
Airborne Fungi in Show Caves from Southern Spain

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cave Locations and Research Background
2.2. Aerobiology
3. Results
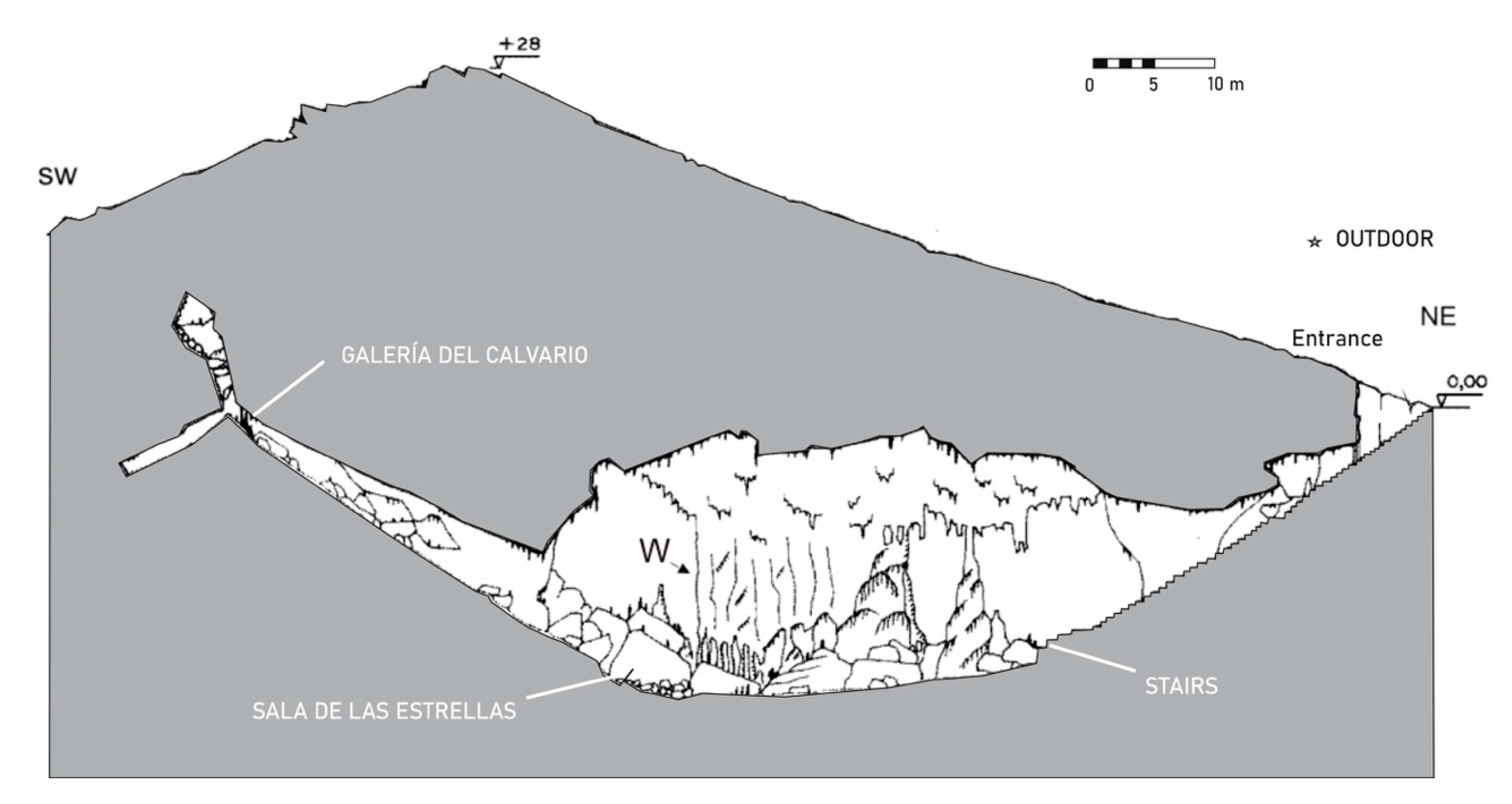
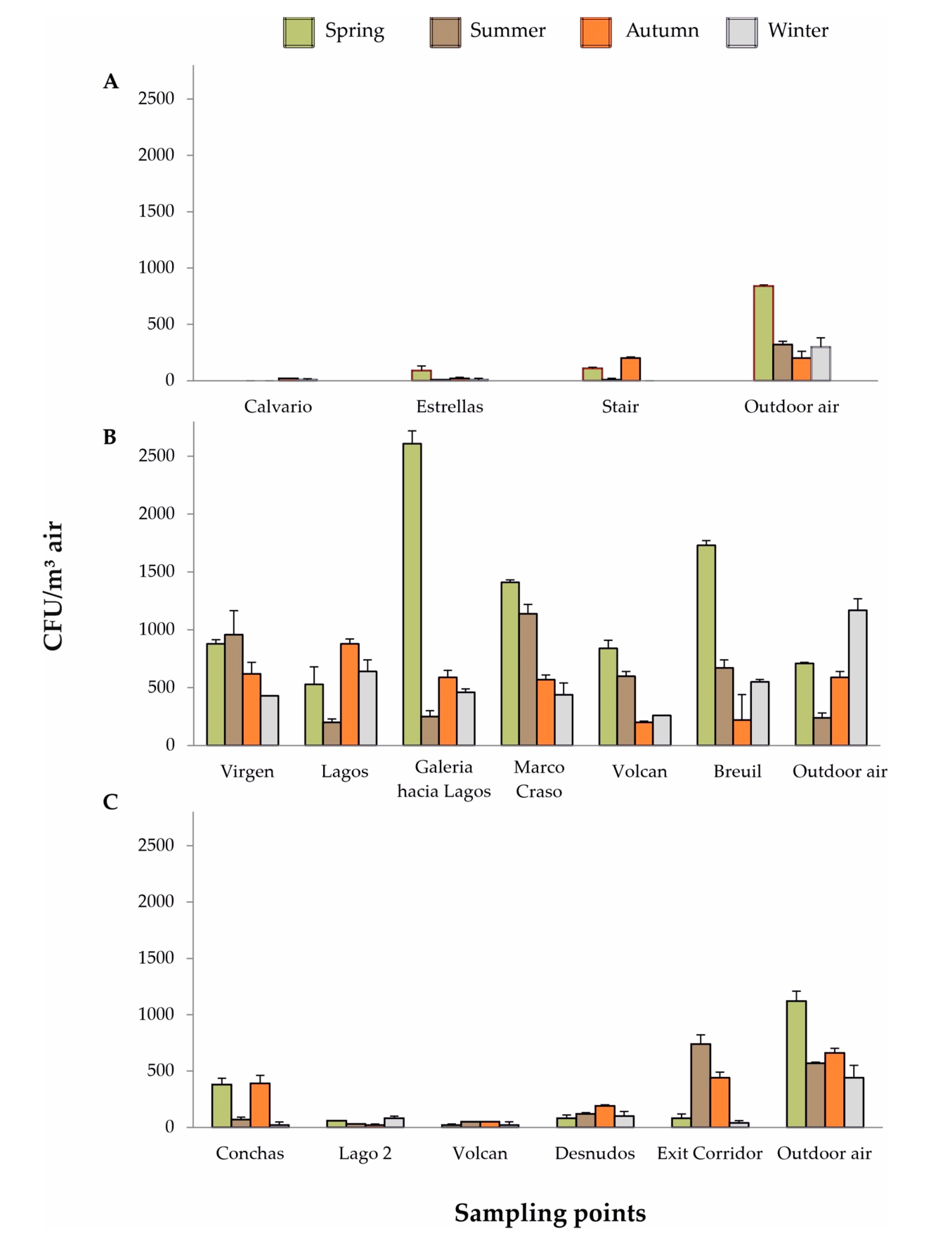
3.1. Cueva de Ardales
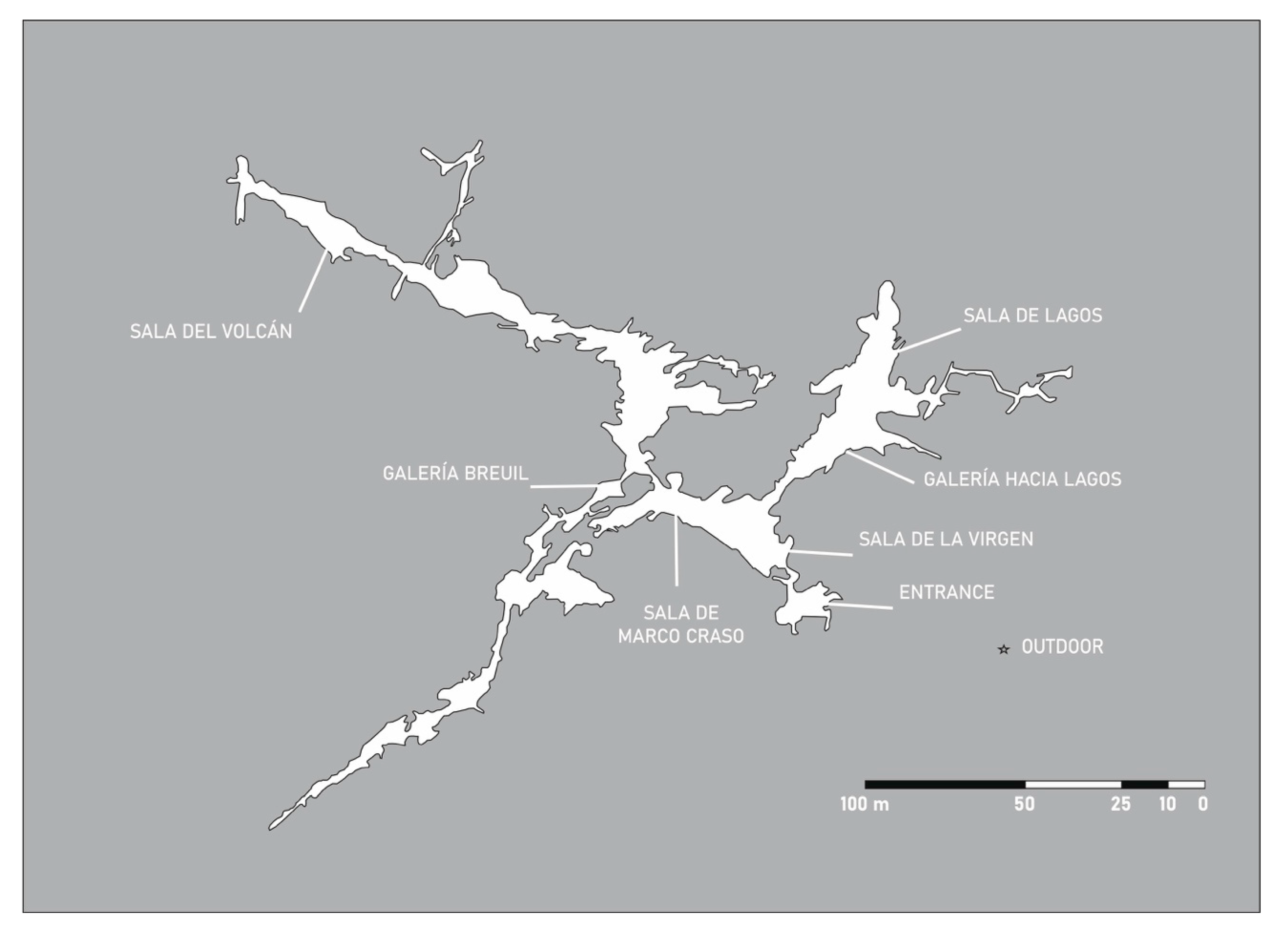
3.2. Cueva del Tesoro
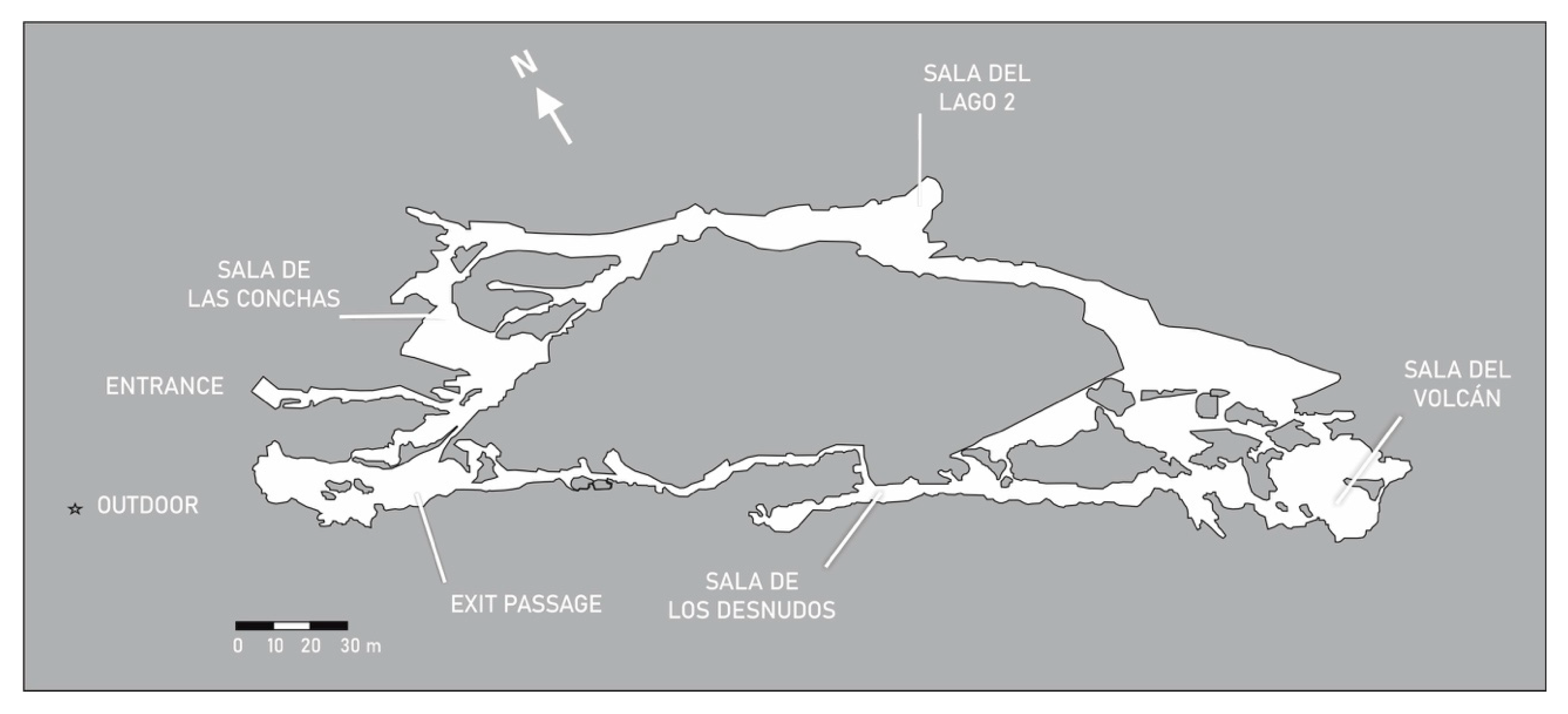
3.3. Gruta de las Maravillas
4. Discussion
4.1. Fungal Ecology
4.2. Fungal Hazard in Show Caves
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dupont, J.; Jacquet, C.; Dennetiere, B.; Lacoste, S.; Bousta, F.; Orial, G.; Cruaud, C.; Couloux, A.; Roquebert, M.F. Invasion of the French Paleolithic painted cave of Lascaux by members of the Fusarium solani species complex. Mycologia 2007, 99, 526–533. [Google Scholar]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. The impact of arthropods on fungal community structure in Lascaux Cave. J. Appl. Microbiol. 2009, 106, 1456–1462. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Porca, E.; Cuezva, S.; Fernandez-Cortes, A.; Sanchez-Moral, S.; Saiz-Jimenez, C. Fungal outbreak in a show cave. Sci. Total Environ. 2010, 408, 3632–3638. [Google Scholar] [CrossRef] [Green Version]
- Caneva, G.; De Nuntiis, P.; Fornaciari, M.; Ruga, L.; Valenti, P.; Pasquariello, G. Aerobiology applied to the preventive conservation of cultural heritage. Aerobiologia 2020, 36, 99–103. [Google Scholar] [CrossRef]
- Fernandez-Cortes, A.; Cuezva, S.; Sanchez-Moral, S.; Porca, E.; Jurado, V.; Saiz-Jimenez, C. Detection of human-induced environmental disturbances in a show cave. Environ. Sci. Pollut. Res. 2011, 18, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Porca, E.; Jurado, V.; Martin-Sanchez, P.M.; Hermosin, B.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Aerobiology: An ecological indicator for early detection and control of fungal outbreaks in caves. Ecol. Indic. 2011, 11, 1594–1598. [Google Scholar] [CrossRef]
- Garcia-Anton, E.; Cuezva, S.; Jurado, V.; Porca, E.; Miller, A.Z.; Fernandez-Cortes, A.; Saiz-Jimenez, C.; Sanchez-Moral, S. Combining stable isotope (δ13C) of trace gases and aerobiological data to monitor the entry and dispersion of microorganisms in caves. Environ. Sci. Pollut. Res. 2014, 21, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Sanchez, P.M.; Jurado, V.; Porca, E.; Bastian, F.; Lacanette, D.; Alabouvette, C.; Saiz-Jimenez, C. Airborne microorganisms in Lascaux Cave (France). Int. J. Speleol. 2014, 43, 295–303. [Google Scholar] [CrossRef]
- Nováková, A.; Kubátová, A.; Sklenář, F.; Hubka, V. Microscopic fungi on cadavers and skeletons from cave and mine environments. Czech Mycol. 2018, 70, 101–121. [Google Scholar] [CrossRef]
- Sanchez-Moral, S.; Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Ontañon, R.; Saiz-Jimenez, C. Environment-driven control of the dispersal of fungi in subterranean ecosystems: The case of La Garma Cave (northern Spain). Int. Microbiol. 2021. submitted. [Google Scholar]
- Docampo, S.; Trigo, M.M.; Recio, M.; Melgar, M.; García-Sánchez, J.; Cabezudo, B. Fungal spores content of the atmosphere of the Cave of Nerja (southern Spain): Diversity and origin. Sci. Total Environ. 2011, 409, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Moñino, I. Evaluación y Control de Comunidades Microbianas en Cuevas Turísticas. Ph.D. Thesis, Instituto de Recursos Naturales y Agrobiología, Sevilla, Spain, 2015. [Google Scholar]
- Jurado, V.; Hernández-Mariné, M.; Rogerio-Candelera, M.A.; Domínguez-Moñino, I.; Gutiérrez-Patricio, S.; Saiz-Jiménez, C. La Cueva del Tesoro: Un Paraíso Microbiano. In Cuevatur 2014/Iberoamérica Subterránea; Calaforra, J.M., Durán, J.J., Eds.; ACTE: Madrid, Spain, 2014; pp. 431–439. [Google Scholar]
- Martínez-Moreno, F.J.; Galindo-Zaldívar, J.; Pedrera, A.; Teixido, T.; Ruano, P.; Peña, J.A.; González-Castillo, L.; Ruiz-Constán, A.; López-Chicano, M.; Martín-Rosales, W. Integrated geophysical methods for studying the karst system of Gruta de las Maravillas (Aracena, Southwest Spain). J. Appl. Geophys. 2014, 107, 149–162. [Google Scholar] [CrossRef]
- Pulido-Bosch, A.; Martín-Rosales, W.; López-Chicano, M.; Vallejos, A. Human impacts in a touristic cave (Aracena, Spain). Environ. Geol. 1997, 31, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Martín-Rosales, W.; López-Chicano, M.; Rodríguez, C.M.; Pulido-Bosch, A. Morfología, espeleotemas y climatología de la Gruta de las Maravillas, Aracena (Huelva). Espeleotemas 1995, 5, 1–12. [Google Scholar]
- Martín-Rosales, W.; Cerón, J.C.; López-Chicano, M.; Fernández, I. Aspectos ambientales e hidrogeológicos de la Gruta de las Maravillas (Huelva, España). Bol. Geol. Min. 2003, 114, 247–254. [Google Scholar]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjevic, Z.; Andersen, B.; Houbraken, J.; Crous, P.W. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef]
- Segers, F.J.J.; van Laarhoven, K.A.; Huinink, H.P.; Adan, O.C.G.; Wösten, H.A.B.; Dijksterhuis, J. The indoor fungus Cladosporium halotolerans survives humidity dynamics markedly better than Aspergillus niger and Penicillium rubens despite less growth at lowered steady-state water activity. Appl. Environ. Microbiol. 2016, 82, 5089–5098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nováková, A. Microscopic fungi isolated from the Domica Cave system (Slovak Karst National Park, Slovakia). A review. Int. J. Speleol. 2009, 38, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.M.; Sung, G.H.; Ban, S.; Nakagiri, A.; Chen, M.J.; Huang, B.; Li, Z.; Spatafora, J.W. New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 2012, 104, 182–197. [Google Scholar] [CrossRef]
- Kim, H.S.; Adhikari, M.; Yadav, D.R.; Kim, S.W.; Um, Y.H.; Lee, H.B.; Lee, Y.S. First report of Metacordyceps chlamydosporia (Cordyceps chlamydosporia) isolated from soil in Korea. Korean J. Med. Mycol. 2016, 44, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeter. Biodegr. 2008, 62, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Zare, R.; Gams, W. More white verticillium-like anamorphs with erect conidiophores. Mycol. Progress 2016, 15, 993–1030. [Google Scholar] [CrossRef]
- Okane, I.; Nonaka, K.; Kurihara, Y.; Abe, J.P.; Yamaoka, Y. A new species of Leptobacillium, L. symbioticum, isolated from mites and sori of soybean rust. Mycoscience 2020, 61, 165–171. [Google Scholar] [CrossRef]
- Bastian, F.; Jurado, V.; Nováková, A.; Alabouvette, C.; Saiz-Jimenez, C. The microbiology of the Lascaux Cave. Microbiology 2010, 156, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, A.; Costa-Comelles, J.; Alonso, A.; Rodríguez, J.M.; Ferrer, J. Ensayo del hongo entomopatógeno Beauveria bassiana (Balsamo) Vuillemin para el control de la mosca blanca de los cítricos Aleurothrixus floccosus (Maskell) (Homoptera: Aleyrodidae) y su acción sobre el parásito Cales noacki (Howard) (Hymenoptera: Aphelinidae). Bol. San. Veg. Plagas 1998, 24, 695–706. [Google Scholar]
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwigia. 2001, 73, 1–50. [Google Scholar] [CrossRef]
- Castillo, S.Y.; Bernal Vega, J.A.; Lezcano, J.; Piepenbring, M.; Cáceres, O. Hongos entomopatógenos asociados a insectos recolectados en plantaciones de café en el oeste de Panamá. Tecnociencia 2013, 15, 29–39. [Google Scholar]
- Chang, S.; Lui, W. Analysis of the mating types of Schizophyllum commune in the natural population of Hong Kong. Bot. Bull. Acad. Sin. 1969, 10, 74–88. [Google Scholar]
- Nováková, A.; Hubka, V.; Saiz-Jimenez, C.; Kolarik, M. Aspergillus baeticus sp. nov. and Aspergillus thesauricus sp. nov.: Two new species in section Usti originating from Spanish caves. Int. J. Syst. Evol. Microbiol. 2012, 62, 2778–2785. [Google Scholar] [CrossRef] [PubMed]
- Grbić, M.L.; Vukojević, J.; Simić, G.S.; Krizmanić, J.; Stupar, M. Biofilm forming cyanobacteria, algae and fungi on two historic monuments in Belgrade, Serbia. Arch. Biol. Sci. 2010, 62, 625–631. [Google Scholar] [CrossRef]
- Gallego-Cartagena, E.; Morillas, H.; Maguregui, M.; Patiño-Camelo, K.; Marcaida, I.; Morgado-Gamero, W.; Silva, L.F.O.; Madariaga, J.M. A comprehensive study of biofilms growing on the built heritage of a Caribbean industrial city in correlation with construction materials. Int. Biodeter. Biodegr. 2020, 147, 104874. [Google Scholar] [CrossRef]
- Vaughan-Martini, A.; Angelini, P.; Zacchi, L. The influence of human and animal visitation on the yeast ecology of three Italian caverns. Ann. Microbiol. 2000, 50, 133–140. [Google Scholar]
- Sugita, T.; Kikuchi, K.; Makimura, K.; Urata, K.; Someya, T.; Kamei, K.; Niimi, M.; Uehara, Y. Trichosporon species isolated from guano samples obtained from bat-inhabited caves in Japan. Appl. Environ. Microbiol. 2005, 71, 7626–7629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashima, M.; Kurakado, S.; Cho, O.; Kikuchi, K.; Sugiyama, J.; Sugita, T. Description of four Apiotrichum and two Cutaneotrichosporon species isolated from guano samples from bat-inhabited caves in Japan. Int. J. Syst. Evol. Microbiol. 2020, 70, 4458–4469. [Google Scholar] [CrossRef] [PubMed]
- Porca Belío, E. Aerobiología: Mecanismos de Dispersión de los Microorganismos en Cuevas Turísticas. Ph.D. Thesis, Instituto de Recursos Naturales y Agrobiología, Sevilla, Spain, 2011. [Google Scholar]
- Urquhart, E.J.; Menzies, J.G.; Punja, Z.K. Growth and biological control activity of Tilletiopsis species against powdery mildew (Sphaerotheca fuliginea) on greenhouse cucumber. Phytopathology 1994, 84, 341–351. [Google Scholar] [CrossRef]
- Fonseca, A.; Boekhout, T.; Fell, J.W. Cryptococcus Vuillemin (1901). In The Yeasts, a Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1661–1737. [Google Scholar]
- Brentassi, M.E.; Medina, R.; de la Fuente, D.; Franco, M.E.E.; Toledo, A.V.; Saparrat, M.C.N.; Balatti, P.A. Endomycobiome associated with females of the planthopper Delphacodes kuscheli (Hemiptera: Delphacidae): A metabarcoding approach. Helyon 2020, 6, e04634. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Sutton, D.A.; Martin-Vicente, A.; Cano-Lira, J.F.; Wiederhold, N.; Guarro, J.; Gené, J. Cladosporium species recovered from clinical samples in the United States. J. Clin. Microbiol. 2015, 53, 2990–3000. [Google Scholar] [CrossRef] [Green Version]
- Tucker, D.L.; Beresford, C.H.; Sigler, L.; Rogers, K. Disseminated Beauveria bassiana infection in a patient with acute lymphoblastic leukemia. J. Clin. Microbiol. 2004, 42, 5412–5414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, C.-C.; Chan, J.F.W.; Pong, W.-M.; Chen, J.H.K.; Ngan, A.H.Y.; Cheung, M.; Lai, C.K.C.; Tsang, D.N.C.; Lau, S.K.P.; Woo, P.C.Y. Cutaneous hyalohyphomycosis due to Parengyodontium album gen. et comb. nov. Med. Mycol. 2016, 54, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Yi, Y.; Cho, S.-Y.; Lee, D.-G.; Chun, H.-S.; Park, C.; Kim, Y.-J.; Park, Y.-J. Pneumonia due to Schizophyllum commune in a patient with acute myeloid leukemia: Case report and literature review. Infect. Chemother. 2021, 53, e67. [Google Scholar]
- Bastos, R.W.; Valero, C.; Silva, L.P.; Schoen, T.; Drott, M.; Brauer, V.; Silva-Rocha, R.; Lind, A.; Steenwyk, J.L.; Rokas, A.; et al. Functional characterization of clinical isolates of the opportunistic fungal pathogen Aspergillus nidulans. mSphere 2020, 5, e00153-20. [Google Scholar] [CrossRef] [Green Version]
- Geltner, C.; Lass-Flörl, C.; Bonatti, H.; Müller, L.; Stelzmüller, I. Invasive pulmonary mycosis due to Penicillium chrysogenum: A new invasive pathogen. Transplantation 2013, 95, e21–e23. [Google Scholar] [CrossRef]
- Cooper, C.R. Yeasts Pathogenic to Humans. In The Yeasts, a Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 9–19. [Google Scholar]
- Mariné, M.; Brown, N.A.; Riaño-Pachón, D.M.; Goldman, G.H. On and under the skin: Emerging basidiomycetous yeast infections caused by Trichosporon species. PLoS Pathog. 2015, 11, e1004982. [Google Scholar] [CrossRef]
- Morales-López, S.E.; Garcia-Effron, G. Infections due to rare Cryptococcus species. A literature review. J. Fungi 2021, 7, 279. [Google Scholar] [CrossRef]
- Jurado, V.; Laiz, L.; Rodriguez-Nava, V.; Boiron, P.; Hermosin, B.; Sanchez-Moral, S.; Saiz-Jimenez, C. Pathogenic and opportunistic microorganisms in caves. Int. J. Speleol. 2010, 39, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Zucconi, L.; Gagliardi, M.; Isola, D.; Onofri, S.; Andaloro, M.C.; Pelosi, C.; Pogliani, P.; Selbmann, L. Biodeterioration agents dwelling in or on the wall paintings of the Holy Saviour’s cave (Vallerano, Italy). Int. Biodeter. Biodegr. 2012, 70, 40–46. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Višňovská, Z.; Tancinová, D.; Zalewski, D. Speleomycology of air and rock surfaces in Driny Cave (Lesser Carpathians, Slovakia). J. Cave Karst Stud. 2016, 78, 119–127. [Google Scholar] [CrossRef]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium album, a frequently reported fungal species in the cultural heritage environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Pyzik, A.; Ciuchcinski, K.; Dziurzynski, M.; Dziewit, L. The Bad and the good—Microorganisms in cultural heritage environments—An update on biodeterioration and biotreatment approaches. Materials 2021, 14, 177. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Jimenez, C.; Miller, A.Z.; Martin-Sanchez, P.M.; Hernandez-Marine, M. Uncovering the origin of the black stains in Lascaux Cave in France. Environ. Microbiol. 2012, 14, 3220–3231. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dominguez-Moñino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne Fungi in Show Caves from Southern Spain. Appl. Sci. 2021, 11, 5027. https://doi.org/10.3390/app11115027
Dominguez-Moñino I, Jurado V, Rogerio-Candelera MA, Hermosin B, Saiz-Jimenez C. Airborne Fungi in Show Caves from Southern Spain. Applied Sciences. 2021; 11(11):5027. https://doi.org/10.3390/app11115027
Chicago/Turabian StyleDominguez-Moñino, Irene, Valme Jurado, Miguel Angel Rogerio-Candelera, Bernardo Hermosin, and Cesareo Saiz-Jimenez. 2021. "Airborne Fungi in Show Caves from Southern Spain" Applied Sciences 11, no. 11: 5027. https://doi.org/10.3390/app11115027
APA StyleDominguez-Moñino, I., Jurado, V., Rogerio-Candelera, M. A., Hermosin, B., & Saiz-Jimenez, C. (2021). Airborne Fungi in Show Caves from Southern Spain. Applied Sciences, 11(11), 5027. https://doi.org/10.3390/app11115027