
New Geometric Morphometric Insights in Digital Taphonomy: Analyses into the Sexual Dimorphism of Felids through Their Tooth Pits


 ,
,
Abstract
:Featured Application
Abstract
1. Introduction
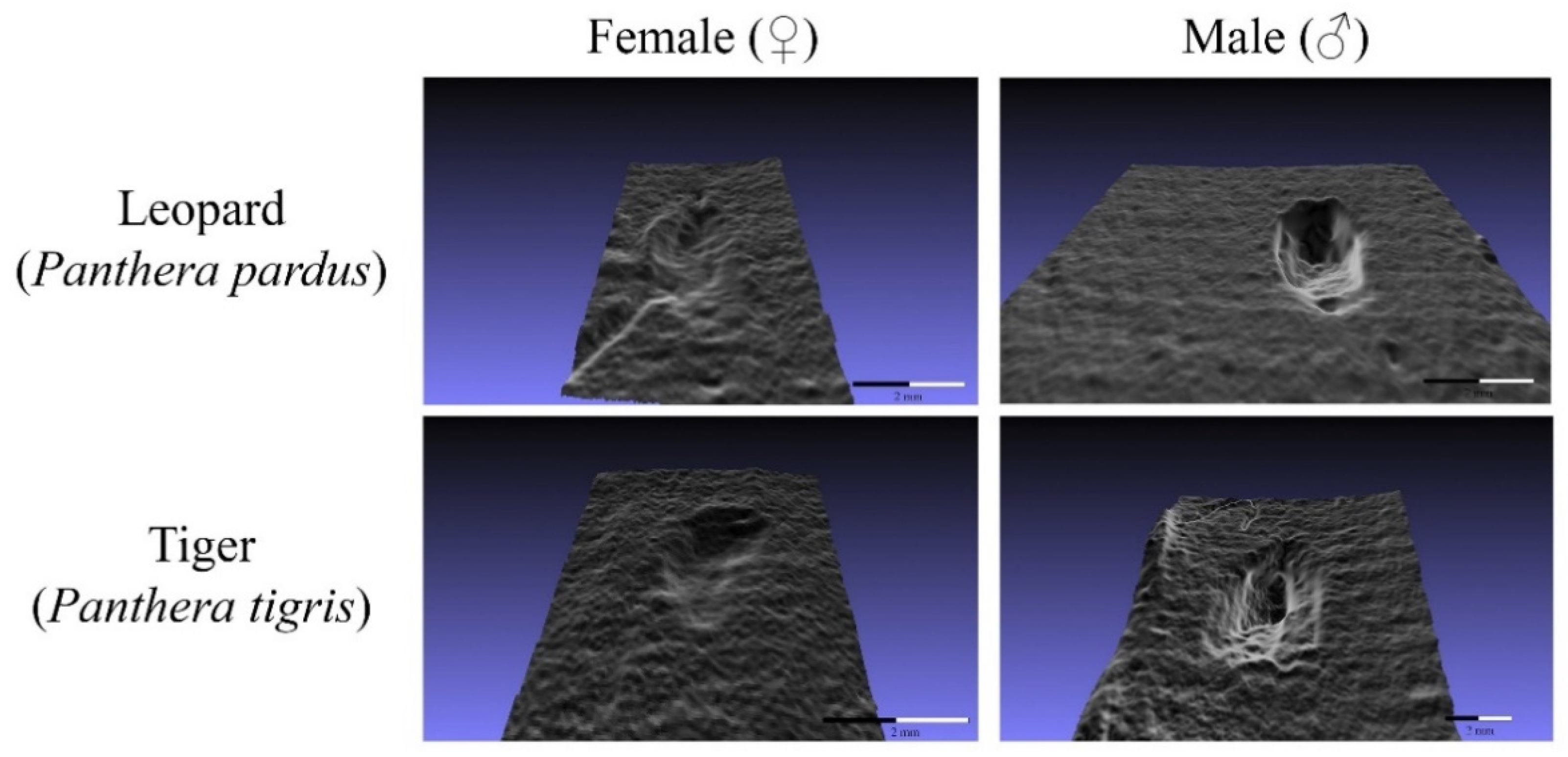
2. Materials and Methods
2.1. Sample
2.2. Methods
2.3. Data Analysis
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aramendi, J.; Maté-González, M.A.; Yravedra, J.; Cruz-Ortega, M.; Arriaza, M.A.; González- Aguilera, D.; Baquedano, E.; Domínguez-Rodrigo, M. Discerning carnivore agency through the three-dimensional study of tooth pits: Revisiting crocodile feeding behaviour at FLK-Zinj and FLK NN3 (Olduvai Gorge, Tanzania). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 488, 93–102. [Google Scholar] [CrossRef]
- Aramendi, J.; Arriaza, M.A.; Yravedra, J.; Maté-González, M.A.; Ortega, M.C.; Courteany, L.; González-Aguilera, D.; Gidna, A.; Mabulla, A.; Baquedano, E.; et al. Who ate OH80 (Olduvai Gorge, Tanzania)? A geometric-morphometric analysis of surface bone modifications of a Paranthropus boisei skeleton. Quat. Int. 2019, 517, 118–130. [Google Scholar] [CrossRef]
- Yravedra, J.; Maté-González, M.A.; Courtenay, L.A.; González-Aguilera, D.; Fernández-Fernández, M. The use of canid tooth marks on bone for the identification of livestock predation. Sci. Rep. 2019, 9, 16301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaggio, M.M.; Wilder, J. Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. JASR 2001, 28, 465–470. [Google Scholar] [CrossRef]
- Delaney-Rivera, C.; Plummer, T.W.; Hodgson, J.A.; Forrest, F.; Hertel, F.; Oliver, J.S. Pits and pitfalls: Taxonomic variability and patterning in tooth mark dimensions. JASR 2009, 36, 2597–2608. [Google Scholar] [CrossRef]
- Andres, M.; Gidna, A.; Yravedra, J.; Dominguez-Rodrigo, M. A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol. Anthropol. Sci. 2012, 4, 209–219. [Google Scholar] [CrossRef]
- Mackay, A. Mitigating Cattle Losses Caused by Wild Predators in British Columbia. A Field Guide for Ranchers; Ministry of Agriculture & Lands, Agriculture et Agroalimentaire British Columbia Canada, 2005; Available online: https://www.cattlemen.bc.ca/docs/mitigating_cattle_losses_a_field_guide_for_ranchers_smaller_file.pdf (accessed on 25 August 2021).
- Carmichael, E.J. Targeted Predator Control. A Guide for Livestock Producers on Option Available for Targeted Predator Control. B. C. Agricultural Research & Development Corporation Wild Predator Loss Prevention Pilot Project. 2011. Available online: https://www.researchgate.net/deref/https%3A%2F%2Fwww.bcac.bc.ca%2Fsites%2Fbcac.localhost%2Ffiles%2FTargeted%2520Predator%2520Control.pdf (accessed on 25 August 2021).
- Caniglia, R.; Fabbri, E.; Mastrogiuseppe, L.; Randi, E. Who is who? Identification of livestock predators using forensic genetic approaches. Forensic Sci. Int. Genet. 2013, 7, 397–404. [Google Scholar] [CrossRef]
- Toledo, V.; Ortega-Ojeda, F.; Fonseca, G.; García-Ruiz, C.; Pérez-Lloret, P. Analysis of tooth mark patterns on bone remains caused by wolves (Canis lupus) and domestic dogs (Canis lupus familiaris) for taxonomic identification: A scoping review focused on their value as a forensic tool. Appl. Anim. Behav. Sci. 2021, 240, 105356. [Google Scholar] [CrossRef]
- Yravedra, J.; García-Vargas, E.; Maté-González, M.A.; Aramendi, J.; Palomeque-González, J.F.; Vallés-Iriso, J.; Matesanz-Vicente, J.; González-Aguilera, D.; Domínguez-Rodrigo, M. The use of Micro-Photogrammetry and Geometric Morphometrics for identifying carnivore agency in bone assemblages. JASR 2017, 14, 106–115. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution in Morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Slice, D.E. Modern Morphometrics. In Modern Morphometrics in Physical Anthropology; Slice, D.E., Ed.; Kluwer Press: New York, NY, USA, 2005. [Google Scholar]
- Bruner, E. Geometric morphometrics and paleoneurology: Brain shape evolution in the genus Homo. J. Hum. Evol. 2004, 47, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Cobb, S.N.; O’Higgins, P. Hominins do not share a common postnatal facial ontoge-netic shape trajectory. J. Exp. Zool. 2004, 302B, 302–321. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.; Collard, M. A geometric morphometrics-based assessment of bladeshape differences among Paleoindian projectile point types from western NorthAmerica. J. Archaeol. Sci. 2010, 37, 350–359. [Google Scholar] [CrossRef]
- Lycett, A.J.; von Cramon-Taubadel, N.; Gowlett, J.A. A comparative 3D geometricmorphometric analysis of Victoria west cores: Implications for the origins of Levalloistechnology. J. Archaeol. Sci. 2010, 37, 1110–1117. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Huguet, R.; Aramendi, J.; Maté-González, M.A.; González-Aguilera, D.; Arriaza, M.C. Combining machine learning algorithms and geometric morphometrics: A study of carnivore tooth marks. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 522, 28–39. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Maté-González, M.A.; Vázquez-Rodríguez, J.M.; Fernández-Fernández, M.; González-Aguilera, D. The effects of prey size on carnivore tooth mark morphologies on bone; the case study of Canis lupus signatus. Hist. Biol. 2020, 1–13. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Herranz-Rodrigo, D.; Huguet, R.; Maté-Gonzalez, M.A.; Gonzalez-Aguilera, D.; Yravedra, J. Obtaining new resolutions in carnivore tooth pit morphological analyses: A methodological update for digital taphonomy. PLoS ONE 2020, 15, e0240328. [Google Scholar] [CrossRef] [PubMed]
- Courtenay, L.A.; Herranz-Rodrigo, D.; González-Aguilera, D.; Yravedra, J. Developments in data science solutions for carnivore tooth pit classification. Sci. Rep. 2021, 11, 10209. [Google Scholar] [CrossRef] [PubMed]
- Courtenay, L.A.; Herranz-Rodrigo, D.; Yravedra, J.; Vázquez-Rodríguez, J.; Huguet, R.; Barja, I.; Maté-González, M.A.; Fernández, M.; González-Aguilera, D. Effects of captivity on carnivore mastication implications in ecological studies of both the past and present. Animals 2021, 11, 2323. [Google Scholar] [CrossRef]
- Gidna, A.; Yravedra, J.; Domínguez-Rodrigo, M. A cautionary note on the use of captive carnivores to model wild predator behavior: A comparison of bone modification patterns on long bones by captive and wild lions. JASR 2013, 40, 1903–1910. [Google Scholar] [CrossRef]
- Sala, N.; Arsuaga, J.L.; Haynes, G. Taphonomic comparison of bone modifications caused by wild and captive wolves (Canis lupus). Quat. Int. 2014, 330, 126–135. [Google Scholar] [CrossRef]
- Kaufmann, C.A.; Rafuse, D.; González, M.E.; Álvarez, M.C.; Massigoge, A.; Scheifler, N.A.; Gutiérrez, M.A. Carcass utilization and bone modifications on guanaco killed by puma in northern Patagonia, Argentina. Quat. Int. 2018, 466 Pt B, 165–177. [Google Scholar] [CrossRef]
- Toledo González, V.; Ortega Ojeda, F.; Fonseca, G.M.; García-Ruiz, C.; Navarro Cáceres, P.; Pérez-Lloret, P.; Marín García, M.d.P. A Morphological and Morphometric Dental Analysis as a Forensic Tool to Identify the Iberian Wolf (Canis Lupus Signatus). Animals 2020, 10, 975. [Google Scholar] [CrossRef] [PubMed]
- Sunquist, M.; Sunquist, F. Wild Cats of the World; Chicago University Press: Chicago, IL, USA, 2002. [Google Scholar]
- Mazák, J.H. On the sexual dimorphism in the skull of the tiger (Panthera tigris). Mam. Biol. 2004, 69, 392–400. [Google Scholar] [CrossRef]
- Farhadinia, M.S.; Kaboli, M.; Karami, M.; Farahmand, H. Patterns of sexual dimorphism in the Persian Leopard (Panthera pardus saxicolor) and implications for sex differentiation. Zool. Middle East 2014, 60, 195–207. [Google Scholar] [CrossRef]
- Pocock, R.I. Panthera pardus (Linnaeus). The Leopard or Panther. The Fauna of British India, including Ceylon and Burma. Mammalia 1930, 1, 222–231. [Google Scholar]
- Castelló, J.R. Felids and Hyenas of the World, Princeton Field Guides; Princeton University Press: Princeton, NJ, USA, 2020. [Google Scholar]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/ SSC Cat Specialist Group. Cat News Spec. Issue 2017, 11, 80. [Google Scholar]
- Meiri, S.; Dayan, T.; Simberloff, D. Variability and sexual size dimorphism in carnivores: Testing the niche variation hypothesis. Ecology 2005, 86, 1432–1440. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.K. Shape statistics: Procrustes superimpositions and tangent spaces. J. Classif. 1999, 16, 197–223. [Google Scholar] [CrossRef]
- Slice, D.E. Landmark coordinates aligned by Procrustes analysis do not lie in Kendall’s shape space. Syst. Biol. 2001, 50, 141–149. [Google Scholar] [CrossRef]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: New York, NY, USA, 1991. [Google Scholar]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis; John Wiley and Sons: New York, NY, USA, 1998. [Google Scholar]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st century. Hystrix 2013, 24, 7–14. [Google Scholar] [CrossRef]
- Bookstein, F.L. Principal warps: Thin plate spline and the decomposition of deformations. Trans. Pattern Anal. Mach. Intell. 1989, 11, 567–585. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for Behavioural Sciences; Routledge: New York, NY, USA, 1988. [Google Scholar]
- Schurimann, D.L. A comparison of the two one-sided test procedure and the power approach for assessing the equivalence of average biovariability. J. Pharm. Biopharm. 1987, 15, 657–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuen, K.K.; Dixon, W.J. The approximate behaviour and performance of the two-sample trimmed t. Biometrika 1973, 60, 369–374. [Google Scholar] [CrossRef]
- Yuen, K.K. The two-sample trimmed t for unequal population variances. Biometrika 1974, 61, 165–170. [Google Scholar] [CrossRef]
- Benjamin, D.J.; Berger, J.O. Three recommendations for improving the use of p-values. Am. Stat. 2019, 73 (Suppl. 1), 186–191. [Google Scholar] [CrossRef] [Green Version]
- Colquhoun, D. The False Positive Risk: A proposal concerning what to do about p-values. Am. Stat. 2019, 73 (Suppl. 1), 192–201. [Google Scholar] [CrossRef] [Green Version]
- Arriaza, M.C.; Yravedra, J.; Domínguez-Rodrigo, M.; Maté-González, M.A.; García-Vargas, E.; Palomeque, J.F.; Aramendi, J.; González-Aguilera, D.; Baquedano, E. On applications of micro-photogrammetry and geometric morphometrics to studies of tooth mark morphology: The modern Olduvai Carnivore Site (Tanzania). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 488, 103–112. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups Compared | F | Res.2 | p-Value | FPR |
|---|---|---|---|---|
| Tiger vs. Leopard | 2.7 | 0.013 | 0.014 | 14.0% |
| Tiger (M vs. F) | 1.5 | 0.014 | 0.183 | 45.7% |
| Leopard (M vs. F) | 3.0 | 0.024 | 0.045 | 27.5% |
| Groups Compared | p-Value | FPR |
|---|---|---|
| Tiger vs. Leopard | 0.042 1 | 26.6% |
| Tiger (M vs. F) | 0.003 | 4.5% |
| Leopard (M vs. F) | 0.009 | 10.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herranz-Rodrigo, D.; Tardáguila-Giacomozzi, S.J.; Courtenay, L.A.; Rodríguez-Alba, J.-J.; Garrucho, A.; Recuero, J.; Yravedra, J. New Geometric Morphometric Insights in Digital Taphonomy: Analyses into the Sexual Dimorphism of Felids through Their Tooth Pits. Appl. Sci. 2021, 11, 7848. https://doi.org/10.3390/app11177848
Herranz-Rodrigo D, Tardáguila-Giacomozzi SJ, Courtenay LA, Rodríguez-Alba J-J, Garrucho A, Recuero J, Yravedra J. New Geometric Morphometric Insights in Digital Taphonomy: Analyses into the Sexual Dimorphism of Felids through Their Tooth Pits. Applied Sciences. 2021; 11(17):7848. https://doi.org/10.3390/app11177848
Chicago/Turabian StyleHerranz-Rodrigo, Darío, Silvia J. Tardáguila-Giacomozzi, Lloyd A. Courtenay, Juan-José Rodríguez-Alba, Antonio Garrucho, Jesús Recuero, and José Yravedra. 2021. "New Geometric Morphometric Insights in Digital Taphonomy: Analyses into the Sexual Dimorphism of Felids through Their Tooth Pits" Applied Sciences 11, no. 17: 7848. https://doi.org/10.3390/app11177848