
Role of Quercetin in Depressive-Like Behaviors: Findings from Animal Models


Abstract
:1. Introduction
2. Methodology
3. Quercetin
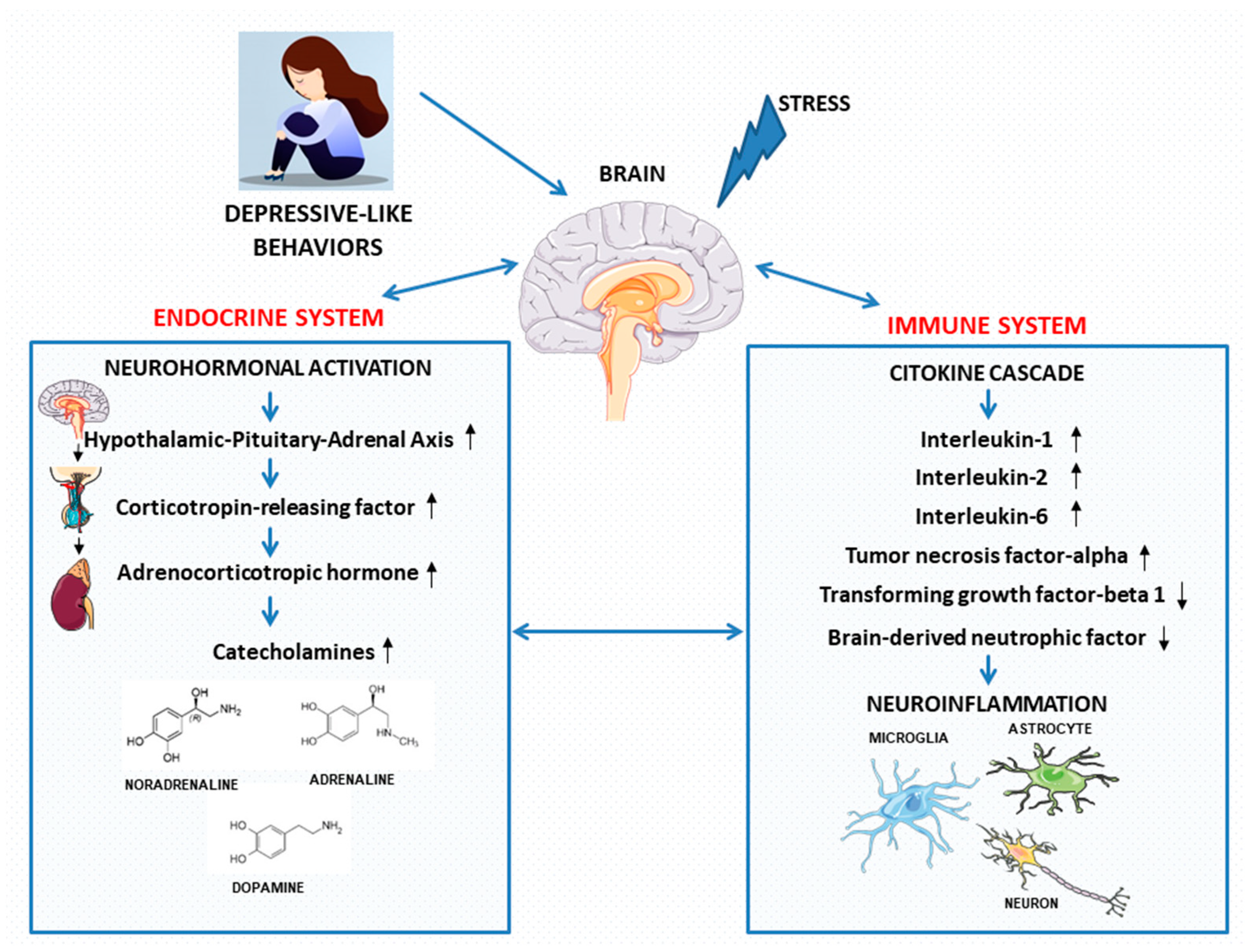
4. Depressive-Like Behaviors
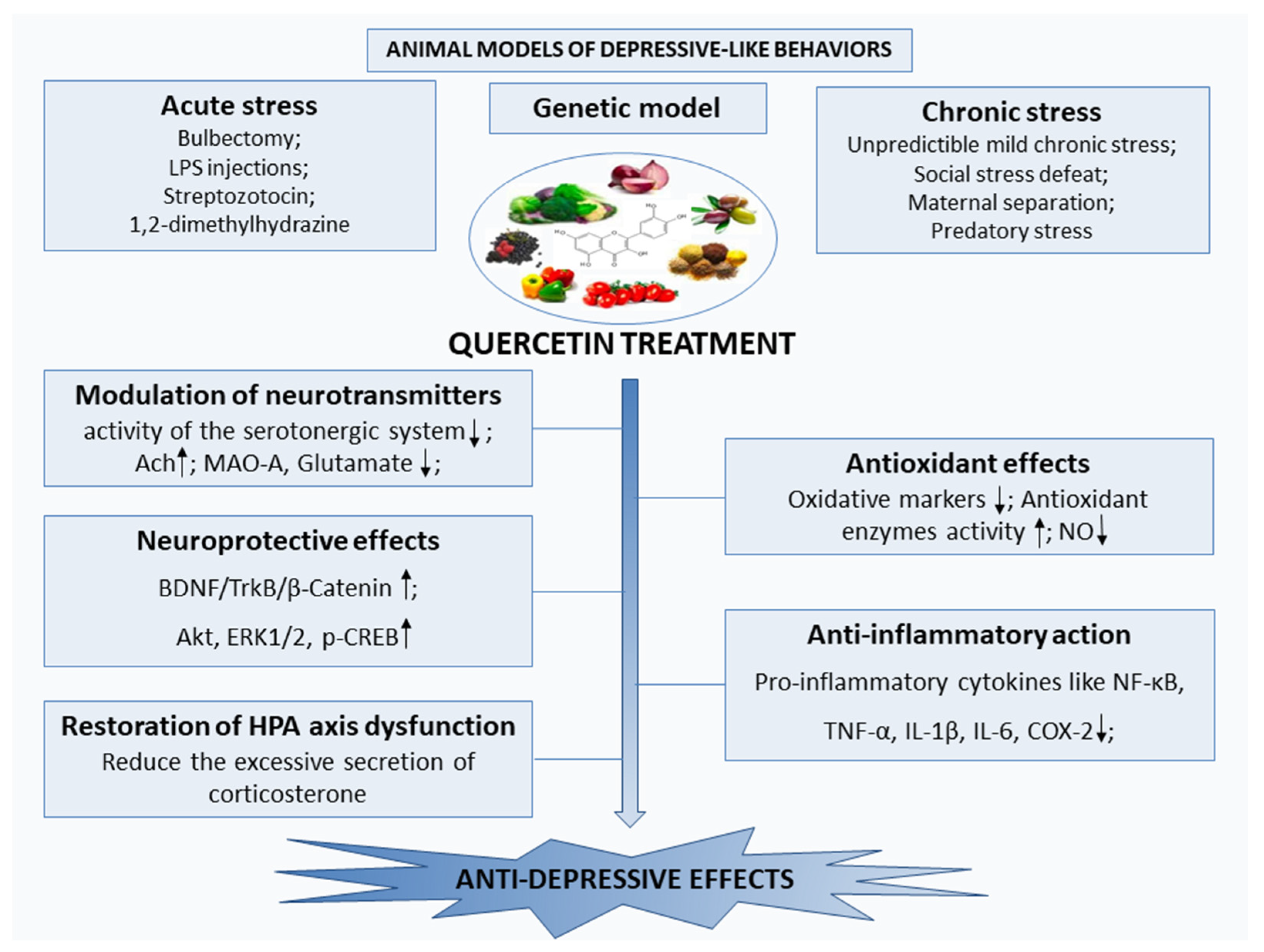
5. Quercetin in Animal Models of Depressive-Like Behaviors
5.1. Quercetin Effects in Animal Models Exposed to Chronic Stress
5.2. Quercetin Effects in Animal Models Exposed to Acute Stress
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahati, K.; Bhagya, V.; Christofer, T.; Sneha, A.; Rao, B.S. Enriched environment ameliorates depression-induced cognitive deficits and restores abnormal hippocampal synaptic plasticity. Neurobiol. Learn. Mem. 2016, 134 Pt B, 379–391. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Cha, D.S.; Soczynska, J.K.; Woldeyohannes, H.O.; Gallaugher, L.A.; Kudlow, P.; Alsuwaidan, M.; Baskaran, A. Cognitive deficits and functional outcomes in major depressive disorder: Determinants, substrates, and treatment interventions. Depress. Anxiety 2013, 30, 515–527. [Google Scholar] [CrossRef]
- Frodl, T.; Reinhold, E.; Koutsouleris, N.; Reiser, M.; Meisenzahl, E.M. Interaction of childhood stress with hippocampus and prefrontal cortex volume reduction in major depression. J. Psychiatr. Res. 2010, 44, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Desmyter, S.; van Heeringen, C.; Audenaert, K. Structural and functional neuroimaging studies of the suicidal brain. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, C.; Duman, R.S. Stress, depression, and neuroplasticity: A convergence of mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef]
- Pariante, C.M.; Miller, A.H. Glucocorticoid receptors in major depression: Relevance to pathophysiology and treatment. Biol. Psychiatry 2001, 49, 391–404. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [PubMed]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal relationship between stressful life events and the onset of major depression. Am. J. Psychiatry 1999, 156, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Racagni, G.; Popoli, M. The pharmacological properties of antidepressants. Int. Clin. Psychopharmacol. 2010, 25, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhir, A. Investigational drugs for treating major depressive disorder. Expert Opin. Investig. Drugs 2017, 26, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Falzon, C.C.; Balabanova, A. Phytotherapy: An Introduction to Herbal Medicine. Prim. Care 2017, 44, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Bakre, A.G.; Olayemi, J.O.; Ojo, O.R.; Odusanya, S.T.; Agu, G.A.; Aderibigbe, A.O. Antidepressant-like effect of ethanol extract of Blighia unijugata Bak. (Sapindaceae) leaves in acute and chronic models of depression in mice. Niger. J. Physiol. Sci. 2019, 34, 191–199. [Google Scholar]
- Chavez-Morales, Y.; Jimenez-Ferrer, E.; Martinez-Hernandez, G.B.; Tortoriello, J.; Roman-Ramos, R.; Zamilpa, A.; Herrera-Ruiz, M. Effect of Standardized Fractions and Tiliroside from Leaves of Tilia americana on Depression Tests in Mice. Iran. J. Pharm. Res. 2019, 18, 1931–1946. [Google Scholar] [PubMed]
- Dovichi, S.; Lajolo, F. Flavonoid and Their Relationship to Diseases of The Central Nervous System. J. Braz. Sociaty Food Nutr. Sao Paulo 2011, 36, 123–135. [Google Scholar]
- Gonzalez-Aguilar, G.; Robles-Sánchez, R.M.; Martínez-Téllez, M.A.; Olivas, G.I.; Alvarez-Parrilla, E.; De La Rosa, L.A. Bioactive compounds in fruits: Health benefits and effect of storage conditions. Stewart Postharvest Rev. 2008, 4, 1–10. [Google Scholar]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Heo, H.J.; Lee, C.Y. Protective effects of quercetin and vitamin C against oxidative stress-induced neurodegeneration. J. Agric. Food Chem. 2004, 52, 7514–7517. [Google Scholar] [CrossRef]
- Youdim, K.A.; Shukitt-Hale, B.; Joseph, J.A. Flavonoids and the brain: Interactions at the blood-brain barrier and their physiological effects on the central nervous system. Free Radic. Biol. Med. 2004, 37, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Dok-Go, H.; Lee, K.H.; Kim, H.J.; Lee, E.H.; Lee, J.; Song, Y.S.; Lee, Y.H.; Jin, C.; Lee, Y.S.; Cho, J. Neuroprotective effects of antioxidative flavonoids, quercetin, (+)-dihydroquercetin and quercetin 3-methyl ether, isolated from Opuntia ficus-indica var. saboten. Brain Res. 2003, 965, 130–136. [Google Scholar] [CrossRef]
- Haleagrahara, N.; Radhakrishnan, A.; Lee, N.; Kumar, P. Flavonoid quercetin protects against swimming stress-induced changes in oxidative biomarkers in the hypothalamus of rats. Eur. J. Pharm. 2009, 621, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Paulke, A.; Schubert-Zsilavecz, M.; Wurglics, M. Determination of St. John’s wort flavonoid-metabolites in rat brain through high performance liquid chromatography coupled with fluorescence detection. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2006, 832, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kim, I.S.; Jang, Y.H.; Kim, A.R.; Lee, S.R. Protective effect of quercetin, a natural flavonoid against neuronal damage after transient global cerebral ischemia. Neurosci. Lett. 2006, 404, 330–335. [Google Scholar] [CrossRef]
- Rojas, P.; Serrano-Garcia, N.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Ogren, S.O.; Rojas, C. Antidepressant-like effect of a Ginkgo biloba extract (EGb761) in the mouse forced swimming test: Role of oxidative stress. Neurochem. Int. 2011, 59, 628–636. [Google Scholar] [CrossRef]
- Sakakibara, H.; Yoshino, S.; Kawai, Y.; Terao, J. Antidepressant-like effect of onion (Allium cepa L.) powder in a rat behavioral model of depression. Biosci. Biotechnol. Biochem. 2008, 72, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Bhutada, P.; Mundhada, Y.; Bansod, K.; Ubgade, A.; Quazi, M.; Umathe, S.; Mundhada, D. Reversal by quercetin of corticotrophin releasing factor induced anxiety- and depression-like effect in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 955–960. [Google Scholar] [CrossRef]
- Kawabata, K.; Kawai, Y.; Terao, J. Suppressive effect of quercetin on acute stress-induced hypothalamic-pituitary-adrenal axis response in Wistar rats. J. Nutr. Biochem. 2010, 21, 374–380. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samad, N.; Saleem, A.; Yasmin, F.; Shehzad, M.A. Quercetin protects against stress-induced anxiety- and depression-like behavior and improves memory in male mice. Physiol. Res. 2018, 67, 795–808. [Google Scholar] [CrossRef]
- Singh, V.; Chauhan, G.; Shri, R. Anti-depressant like effects of quercetin 4’-O-glucoside from Allium cepa via regulation of brain oxidative stress and monoamine levels in mice subjected to unpredictable chronic mild stress. Nutr. Neurosci. 2021, 24, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Najmi, A.; Akhtar, M. A Natural Phenolic Compound Quercetin Showed the Usefulness by Targeting Inflammatory, Oxidative Stress Markers and Augment 5-HT Levels in One of the Animal Models of Depression in Mice. Drug Res. 2019, 69, 392–400. [Google Scholar] [CrossRef]
- Mehta, V.; Parashar, A.; Udayabanu, M. Quercetin prevents chronic unpredictable stress induced behavioral dysfunction in mice by alleviating hippocampal oxidative and inflammatory stress. Physiol. Behav. 2017, 171, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Xu, Y.; Jiang, X.; Wang, Z.; Guo, Y.; Pan, W.; Hou, J. Avicularin Relieves Depressive-Like Behaviors Induced by Chronic Unpredictable Mild Stress in Mice. Med. Sci. Monit. 2019, 25, 2777–2784. [Google Scholar] [CrossRef]
- Ma, Z.X.; Zhang, R.Y.; Rui, W.J.; Wang, Z.Q.; Feng, X. Quercetin alleviates chronic unpredictable mild stress-induced depressive-like behaviors by promoting adult hippocampal neurogenesis via FoxG1/CREB/BDNF signaling pathway. Behav. Brain Res. 2021, 406, 113245. [Google Scholar] [CrossRef]
- Donoso, F.; Egerton, S.; Bastiaanssen, T.F.S.; Fitzgerald, P.; Gite, S.; Fouhy, F.; Ross, R.P.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Polyphenols selectively reverse early-life stress-induced behavioural, neurochemical and microbiota changes in the rat. Psychoneuroendocrinology 2020, 116, 104673. [Google Scholar] [CrossRef]
- Sahin, T.D.; Gocmez, S.S.; Duruksu, G.; Yazir, Y.; Utkan, T. Resveratrol and quercetin attenuate depressive-like behavior and restore impaired contractility of vas deferens in chronic stress-exposed rats: Involvement of oxidative stress and inflammation. Naunyn Schmiedebergs Arch. Pharm. 2020, 393, 761–775. [Google Scholar] [CrossRef]
- Anggreini, P.; Ardianto, C.; Rahmadi, M.; Khotib, J. Quercetin attenuates acute predator stress exposure-evoked innate fear and behavioral perturbation. J. Basic Clin. Physiol. Pharm. 2019, 30. [Google Scholar] [CrossRef]
- Zhang, J.; Ning, L.; Wang, J. Dietary quercetin attenuates depressive-like behaviors by inhibiting astrocyte reactivation in response to stress. Biochem. Biophys. Res. Commun. 2020, 533, 1338–1346. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.; Cao, C.; Hou, Y.; Li, Y.; Wei, X.; Li, S.; Jia, S.; Zhao, X. Effects of quercetin on the alterations of serum elements in chronic unpredictable mild stress-induced depressed rats. Biometals 2021, 34, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Li, H.R.; Chen, X.X.; Gao, X.R.; Huang, L.L.; Du, A.Q.; Jiang, C.; Li, H.; Ge, J.F. Quercetin Alleviates LPS-Induced Depression-Like Behavior in Rats via Regulating BDNF-Related Imbalance of Copine 6 and TREM1/2 in the Hippocampus and PFC. Front. Pharm. 2019, 10, 1544. [Google Scholar] [CrossRef] [PubMed]
- Sadighparvar, S.; Darband, S.G.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Babaei, G.; Mobaraki, K.; Majidinia, M. Combination of quercetin and exercise training attenuates depression in rats with 1,2-dimethylhydrazine-induced colorectal cancer: Possible involvement of inflammation and BDNF signalling. Exp. Physiol. 2020, 105, 1598–1609. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Lei, C.; Lei, X.; Zhu, X.; Yang, L.; Zhang, R. Quercetin exerts antidepressant and cardioprotective effects in estrogen receptor alpha-deficient female mice via BDNF-AKT/ERK1/2 signaling. J. Steroid Biochem. Mol. Biol. 2021, 206, 105795. [Google Scholar] [CrossRef]
- Merzoug, S.; Toumi, M.L.; Tahraoui, A. Quercetin mitigates Adriamycin-induced anxiety- and depression-like behaviors, immune dysfunction, and brain oxidative stress in rats. Naunyn Schmiedebergs Arch. Pharm. 2014, 387, 921–933. [Google Scholar] [CrossRef]
- Demir, E.A.; Gergerlioglu, H.S.; Oz, M. Antidepressant-like effects of quercetin in diabetic rats are independent of hypothalamic-pituitary-adrenal axis. Acta Neuropsychiatr. 2016, 28, 23–30. [Google Scholar] [CrossRef]
- Holzmann, I.; da Silva, L.M.; Correa da Silva, J.A.; Steimbach, V.M.; de Souza, M.M. Antidepressant-like effect of quercetin in bulbectomized mice and involvement of the antioxidant defenses, and the glutamatergic and oxidonitrergic pathways. Pharm. Biochem. Behav. 2015, 136, 55–63. [Google Scholar] [CrossRef]
- Rinwa, P.; Kumar, A. Quercetin suppress microglial neuroinflammatory response and induce antidepressent-like effect in olfactory bulbectomized rats. Neuroscience 2013, 255, 86–98. [Google Scholar] [CrossRef]
- Nuutila, A.M.; Puupponen-Pimia, R.; Aarni, M.; Oksman-Caldentey, K.M. Comparison of antioxidant activities of onion and garlic extracts by inhibition of lipid peroxidation and radical scavenging activity. Food Chem. 2003, 81, 485–493. [Google Scholar] [CrossRef]
- Singh, J.; Upadhyay, A.K.; Prasad, K.; Bahadur, A.; Rai, M. Variability of carotenes, vitamin C, E and phenolics in Brassica vegetables. J. Food Compos. Anal. 2007, 20, 106–112. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomas-Barberan, F.; García-Viguera, C. Effect of climatic and sulphur fertilisation conditions, on phenolic compounds and vitamin C, in the inflorescences of eight broccoli cultivars. Eur. Food Res. Technol. 2003, 216, 395–401. [Google Scholar] [CrossRef]
- Gliszczynska-Swiglo, A.; Kaluzewicz, A.; Lemanska, K.; Knaflewski, M.; Tyrakowska, B. The effect of solar radiation on the flavonol content in broccoli inflorescence. Food Chem. 2007, 100, 241–245. [Google Scholar] [CrossRef]
- Mogren, L.M.; Olsson, M.E.; Gertsson, U.E. Effects of cultivar, lifting time and nitrogen fertiliser level on quercetin content in onion(Allium cepa L.) at lifting. J. Sci. Food Agric. 2007, 87, 470–476. [Google Scholar] [CrossRef]
- Patil, B.S.; Pike, L.M.; Hamilton, B.K. Changes in Quercetin Concentration in Onion (Allium cepa L.) Owing to Location, Growth Stage and Soil Type. New Phytol. 1995, 130, 349–355. [Google Scholar] [CrossRef]
- Aaby, K.; Wrolstad, R.E.; Ekeberg, D.; Skrede, G. Polyphenol composition and antioxidant activity in strawberry purees; impact of achene level and storage. J. Agric. Food Chem. 2007, 55, 5156–5166. [Google Scholar] [CrossRef] [PubMed]
- Rawson, A.; Koidis, A.; Rai, D.K.; Tuohy, M.; Brunton, N. Influence of Sous Vide and water immersion processing on polyacetylene content and instrumental color of parsnip (Pastinaca sativa) disks. J. Agric. Food Chem. 2010, 58, 7740–7747. [Google Scholar] [CrossRef]
- Volden, J.; Bengtsson, G.B.; Wicklund, T. Glucosinolates, L-ascorbic acid, total phenols, anthocyanins, antioxidant capacities and colour in cauliflower (Brassica oleracea L. ssp. botrytis); effects of long-term freezer storage. Food Chem. 2009, 112, 967–976. [Google Scholar] [CrossRef]
- Rodrigues, A.; Pérez-Gregorio, M.; García-Falcón, M.; Simal-Gándara, J. Effect of curing and cooking on flavonols and anthocyanins in traditional varieties of onion bulbs. Food Res. Int. 2009, 42, 1331–1336. [Google Scholar] [CrossRef]
- Gennaro, L.; Leonardi, C.; Esposito, F.; Salucci, M.; Maiani, G.; Quaglia, G.; Fogliano, V. Flavonoid and carbohydrate contents in Tropea red onions: Effects of homelike peeling and storage. J. Agric. Food Chem. 2002, 50, 1904–1910. [Google Scholar] [CrossRef]
- Dinelli, G.; Bonetti, A.; Minelli, M.; Marotti, I.; Catizone, P.; Mazzanti, A. Content of flavonols in Italian bean (Phaseolus vulgaris L.) ecotypes. Food Chem. 2006, 99, 105–114. [Google Scholar] [CrossRef]
- Elizabeth, R.G.N.; Annete, H.; Francisco, G.L.R.; Javier, I.P.F.; Graciela, Z.G.; Alberto, G.I.J. Antioxidant and antimutagenic activity of phenolic compounds in three different colour groups of common bean cultivars (Phaseolus vulgaris). Food Chem. 2007, 103, 521–527. [Google Scholar]
- Manach, C.; Mazur, A.; Scalbert, A. Polyphenols and prevention of cardiovascular diseases. Curr. Opin. Lipidol. 2005, 16, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.E.; Hong, Y.J.; Koh, E.; Barrett, D.M.; Bryant, D.E.; Denison, R.F.; Kaffka, S. Ten-year comparison of the influence of organic and conventional crop management practices on the content of flavonoids in tomatoes. J. Agric. Food Chem. 2007, 55, 6154–6159. [Google Scholar] [CrossRef] [PubMed]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, C.G.; Haenen, G.R.; Oostveen, R.M.; Stalpers, E.M.; Bast, A. Protection of flavonoids against lipid peroxidation: The structure activity relationship revisited. Free Radic. Res. 2002, 36, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadzak, A.; Mravljak, J.; Maltar-Strmecki, N.; Arsov, Z.; Baranovic, G.; Erceg, I.; Kriechbaum, M.; Strasser, V.; Pribyl, J.; Segota, S. The Structural Integrity of the Model Lipid Membrane during Induced Lipid Peroxidation: The Role of Flavonols in the Inhibition of Lipid Peroxidation. Antioxidants 2020, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Luiz da Silva, E.; Tsushida, T.; Terao, J. Inhibition of mammalian 15-lipoxygenase-dependent lipid peroxidation in low-density lipoprotein by quercetin and quercetin monoglucosides. Arch. Biochem. Biophys. 1998, 349, 313–320. [Google Scholar] [CrossRef]
- Yamamoto, N.; Moon, J.H.; Tsushida, T.; Nagao, A.; Terao, J. Inhibitory effect of quercetin metabolites and their related derivatives on copper ion-induced lipid peroxidation in human low-density lipoprotein. Arch. Biochem. Biophys. 1999, 372, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Ruiz, M.E.; Guarner-Lans, V.; Cano-Martinez, A.; Diaz-Diaz, E.; Manzano-Pech, L.; Gamas-Magana, A.; Castrejon-Tellez, V.; Tapia-Cortina, C.; Perez-Torres, I. Resveratrol and Quercetin Administration Improves Antioxidant DEFENSES and reduces Fatty Liver in Metabolic Syndrome Rats. Molecules 2019, 24, 1297. [Google Scholar] [CrossRef] [Green Version]
- Korkina, L.G.; Afanas’ev, I.B. Antioxidant and chelating properties of flavonoids. Adv. Pharm. 1997, 38, 151–163. [Google Scholar]
- de Souza, R.F.; De Giovani, W.F. Antioxidant properties of complexes of flavonoids with metal ions. Redox Rep. 2004, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Ferrali, M.; Signorini, C.; Caciotti, B.; Sugherini, L.; Ciccoli, L.; Giachetti, D.; Comporti, M. Protection against oxidative damage of erythrocyte membrane by the flavonoid quercetin and its relation to iron chelating activity. FEBS Lett. 1997, 416, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.C. The effects of stressful life events on depression. Annu. Rev. Psychol. 1997, 48, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Ng, F.; Berk, M.; Dean, O.; Bush, A.I. Oxidative stress in psychiatric disorders: Evidence base and therapeutic implications. Int. J. Neuropsychopharmacol. 2008, 11, 851–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chwastiak, L.A.; Von Korff, M. Disability in depression and back pain: Evaluation of the World Health Organization Disability Assessment Schedule (WHO DAS II) in a primary care setting. J. Clin. Epidemiol. 2003, 56, 507–514. [Google Scholar] [CrossRef]
- Chand, S.P.; Arif, H. Depression; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- McCoy, C.R.; Jackson, N.L.; Day, J.; Clinton, S.M. Genetic predisposition to high anxiety- and depression-like behavior coincides with diminished DNA methylation in the adult rat amygdala. Behav. Brain Res. 2017, 320, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Kohli, M.A.; Lucae, S.; Saemann, P.G.; Schmidt, M.V.; Demirkan, A.; Hek, K.; Czamara, D.; Alexander, M.; Salyakina, D.; Ripke, S.; et al. The neuronal transporter gene SLC6A15 confers risk to major depression. Neuron 2011, 70, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Zanier-Gomes, P.H.; de Abreu Silva, T.E.; Zanetti, G.C.; Benati, E.R.; Pinheiro, N.M.; Murta, B.M.; Crema, V.O. Depressive behavior induced by social isolation of predisposed female rats. Physiol. Behav. 2015, 151, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Hirschfeld, R.M. History and evolution of the monoamine hypothesis of depression. J. Clin. Psychiatry 2000, 61 (Suppl. S6), 4–6. [Google Scholar]
- Nutt, D.J. Relationship of neurotransmitters to the symptoms of major depressive disorder. J. Clin. Psychiatry 2008, 69 (Suppl. SE1), 4–7. [Google Scholar] [PubMed]
- Gorwood, P. Neurobiological mechanisms of anhedonia. Dialogues Clin. Neurosci. 2008, 10, 291–299. [Google Scholar]
- Newberg, A.; Amsterdam, J.; Shults, J. Dopamine transporter density may be associated with the depressed affect in healthy subjects. Nucl. Med. Commun. 2007, 28, 3–6. [Google Scholar] [CrossRef]
- Brietzke, E.; Stertz, L.; Fernandes, B.S.; Kauer-Sant’anna, M.; Mascarenhas, M.; Escosteguy Vargas, A.; Chies, J.A.; Kapczinski, F. Comparison of cytokine levels in depressed, manic and euthymic patients with bipolar disorder. J. Affect. Disord. 2009, 116, 214–217. [Google Scholar] [CrossRef]
- Chen, Y.; Ouyang, J.; Liu, S.; Zhang, S.; Chen, P.; Jiang, T. The Role of Cytokines in the Peripheral Blood of Major Depressive Patients. Clin. Lab. 2017, 63, 1207–1212. [Google Scholar] [CrossRef]
- Fernandes, B.S.; Berk, M.; Turck, C.W.; Steiner, J.; Goncalves, C.A. Decreased peripheral brain-derived neurotrophic factor levels are a biomarker of disease activity in major psychiatric disorders: A comparative meta-analysis. Mol. Psychiatry 2014, 19, 750–751. [Google Scholar] [CrossRef]
- Werner, F.M.; Covenas, R. Classical neurotransmitters and neuropeptides involved in major depression: A review. Int. J. Neurosci. 2010, 120, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Grippo, A.J.; Johnson, A.K. Stress, depression and cardiovascular dysregulation: A review of neurobiological mechanisms and the integration of research from preclinical disease models. Stress 2009, 12, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The interplay between inflammation, oxidative stress, DNA damage, DNA repair and mitochondrial dysfunction in depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef]
- Bishop, A.; Anderson, J.E. NO signaling in the CNS: From the physiological to the pathological. Toxicology 2005, 208, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Drevets, W.C.; Savitz, J.; Trimble, M. The subgenual anterior cingulate cortex in mood disorders. CNS Spectr. 2008, 13, 663–681. [Google Scholar] [CrossRef]
- Sen, S.; Duman, R.; Sanacora, G. Serum brain-derived neurotrophic factor, depression, and antidepressant medications: Meta-analyses and implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafo, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological links between depression and AD: The role of TGF-beta1 signaling as a new pharmacological target. Pharm. Res. 2018, 130, 374–384. [Google Scholar] [CrossRef]
- Myint, A.M.; Leonard, B.E.; Steinbusch, H.W.; Kim, Y.K. Th1, Th2, and Th3 cytokine alterations in major depression. J. Affect. Disord. 2005, 88, 167–173. [Google Scholar] [CrossRef]
- Musil, R.; Schwarz, M.J.; Riedel, M.; Dehning, S.; Cerovecki, A.; Spellmann, I.; Arolt, V.; Muller, N. Elevated macrophage migration inhibitory factor and decreased transforming growth factor-beta levels in major depression—No influence of celecoxib treatment. J. Affect. Disord. 2011, 134, 217–225. [Google Scholar] [CrossRef]
- Corbineau, S.; Breton, M.; Mialet-Perez, J.; Costemale-Lacoste, J.F. Major depression and heart failure: Interest of monoamine oxidase inhibitors. Int. J. Cardiol. 2017, 247, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Medrihan, L.; Sagi, Y.; Inde, Z.; Krupa, O.; Daniels, C.; Peyrache, A.; Greengard, P. Initiation of Behavioral Response to Antidepressants by Cholecystokinin Neurons of the Dentate Gyrus. Neuron 2017, 95, 564–576 e564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, M.M.; Waseem, M. Tricyclic Antidepressant Toxicity; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Bradley, A.J.; Lenox-Smith, A.J. Does adding noradrenaline reuptake inhibition to selective serotonin reuptake inhibition improve efficacy in patients with depression? A systematic review of meta-analyses and large randomised pragmatic trials. J. Psychopharmacol. 2013, 27, 740–758. [Google Scholar] [CrossRef]
- Kumar, U.; Medel-Matus, J.S.; Redwine, H.M.; Shin, D.; Hensler, J.G.; Sankar, R.; Mazarati, A. Effects of selective serotonin and norepinephrine reuptake inhibitors on depressive- and impulsive-like behaviors and on monoamine transmission in experimental temporal lobe epilepsy. Epilepsia 2016, 57, 506–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.P.; Fu, T.T.; Zhang, X.Y.; Yang, F.Y.; Zheng, G.X.; Xue, W.W.; Chen, Y.Z.; Yao, X.J.; Zhu, F. Differentiating physicochemical properties between NDRIs and sNRIs clinically important for the treatment of ADHD. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2766–2777. [Google Scholar] [CrossRef] [PubMed]
- Moret, C.; Isaac, M.; Briley, M. Problems associated with long-term treatment with selective serotonin reuptake inhibitors. J. Psychopharmacol. 2009, 23, 967–974. [Google Scholar] [CrossRef]
- Czéh, B.; Fuchs, E.; Wiborg, O.; Simon, M. Animal models of major depression and their clinical implications. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 293–310. [Google Scholar] [CrossRef]
- Duman, C.H. Models of depression. Vitam. Horm. 2010, 82, 1–21. [Google Scholar]
- Deussing, J.M. Animal models of depression. Drug Discov. Today Dis. Models 2006, 3, 375–383. [Google Scholar] [CrossRef]
- Cairncross, K.; Cox, B.; Forster, C.; Wren, A.F. A new model for the detection of antidepressant drugs: Olfactory bulbectomy in the rat compared with existing models. J. Pharmacol. Methods 1978, 1, 131–143. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef]
- Lumia, A.R.; Teicher, M.H.; Salchli, F.; Ayers, E.; Possidente, B. Olfactory Bulbectomy as a Model for Agitated Hyposerotonergic Depression. Brain Res. 1992, 587, 181–185. [Google Scholar] [CrossRef]
- O’Neil, M.F.; Moore, N.A. Animal models of depression: Are there any? Hum. Psychopharmacol. Clin. Exp. 2003, 18, 239–254. [Google Scholar] [CrossRef]
- Barr, A.M.; Markou, A. Psychostimulant withdrawal as an inducing condition in animal models of depression. Neurosci. Biobehav. Rev. 2005, 29, 675–706. [Google Scholar] [CrossRef] [PubMed]
- Bucan, M.; Abel, T. The mouse: Genetics meets behaviour. Nat. Rev. Genet. 2002, 3, 114–123. [Google Scholar] [CrossRef]
- Scott, P.A.; Cierpial, M.A.; Kilts, C.D.; Weiss, J.M. Susceptibility and resistance of rats to stress-induced decreases in swim-test activity: A selective breeding study. Brain Res. 1996, 725, 217–230. [Google Scholar] [CrossRef]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav. Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef]
- Blanchard, R.J.; Hebert, M.; Sakai, R.R.; McKittrick, C.; Henrie, A.; Yudko, E.; McEwen, B.S.; Blanchard, D.C. Chronic social stress: Changes in behavioral and physiological indices of emotion. Aggress. Behav. J. Int. Soc. Res. Aggress. 1998, 24, 307–321. [Google Scholar] [CrossRef]
- Katz, R.J.; Roth, K.A.; Carroll, B.J. Acute and chronic stress effects on open field activity in the rat: Implications for a model of depression. Neurosci. Biobehav. Rev. 1981, 5, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef]
- Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; An, S.-C.; Ma, X.-M. Dendritic spines in depression: What we learned from animal models. Neural. Plast. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.; Carvalho, M.; Padovan, C. Both serotonergic and noradrenergic systems modulate the development of tolerance to chronic stress in rats with lesions of the serotonergic neurons of the median raphe nucleus. Behav. Brain Res. 2019, 357, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, T.J.; McCausland, H.C.; Morris, H.D.; Padmanaban, V.; Cameron, H.A. Stress and loss of adult neurogenesis differentially reduce hippocampal volume. Biol. Psychiatry 2017, 82, 914–923. [Google Scholar] [CrossRef]
- Marsden, W. Synaptic plasticity in depression: Molecular, cellular and functional correlates. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 43, 168–184. [Google Scholar] [CrossRef]
- Ménard, C.; Pfau, M.L.; Hodes, G.E.; Russo, S.J. Immune and neuroendocrine mechanisms of stress vulnerability and resilience. Neuropsychopharmacology 2017, 42, 62–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Timberlake, M.A., II; Prall, K.; Dwivedi, Y. The recent progress in animal models of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Arad, M.; Terrillion, C.E.; Piantadosi, S.C.; Gould, T.D. The mouse forced swim test. J. Vis. Exp. 2012, e3638. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, S.; Hara, A.; Sakakibara, H.; Kawabata, K.; Tokumura, A.; Ishisaka, A.; Kawai, Y.; Terao, J. Effect of quercetin and glucuronide metabolites on the monoamine oxidase-A reaction in mouse brain mitochondria. Nutrition 2011, 27, 847–852. [Google Scholar] [CrossRef]
- Kumar, A.; Garg, R.; Prakash, A.K. Effect of St. John’s Wort (Hypericum perforatum) treatment on restraint stress-induced behavioral and biochemical alteration in mice. BMC Complementary Altern. Med. 2010, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Liaquat, L.; Batool, Z.; Sadir, S.; Rafiq, S.; Shahzad, S.; Perveen, T.; Haider, S. Naringenin-induced enhanced antioxidant defence system meliorates cholinergic neurotransmission and consolidates memory in male rats. Life Sci. 2018, 194, 213–223. [Google Scholar] [CrossRef]
- Jeon, S.W.; Kim, Y.K. Inflammation-induced depression: Its pathophysiology and therapeutic implications. J. Neuroimmunol. 2017, 313, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Wium-Andersen, M.K.; Orsted, D.D.; Nielsen, S.F.; Nordestgaard, B.G. Elevated C-reactive protein levels, psychological distress, and depression in 73,131 individuals. JAMA Psychiatry 2013, 70, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, G.M.; Pearson, R.M.; Zammit, S.; Lewis, G.; Jones, P.B. Association of serum interleukin 6 and C-reactive protein in childhood with depression and psychosis in young adult life: A population-based longitudinal study. JAMA Psychiatry 2014, 71, 1121–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, K.I.; Miller, D.L.; Roecklein, K.A. The Aging Hippocampus: Interactions between Exercise, Depression, and BDNF. Neuroscientist 2012, 18, 82–97. [Google Scholar] [CrossRef]
- Halder, S.; Kar, R.; Mehta, A.K.; Bhattacharya, S.K.; Mediratta, P.K.; Banerjee, B.D. Quercetin Modulates the Effects of Chromium Exposure on Learning, Memory and Antioxidant Enzyme Activity in F1 Generation Mice. Biol. Trace Elem. Res. 2016, 171, 391–398. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors Names | Name of the Ethics Committee or Institutional Review Board | Project Identification Code | Date of Approval | Reference |
|---|---|---|---|---|
| Samad N et al. | Institutional ethics and animal care committee | - | - | [32] |
| Singh V. et al. | Institutional Animal Ethics Committee (IAEC) | No. 107/GO/ReBi/S/99/CPCSEA/2017-01 | 27 February 2017 | [33] |
| Khan K. et al. | Institutional Animal Ethics Committee (IAEC) | No. 1314. | - | [34] |
| Mehta V. et al. | Institute Animal Ethics Committee in accordance with the guidelines of CPCSEA, India | - | - | [35] |
| Shen, Z. et al. | Institutional Animal Care and Use Committee at Jiaxing University | - | - | [36] |
| Ma Z.X. et al. | General Project of Science and Technology Development Fund of Nanjing Medical University (Item) | No. NMUB2019114 | - | [37] |
| Donoso F. et al. | Ethics Committee of University College Cork | - | - | [38] |
| Sahin T.D. et al. | Animal Research Ethics Committee of Kocaeli University, Turkey | No: KOÜ HADYEK 2/9-2017 | 09/02/2017 | [39] |
| Anggreini P. et al. | Ethical Committee of the Faculty of Veterinary Medicine, Universitas Airlangga | No. 2.KE.007.05.2019. | - | [40] |
| Zhang J. et al. | Institutional Animal Care and Use Committee (IACUC) of University of Science and Technology of China. | - | - | [41] |
| Guan et al. | Medical Ethics Committee of Harbin Medical University | - | - | [42] |
| Fang K. et al. | Animal Care and Use committee at Anhui Medical University | - | - | [43] |
| Sadighparvar S. et al. | Animal Care and Use Committee at the Urmia University of Medical Sciences, Urmia, Iran | IR.UMSU.REC.1396.316 | - | [44] |
| Wang G. et al. | Animal Care Committee of Jinan University | No. 2,019,671 | - | [45] |
| Merzoug S et al. | NIH revised Guidelines for the Care and Use of Laboratory Animals | No. 80-23 | - | [46] |
| Demir E.A. et al. | Local Ethics and Animal Care Committee of Necmettin Erbakan University | 2013-053 | - | [47] |
| Holzmann et al. | Institutional Ethics Committee of UNIVALI, Itajaí, SC, Brazil | No.021/13 | - | [48] |
| Rinwa P. et al. | Institutional Animal Ethics Committee (IAEC) of the Panjab University | IAEC/282/UIPS/39 | 30/8/12 | [49] |
| Models | Compounds | Quercetin Dose | Treatments | Results | Ref. |
|---|---|---|---|---|---|
| Chronic Stress | |||||
| Depressive-like behaviors model induced by immobilization-stressed in Albino Wistar mice | Quercetin | 20 mg/kg | Daily intraperitoneal injections for 14 days | Quercetin induced antidepressant and anxiolytic effects through the reduction of the stress-induced lipid peroxidation and increased antioxidant enzymes such SOD, CAT and GPx. Moreover, quercetin significantly increased Ach and reduced 5-HIAA levels in the brain of stressed mice, ameliorating serotonergic and cholinergic functions. | [32] |
| Depressive-like behaviors model induced by unpredictable mild chronic stress in Swiss Albino mice | Quercetin or Quercetin 4′-O-glucoside | 10 and 20 mg/kg | Daily orally administration for 7 or 21 days | Quercetin and quercetin 4′-O-glucoside exerted their antidepressive activity through decreasing immobility time in the FST and improve locomotor activity. Moreover, in a mouse model of unpredictable mild chronic stress-induced depressive behavior, quercetin 4′-O-glucoside exhibited anti-depressive effects, increasing 5-HT levels in the brain and reducing ROS levels and MAO-A activity. | [33] |
| Depressive-like behavior model induced by unpredictable mild chronic stress in Swiss Albino mice | Quercetin or fluoxetine | 25 mg/kg | Daily orally administration for 28 days | Treatment with quercetin significantly increased SOD, GSH, catalase and 5-HT levels as well as reversed glutamate and increase pro-inflammatory cytokines TNF-α and IL-6 levels induced by unpredictable chronic mild stress. | [34] |
| Depression-like behaviors model induced by chronic unpredicted stress in Swiss Albino mice | Quercetin | 30 mg/kg | Daily orally ad-ministration for 21 days | Quercetin reduced anxiety in animals, mitigated anhedonia and ameliorated the short- and long-term memory deficits. It also decreased levels of total thiol, TBARS and NO; conversely, it increased CAT levels. Additionally, treatment reduced hippocampal gene expression levels for TNF-α, IL-6, IL-1β, COX-2, and protected neurons from stress-mediated damage. | [35] |
| Depressive-like behavior model induced by unpredictable mild chronic stress in ICR mice | Quercetin or fluoxetine | 15 and 30 mg/kg | Daily administration via gavage for 21 days | Quercetin, at both doses, ameliorated the depressive-like behaviors, restoring stress-induced weight loss, improving locomotor activity, decreasing immobility time and improving anhedonia. It also could induce maturation and survival of neural progenitor cells hippocampus. Moreover, this compound increased levels of FoxG1, p-CREB and BDNF, thus it could induce antidepressive effects via FoxG1/CREB/BDNF signaling pathway. | [37] |
| Early life stress-induced depressive like behavior in pups Sprague Dawley mice | Quercetin | 20 mg/kg | Dietary intervention for 56 days | Quercetin dietary raised the levels of plasma BDNF. It also improved HPA axis dysregulation, reducing corticosterone plasma levels. Quercetin also prevented depressive-like behaviors induced by maternal separation. | [38] |
| Depressive-like behavior induced by unpredictable chronic mild stress in Wistar albino rats | Quercetion or resveratrol | 30 mg/kg | Intraperitoneally injection for 35 days along with stress induction | Quercetin and resveratrol reduced immobility time and increased sucrose consumption, exerting anti-depressive effects. Both flavonoids enhanced antioxidants defense (SOD, CAT, and GSH) and reduced MDA levels. These flavonoids also reduced NF-κB and pro-inflammatory cytokine (IL-6, TNF-α, and IL-1β) expression levels. They also restored the HPA axis decreasing serum corticosterone levels. | [39] |
| Psychological stress model induced by predatory stress in ICR mice | Quercetin | 50 mg/kg | Intraperitoneally injection for 3 days along with stress induction | After 3 days of predatory stress exposure, quercetin reduced the innate fear induced by stress. Also, treatment improved locomotory immobility time and spatial working memory and voluntary exploration of mice as demonstrated by the Y maze paradigm. | [40] |
| Depressive-like behaviors induced by chronic social defeat stress in C57BL/6N mice | Quercetin | 0.5 and 2 g/kg | Oral administration via the diet | A quercetin-enriched diet (2 g/kg) for 14 days before exposure to stress ameliorated depressive-like behaviors and increased neuronal activity in the medial prefrontal cortex and hippocampus. Moreover, quercetin-enriched diets inhibited the expression of genes related to microglia and astrocyte activation in both the medial prefrontal cortex and hippocampus. | [41] |
| Model of depression induced by chronic unpredictable mild stress in rats | Quercetin | 10 and 50 mg/kg | Daily administration via gavage for 8 weeks | Quercetin (50 mg/kg) treatment increased the sucrose preference of depressed rats and decreased pro-inflammatory cytokines serum levels. Moreover, it regulated serum levels of iron, copper, calcium, magnesium, zinc, selenium, and cobalt. | [42] |
| Acute stress | |||||
| Depressive-like behaviors model induced by intraperitoneal injection of LPS in Sprague-Dawley rats | Quercetin, ibuprofen or fluoxetine | 40 mg/kg | Daily administration via gavage for 14 days | Quercetin improved ameliorated the impaired LPS-induced neuropsychiatric behaviors including anxiety, depression and memory impairment. Moreover, treatment with quercetin exerted its anti-depressive actions regulating the protein expression of BDNF, p-TrkB/TrkB, TREM1/TREM2, Copine 6, and synapsin-1, both in the hippocampus and prefrontal cortex. | [43] |
| Depressive-like behaviors induced by DMH injections in Wistar rats | Quercetin | 50 mg/kg | Daily orally administration for 84 days | Quercetin reduced the immobility time and enhanced ambulation and rearing counts, improving the DMH-induced depressive-like behaviors. Quercetin exerted antidepressive effects decreasing inflammatory cytokines, such as IL-1β and TNF-α; which in turn upregulated BDNF/TrKB/β-catenin signaling. | [44] |
| Depressive-like behaviors induced by estrogen receptor α deficiency in female mice | Quercetin | 100 mg/kg | Daily orally administration for 70 days | Treatment ameliorated the depressive responses in estrogen receptor α deficiency-female mice. It also increased the number of hippocampal neurons and reduced apoptosis. Moreover, quercetin increased the expression levels of BDNF and of its phosphorylation downstream targets such as TrkB, Akt, and ERK1/2, showing that it could exert antidepressive effects modulating this pathway. | [45] |
| Anxiety-depressive-like behavior model induced by intraperitoneal injection of Adriamycin in Wistar rats | Quercetin | 60 mg/kg | Single intraperitoneal injections at 24, 5, and 1 h | Quercetin ameliorated the anxiety-depressive-like behavior, restoring the plasma corticosterone concentration and reducing oxidative stress. In detail, quercetin restored the GSH, GST, and MDA levels. Moreover, quercetin treatment reversed leucopenia and attenuated lymphopenia and monocytosis. | [46] |
| Depressive-like behavior model induced by intraperitoneal injection of streptozotocin in Wistar Albino rats | Quercetin | 50 and 100 mg/kg | Daily intraperitoneal injection for 21 days | Quercetin treatment at a dose of 50 mg/kg ameliorated depressive-like behaviors; conversely, the higher dose did not induce antidepressant effects. Quercetin did not change the plasmatic level of adrenocorticotropic hormone and corticosterone, showing that this flavonoid exerted antidepressive effects independent of the HPA axis. | [47] |
| Depression induced by olfactory bulbectomy in Wistar rats | Quercetin, fluoxetine L-arginine, methylene blue, NMDA, 7-nitroindazole, sildenafil | 25 mg/kg | Daily orally administration for 14 days | Quercetin exerted antidepressant effects reducing lipid hydroperoxide levels in the hippocampus. These antidepressive effects were reversed following treatment with NMDA, L-arginine, and sildenafil. The administration of methylene blue and 7-nitroindazole, in combination with quercetin, showed a quercetin-like antidepressant effect, highlighting that anti-depressive effects of quercetin could be linked to inhibition of NMDA receptors and NO synthesis. | [48] |
| Depression induced by olfactory bulbectomy in Wistar rats | Quercetin and/or minocycline | 20, 40 and 80 mg/kg | Daily orally administration for 14 days | Quercetin ameliorated behavioral alterations induced by olfactory bulbectomy, restoring oxidative/nitrosative stress markers (such as nitrite, MDA, GSH, SOD, and CAT). It also reduced neuroinflammation reducing TNF-α and IL-6 levels in both the cerebral cortex and hippocampus. Quercetin also reduced caspase-3 activity. Quercetin could exert its anti-depressive effects by reducing microglial activation. | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvestro, S.; Bramanti, P.; Mazzon, E. Role of Quercetin in Depressive-Like Behaviors: Findings from Animal Models. Appl. Sci. 2021, 11, 7116. https://doi.org/10.3390/app11157116
Silvestro S, Bramanti P, Mazzon E. Role of Quercetin in Depressive-Like Behaviors: Findings from Animal Models. Applied Sciences. 2021; 11(15):7116. https://doi.org/10.3390/app11157116
Chicago/Turabian StyleSilvestro, Serena, Placido Bramanti, and Emanuela Mazzon. 2021. "Role of Quercetin in Depressive-Like Behaviors: Findings from Animal Models" Applied Sciences 11, no. 15: 7116. https://doi.org/10.3390/app11157116