbial populations, within the hybrid anaerobic reactor, under this last operational condition (
Table 3). The ammonium nitrogen data did not provide much additional information due to the small difference registered between the substrate and digestate concentrations. Considering that the proportion of ionized and non-ionized forms (the toxic one) in the medium is pH-dependent and that values in the basic pH range (7.9–8.1) were registered in this work, some toxicity from ammonia could be considered. Nevertheless, from the available data, the initial amount of 800 mg NH
4+-N/L is low enough to have any inhibitory effect on the process [
11]. As expected, ammonium nitrogen concentration increased in the digestate during the first stage, although not in a very marked way. In the next phase (HRT 3 d), a low removal of ammonium was registered, indicating that both the ammonium contained in the substrate and the ammonium obtained from the conversion of compounds were used in the process. In fact, the greatest VFA removal (95%,
Table 3) was registered in the HRT 3 d operational condition, signifying the presence of an adequate buffer capacity in the medium.
3.2.2. Molecular Characterization of Microbial Communities in the Hybrid Anaerobic Reactor
To compare distribution of microbial populations during the experiment, a microbial analysis of effluents and samples, collected at the beginning of HRT 6.7 d IN and at the end of HRT 3 d OUT, was conducted. The sample collected at the end of the first phase (HRT 6.7 d OUT) of anaerobic digestion failed library preparation and was excluded from the sequencing run.
After NGS, the samples generated were between the minimum of 609,126 and maximum of 682,804 raw sequence reads, which corresponded to samples HRT 6.7 d (IN) and HRT 3 d (OUT), respectively. A total of 777,827 sequences (775,651 bacterial, 2123 archaeal, and 53 unassigned) were retrieved and analyzed (
Table 5). Libraries were composed of a total 2634 observed OTUs, that were detected in each sample.
The highest Shannon–Wiener indices (4.95 and 5.28 for bacteria and archaea, respectively,
Table 5), suggested that microbial communities were the most diverse at the beginning of experiment (HRT 6.7 d IN). The further decrease observed in Shannon–Wiener index values, until the end of the anaerobic digestion, confirms the occurrence of the microbial acclimation (bacteria and archaea) to the operating hydraulic condition, that is, shortening HRT to 3 d with concomitant increase in OLR up to 10 g COD/L·d. The decrease in microbial diversity means that the microbiota suffers a specialization shift in the microbial structure, consistent with the good performance of the hybrid anaerobic reactor.
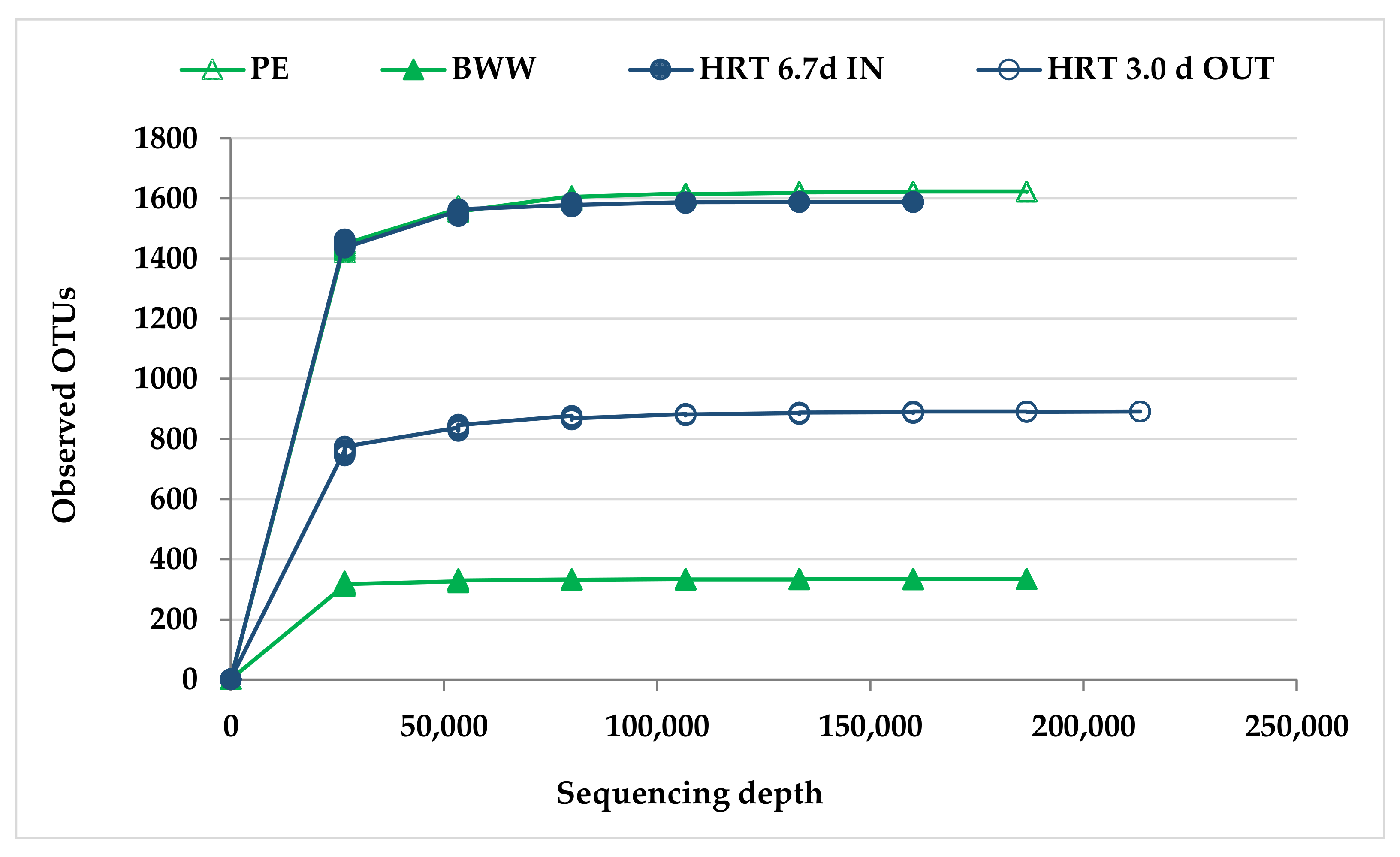
In the case of microbial diversity analysis, it is necessary to know whether the number of reads of the sequencing reaches a reasonable amount so that more sequencing does not significantly increase species diversity.
Figure 4 indicates that, with 97% similarity levels, the rarefaction curve was asymptotic; that is, a flat portion of observed OTUs was attained, representing that the sequencing was sufficient for all samples, and the dataset of sequences had thoroughly sampled diversity in this analysis and sufficient sequence depth was achieved.
Relative abundance of bacterial and archaeal groups was determined in terms of the percentage of total number of sequences in each sample. As predictable, bacteria predominated the microbial populations in all samples, accounting for more than 99.7% of relative abundance at domain level (
Table 6). The discrepancy found between Bacteria and Archaea is normal, real and has already been reported by other authors in anaerobic digestion processes [
59,
60,
61]. Most of the microorganisms presented in the substrate (HRT 6.7 d IN) belonged to the Bacteria domain, with 0.7% represented by the Archaea domain (
Table 6).
During the second phase (HRT 3 d) of the anaerobic digestion process in the hybrid anaerobic reactor, archaea populations drastically decreased yet, at a later phase, HRT 1 d, they recovered to the initial values of 0.7% (data not shown), confirming the maintenance quality of the biogas, at methane levels of around 80%.
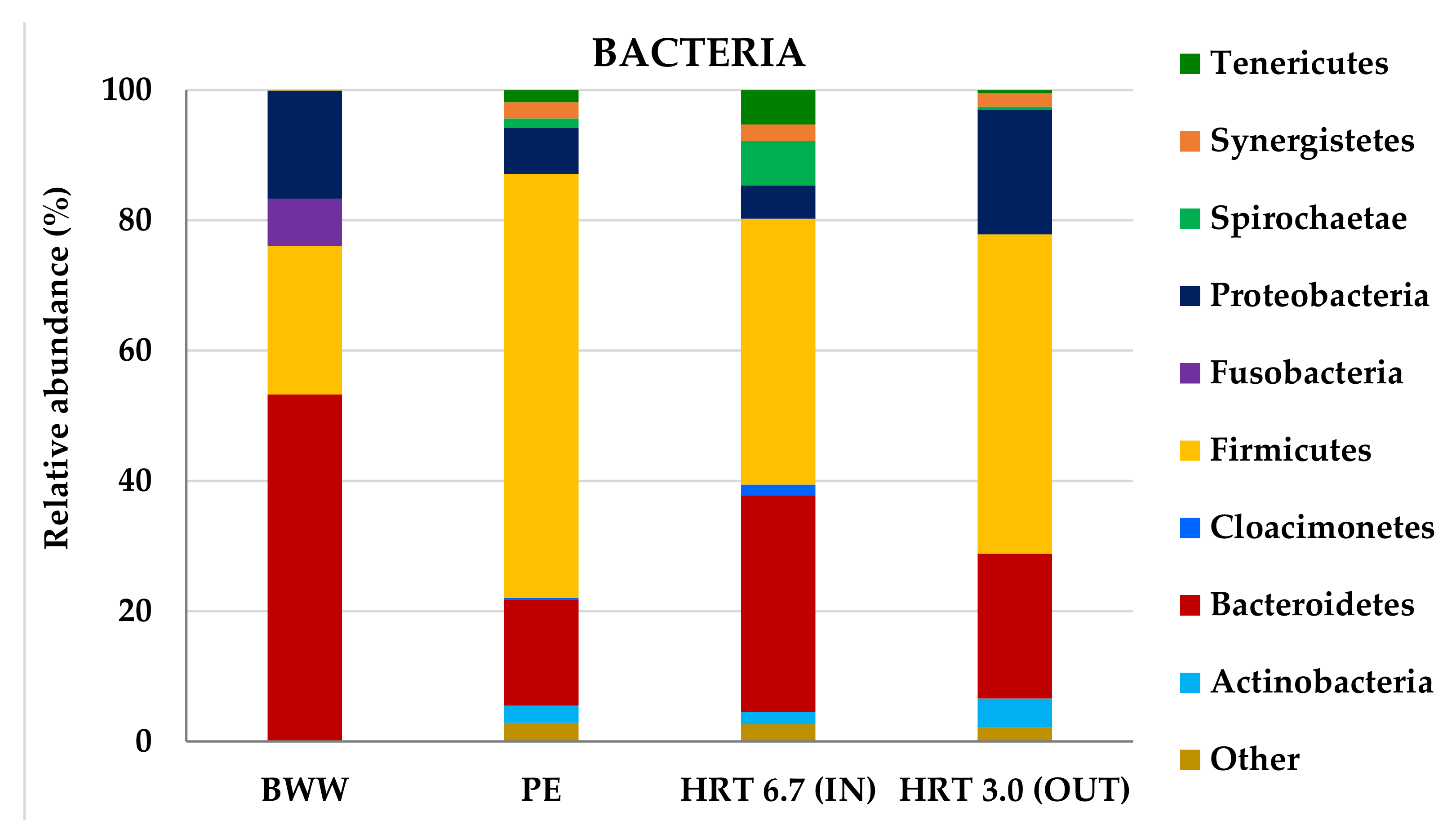
The composition of the phyla with relative abundances of bacterial communities greater than 1% in at least one sample is illustrated in
Figure 5. There were detected nine major phyla including Actinobacteria, Bacteroidetes, Cloacimonetes, Firmicutes, Fusobacteria, Proteobacteria, Spirochaetae, Synergistetes, and Tenericutes, accounting for 97.1% (PE)–99.9% (BWW) of the entire bacterial community. Members of phyla Bacteroidetes, Cloacimonetes, Firmicutes, Proteobacteria, Spirochaetae, and Synergistetes are known to be part of a structural community that efficiently converts the biomass, chemically complex feedstocks, into high-yield methane containing biogas [
62]. Proteobacteria, Firmicutes, and Bacteroidetes are likely abundant in all anaerobic digestion systems and have been referred to by several authors (e.g., [
59,
60,
61,
63]). Moreover, these phyla contain several species whose metabolism can contribute to one or more of the phases of the general anaerobic digestion process.
A diverse bacterial population was found in both effluents (
Figure 5), being dominated mainly by Bacteroidetes (53%) on BWW and by Firmicutes (65%) on PE. These bacterial populations remain predominant throughout the experiment, and Firmicutes increases from 41% to 49% on the last phase (HRT 3 d OUT) becoming the predominant phylum in this sample. The observed decrease in Bacteroidetes during anaerobic digestion further implies that it may have competed with Firmicutes over the same resources on the reactor, the latter prevailing. Buhlmann et al. [
64] already mentioned that Bacteroidetes and Firmicutes have been identified as the main bacterial phyla present within anaerobic digesters. Most of members of the phyla Bacteroidetes are known for their ability either to convert easily degradable amino acids, sugars, and alcohols into VFA, which emphasizes their potential crucial role in acido- and acetogenesis. On the other hand, the Bacteroidetes activity can be self-inhibited by the accumulation of acids [
65]. Some Firmicutes are known to be syntrophic bacteria capable of degrading VFAs, such as butyrate and its analogues [
63]. In addition, most of strains in the phylum Firmicutes might hydrolyze cellulose, glucose, starch, and proteins into small molecular organic acids such as acetate, highly contributing to improvement of the methane production yield [
66,
67]. Microbial community profiles are demonstrated with more detail at genus level in
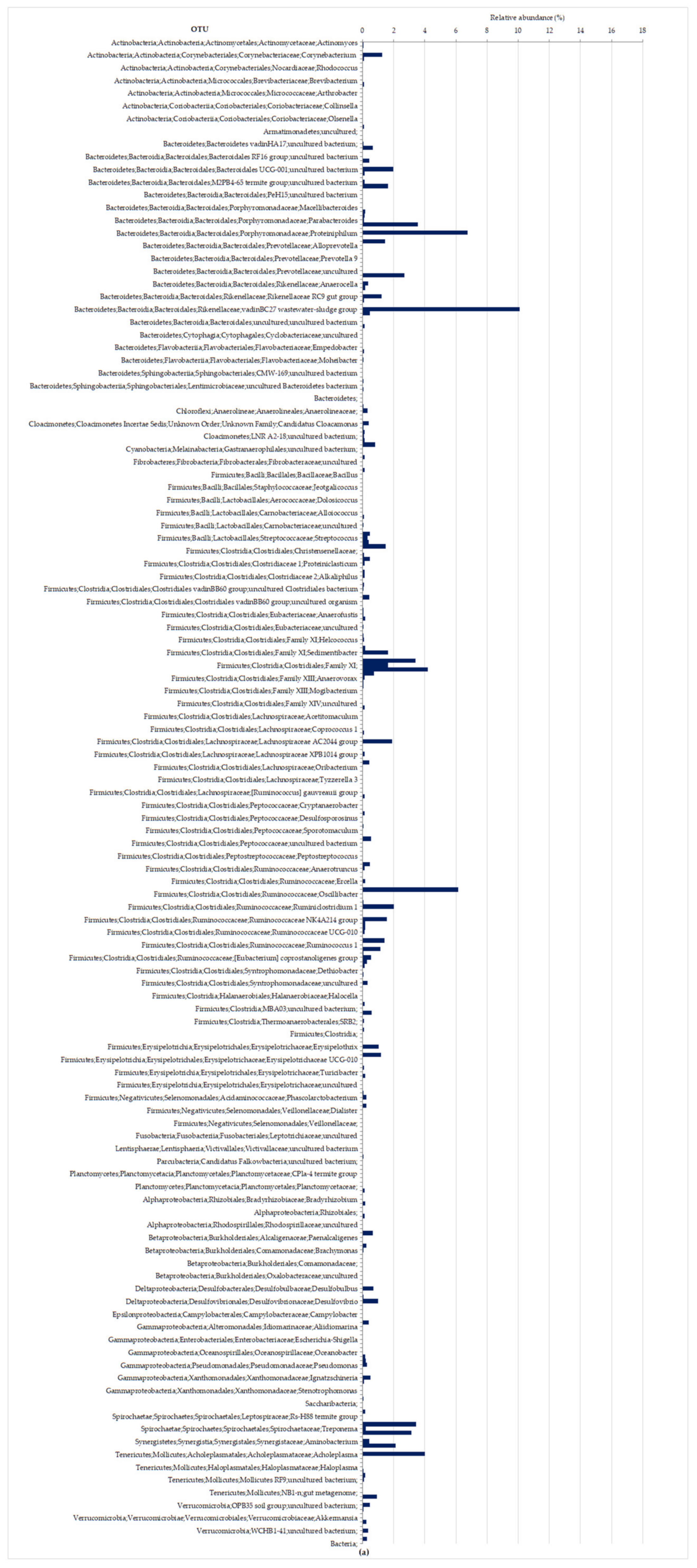
Figure 6. In the anaerobic digestion process, most of the members of Bacteroidetes were assigned to
Bacteroidales or Flavobacteriales, respectively, at 33% and 18% for HRT 6.7 d IN and HRT 3 d OUT, and most of the members of Firmicutes were assigned to
Clostridiales (35% and 37%, respectively, for HRT 6.7 d IN and HRT 3 d OUT). At the end of the HRT 3 d phase (
Figure 6b), microbial populations of the order
Flavobacteriales became predominant (18%), and consisted mainly of members of the family
Flavobacteriaceae, genus
Moheibacter (17.8%), replacing the
Bacteroidales populations.
Moheibacter has been described as a strictly aerobic bacterium and without an important role in the breakdown of complex or other carbohydrates [
68,
69], which can explain its sudden appearance at this stage. Apparently, these changes in bacterial populations are meaningless and without much influence on the performance of the process. During this phase,
Bacillales also became part of the composition structure of microbial communities (7.8%).
Bacillales, whose order consisted mainly of members of the family
Planococcaceae, and genus
Sporosarcina (7.4%), which are known as good carbohydrate-utilizing species [
60], and as
Clostridiales, also
Bacillales can decompose protein, cellulose, fat, and carbohydrates to produce VFAs [
65].
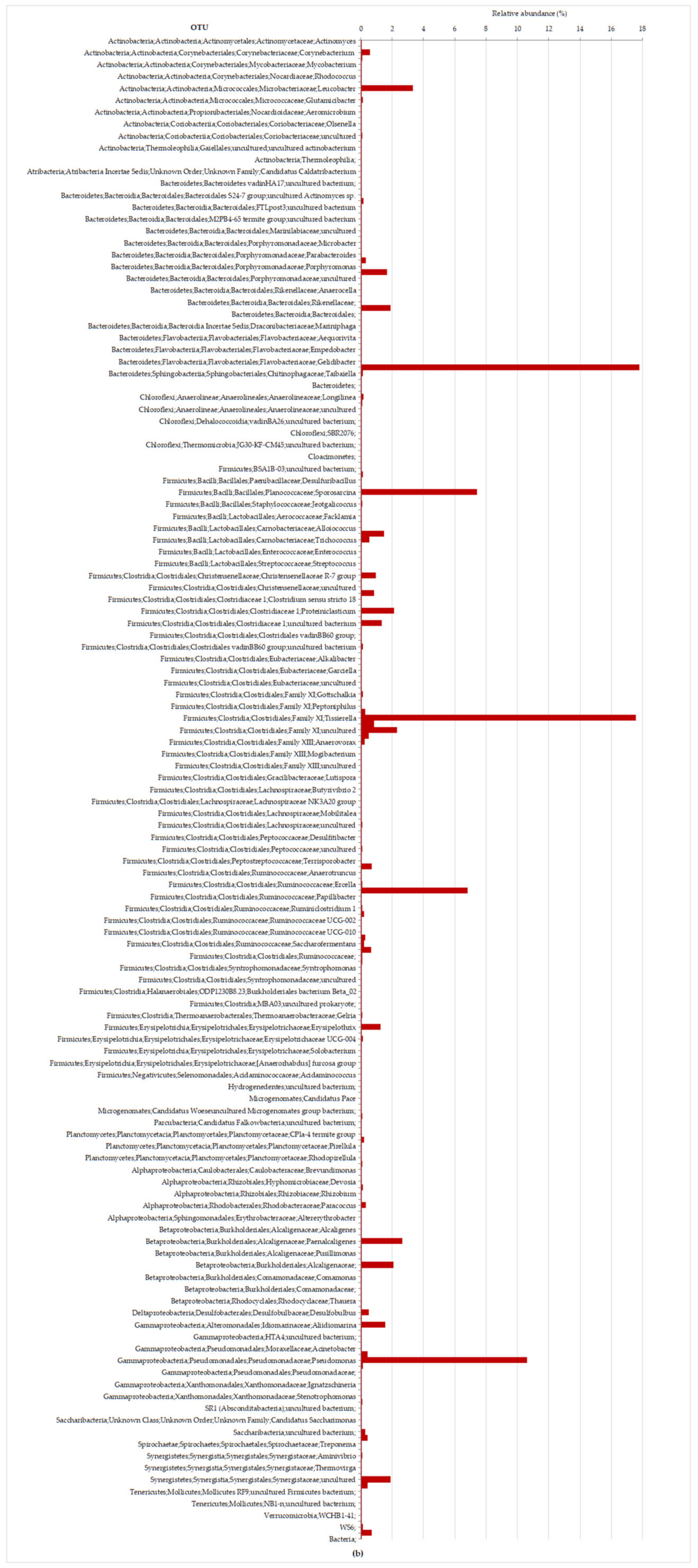
As can be observed in
Figure 5, Proteobacteria, known degraders of polysaccharides and more complex substrates [
65], predominate at phase HRT 3 d (19%). All five classes within the Proteobacteria were detected during the experiment, Beta- and Gammaproteobacteria dominated at HRT 3 d representing, respectively, 5% and 13% of the total bacterial sequences (
Figure 6b). Betaproteobacteria is involved in the first steps of degradation during the anaerobic digestion process, and they are the main consumers of propionate, butyrate, and acetate [
70]. Most of the members of Betaproteobacteria belong to the family
Alcaligenaceae that increased from 1% (6.7-d HRT,
Figure 6a) to 4.8% (3-d HRT,
Figure 6b) of total sequences. At HRT 3 d (
Figure 6b), Gammaproteobacteria represented 28,784 sequences, with the genus
Pseudomonas representing 81% of those sequences and 11% of bacterial sequences. Pseudomonadaceae is a group with important environmental functions, such as bioremediation of toxic residues, as well as xenobiotic compound degradation. The presence of
Pseudomonas in the microbiota also has been related with phenol degradation and organic matter removal, during the effluents biotreatment processes [
32].
During the anaerobic digestion of the mixture BWW + PE (60:40, v/v), slight changes in the structure of the bacterial populations were observed, mainly at HRT 3 d, in which the pronounced increase in Bacillales, Pseudomonadaceae and Alcaligenaceae certainly may have contributed to the stabilization of the process at a continuous feeding regime with OLR of 10 g COD/L·d, achieving 95% VFA removal rates, and around 2 L biogas/L·d.
Methanogenesis has three major pathways, each converting a different substrate into methane gas [
71]: methylotrophic (decarboxylation of methyl alcohols/methyl amines/methyl sulfides); hydrogenotrophic (reduction of H
2/CO
2); and acetoclastic methanogenesis (decarboxylation of acetate). The pathway which leads to the methane production occurring in an anaerobic reactor only depends on the methanogenic populations and the availability of the suitable substrates for the anaerobic digestion process.
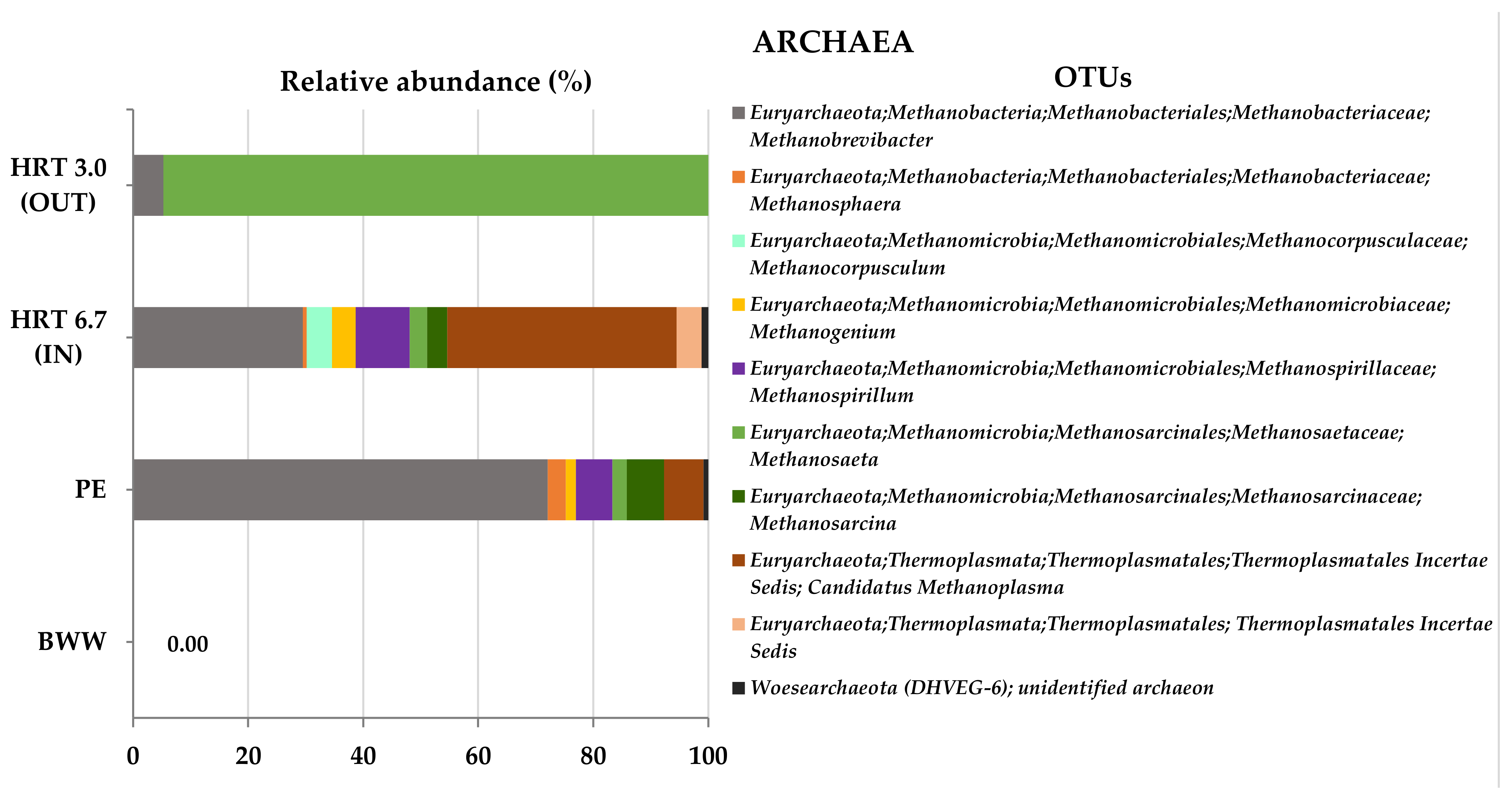
Regarding the Archaea domain,
Figure 7 shows considerable changes that occurred during anaerobic digestion in the hybrid anaerobic reactor. Of most archaeal sequences, 99% were classified within the phylum Euryarchaeota, and only 18 sequences were assigned to the phylum Woesearchaeota, present in the piggery effluent and at the beginning (HRT 6.7 d IN) of the assay, which were later not detected after anaerobic digestion. As for the Bacteria domain, Archaea also shows high diversity, 5.28 (
Table 5) on samples at the beginning of the anaerobic digestion (HRT 6.7 d IN), with predominance in archaea communities of the obligately hydrogen-dependent methylotroph
Candidatus Methanoplasma (40%) and the hydrogenotrophs
Methanobrevibacter (29%), and
Methanospirillum (9%), as shown in
Figure 7.
Since no archaea populations were detected in brewery wastewater, it can be concluded that the archaeal populations in piggery manure were crucial to further production of methane in this experiment. After the anaerobic digestion of the mixture BWW + PE (60:40,
v/
v), a considerable shift in the archaeal populations had occurred, and
Methanosaeta, an obligate acetoclastic methanogen, became the dominant genus, reaching 95% of archaea populations at the end of HRT 3 d (
Figure 7), and 84% in a later phase, HRT 1 d (data not shown). The presence of
Methanosaeta in these phases means that the methane produced during anaerobic digestion in the hybrid anaerobic reactor was formed through acetoclastic methanogenesis as the main pathway [
49,
72,
73]. These results are in line with the high biogas composition in methane (between 77% and 80%) along the anaerobic digestion (
Figure 2), indicating the presence of the active methanogenic archaea populations at all tested conditions. The important involvement of
Methanosaeta in the methanogenesis process is evident, also being reported in the literature as predominant in several anaerobic digestion systems [
49,
59,
74].
In addition, VFA concentrations (
Table 3) never reached the inhibitory level for methanogenic activity throughout the experiment and, more importantly, the limit of propionic acid concentration was maintained lower than 2000 mg/L, as stated by Lee et al. [
75], not inhibiting the methanogens.
These results show that communities of bacterial and archaeal populations were affected by the composition of the substrates mixture of brewery wastewater and piggery effluent, and by the HRT and OLR operating conditions.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}