Abstract
Genotoxicity is the ability of specific substances to cause DNA damage, affecting development, physiology, and reproduction. This is often mediated by induction of oxidative stress. This in vitro study aims to test the ability of two antioxidants, ellagic acid (EA, 100 µM) and curcumin (Cur, 40 µM) to protect zebrafish blood cells from the genotoxic action of benzene (10 µL/mL). Cells were treated for 30, 60, and 90 min with EA or Cur alone and in combination with benzene. The antigenotoxic role of antioxidants was evaluated in terms of cytotoxicity by trypan blue dye, genome stability by RAPD-PCR technique, DNA fragmentation and percentage of apoptotic cells using Comet and Diffusion assay, respectively. The results did not show statistical differences in terms of cell viability, genome stability, DNA damage and apoptosis between cells treated with antioxidants. When zebrafish blood cells were co-incubated with individual antioxidants and benzene, a significant improvement of these parameters was observed in comparison with cells incubated in benzene. Our results suggested that EA and Cur are able to protect zebrafish blood cells against DNA damage and apoptosis caused by mutagenic substance, and laid the foundation for future studies investigating their antigenotoxic potential in DNA oxidative damage therapy.
1. Introduction
Genotoxicity is the capacity of a substance to damage the DNA by chemically modifying the nucleotide sequence or the DNA structure [1]. This can have severe repercussions on cellular functions and survival, and can be transmitted to the offspring when germ cells are damaged [1]. Reactive oxygen species (ROS) are genotoxic molecules, playing a role in several pathologies including cancer [2,3]. ROS are highly unstable molecules due to the presence in their electronic shell of one or more unpaired electrons; hence, they are highly reactive with any cellular components (i.e., lipids, proteins, and DNA) and, when generated in excess, their oxidative power can significantly compromise cellular functions and trigger the apoptosis. Oxidative stress (OS) is due to the breaking of the physiological balance between the production and elimination of ROS, causing pathological conditions.
Several genotoxic substances are present in the environment as pollutants due to their use in industrial production. Exposure to cadmium, a heavy metal present in pigments used for plastic products [4], significantly increases the rate of DNA damage in zebrafish [5], mainly through the generation of OS [6]. Further, in vitro exposure of human sperm cells to nano-contaminant resulted in an increased rate of DNA damage and reduced DNA integrity, along with increased ROS generation [7].
As OS have a genotoxic effect due to ROS-mediated DNA damage, exogenous antioxidants have been investigated for their potential to counteract DNA damage. The phytochemicals contained in plants are known to be able to prevent oxidative DNA damage. In vivo and in vitro studies have demonstrated the antigenotoxic and antioxidant properties of the extract of Ziziphus jujuba fruit, a plant belonging to the family Rhamnaceae, against genotoxins such as methyl methan sulfonate and hydrogen peroxide (H2O2) [8]. Similarly, Croton antisyphiliticus extracts reportedly inhibited the cytogenetic damage induced by genotoxic agents in human lymphocytes [9], while the antioxidant and antigenotoxic potential of a-tocopherol and anthocyanin are demonstrated in vitro and in vivo [10]. Natural sources of these molecules include berries, fruits, vegetables, seeds, and cereals [11,12].
Ellagic acid (EA) and curcumin (Cur) are emerging as natural compounds with antioxidant and antigenotoxic properties [13,14]. EA is a natural polyphenolic compound present in many fruits (such as pomegranate and Goji berries) and it has demonstrated antioxidant properties: it can eliminate two radicals per cycle, it is able to regenerate itself continuously, and its secondary metabolites are scavengers of a large number of radicals as well [15,16,17]. Moreover, an in vitro study showed EA to prevent cancer proliferation in Balb/C mice, by inducing the expression and activation of Bax and caspase-3, involved in the apoptosis, and inhibiting the expression of Bcl-2 and cyclin D1, involved in the regulation of the cell cycle [18]. EA anti-genotoxic power is also proven in zebrafish (Danio rerio) embryos exposed to H2O2 [19] as well as in human sperm exposed to benzene [13].
Cur is the main biologically active component of Curcuma longa (Turmeric). It is a natural polyphenolic compound with antioxidant, antimicrobial, antifungal, anti-inflammatory, and anti-tumor properties [20]. It has been reported to minimize colonization by Candida albicans, a fungus that causes yeast infections [21]. Also, Cur blocks the tumor growth by inhibiting cell cycle progression and angiogenesis, inducing apoptosis and modulating immune responses [22]. Its antioxidant potential was recently reported in vivo in zebrafish embryos exposed to H2O2, as they showed lower lipid peroxidation, ROS generation and apoptotic rates after co-exposure to turmeric leaf extract [23].
The objective of this study was to evaluate the anti-genotoxic power of EA and Cur against benzene, a well-known genotoxic agent [24], on zebrafish blood cells in vitro. Zebrafish was chosen because it is a model widely used in genetic toxicology studies as an excellent bioindicator [25]. Cellular blood cell vitality was evaluated by trypan blue staining, Comet assay and diffusion assay were performed to evaluate the degree of DNA fragmentation and apoptosis, respectively, while RAPD-PCR was conducted to analyze genomic stability.
2. Materials and Methods
2.1. Preparation of Solutions
Chemicals used in this study were supplied by Sigma-Aldrich (St. Louis, MO, USA) and included: benzene (>99.0%, CAS number 71-43-2, 99.8% purity), EA (CAS number 47-66-4, ≥95.0% purity), and Cur (CAS number 458-37-7, ≥94% (curcuminoid content), ≥80% (Curcumin)). EA and Cur powders were dissolved in 0.5% absolute ethanol (Thermo Fisher Scientific Waltham, MA, USA, CAS: 64-17-5).
2.2. In Vitro Study Design
We tested EA and Cur antigenotoxicity on zebrafish blood cells in vitro. Maintenance and experimental procedures were in accordance with the recommendations reported in the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes [26]. Zebrafish (n = 60) obtained from a local source (CARMAR sas, San Giorgio a Cremano, Italy) were bred in a large tank containing ~80 L of water (temperature ≥ 25 °C, pH = 7.6), and respecting the photoperiod of 10 h (h) of darkness and 14 h of light, fed every two days (Tetramin Tropical Flakes, Spectrum Brands, Blacksburg, VA, USA). Adult fish were anesthetized with ethyl 3-aminobenzoate methanesulfonate (E10521, Sigma-Aldrich) and the blood collected by using a heparinized syringe. Zebrafish blood cells obtained from 60 zebrafish were pooled and washed in phosphate buffered saline (PBS) 1X. Samples were split into six different aliquots (10 × 106 cells/mL) according to the study design: 100 μM EA (EA group); 40 μM Cur (Cur group); 100 μM EA plus 10 μL/mL benzene (EA+ BZ group); 40 μM Cur plus 10 μL/mL benzene (Cur + BZ group); positive control (benzene 10 μL/mL) and negative control groups. In the negative control group, ethanol was added at the same volume used to dissolve Cur and EA. Experiments were performed at 30, 60, and 90 exposure minutes (min).
2.3. Cytotoxicity Assay
Zebrafish blood cells were stained with trypan blue (Thermo Fisher Scientific, Waltham, MA, USA) to determine cell viability [27]. Specifically, 10 μL of each sample was mixed with 0.4% trypan blue dye at room temperature for 10 min. Smears on frosted slides were prepared in triplicate. Slides were observed under optical microscope: live cells showed bright centers and dark edges, while dead cells were uniformly blue colored.
2.4. Comet Assay
The alkaline (pH 12.1) Comet assay was performed to investigate the presence of DNA strand breaks in zebrafish blood cells at different exposure conditions, as previously reported [28]. A total of 15 μL of blood cell suspension containing 106 cells/mL were mixed with 65 μL of low melting point agarose (0.7%) and inserted into layers of normal melting agarose (1%) on slides. Slides were prepared in triplicate for each sample. These were incubated in lysis solution (NaCl 2.5 M, Na2EDTA 0.1 M, tris-base 0.4 M, triton-X100 1%, dimethyl sulfoxide-DMSO, 10%, pH 10), followed by incubation in alkaline solution (NaOH 10 N, ethylenediaminetetraacetic acid—EDTA 200 mM, pH 12.1). Slides were then exposed to electrophoresis (25 V, 300 mA, 0.7 V/cm) for 20 min to allow the migration of DNA, followed by fixation in cold methanol and staining with ethidium bromide 1×. A total of 50 cells per slide was analyzed under fluorescence microscope (Nikon Eclipse E-600, 60× magnification, BP 515–560 nm filter, LP 580 nm filter). The images were captured and analyzed by OpenComet software [29]. DNA strand breaks were assessed by analyzing damaged DNA present in the comet tail (% tail DNA).
2.5. Diffusion Assay
Diffusion assay methodology is a modified version of Comet assay, where the cell slides are not subjected to electrophoresis. In this assay, apoptotic cells are differentiated from necrotic cells, as the former show irregular edges, with the presence of highly dispersed DNA into the nuclei, while the latter show larger and not-defined nuclei [30]. Diffusion assay was performed in triplicate. Only apoptotic cell class (class 5) according to degree of DNA diffusion pattern were considered [31].
2.6. Random Amplified Polymorphic DNA-Polymerase Chain Reaction (RAPD-PCR)
The random amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR) involves the use of small primers of arbitrary sequence, which are ideal for amplifying random portions of DNA through PCR [5]. DNA isolation from zebrafish blood cells was performed using a commercial kit (High Pure PCR Template Preparation Kit, Roche Diagnostics, Basel, Switzerland). A final reaction volume of 25 µL containing PuREtaq Ready-to-go-PCR (GE Healthcare, Chicago, IL, USA), DNA (40 ng) and the primer 6 (5-d[CCCGTCAGCA]-30) (5 pmol/µL) was used for the amplification reaction: this cyclic program included an initial step (5 min—95 °C), followed by 45 cycles (1 min—95 °C, 1 min 36 °C and 2 min—72 °C). The amplification patterns were analyzed electrophoretically on 2% agarose gel containing 1X ethidium bromide. The RAPD-PCR profiles generated allowed the calculation of the genomic template stability (GTS %) for each experimental group as follows:
where a is the number of polymorphic bands detected in exposed groups (appearance of new bands and disappearance of bands respect to the control), and n is the total number of bands in the controls group.
GTS = (1 − a/n) × 100
2.7. Statistical Analysis
Differences among the experimental groups were analyzed using ANOVA test by GraphPad Prism 6. Data is reported as mean ± standard deviation (SD). Variation was considered significant when p-value (p) < 0.05.
3. Results
3.1. Viability Assay
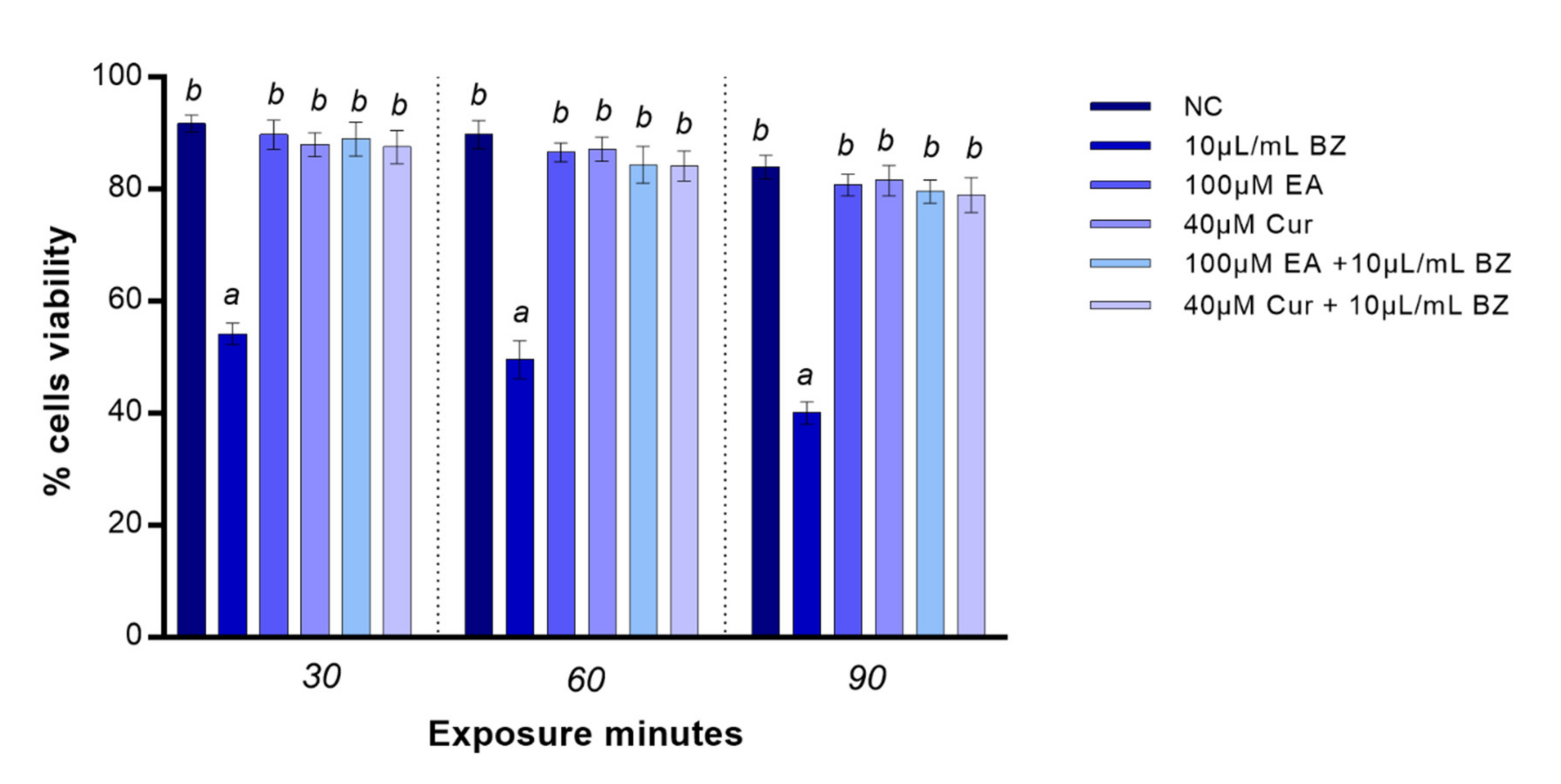
EA and Cur treatment did not affect zebrafish blood cells viability for all exposure times. Benzene provoked a significant (p < 0.05) reduction in viability already at 30 min. Benzene co-exposure with EA or Cur increased cells viability at all timepoints (p-value < 0.05) in comparison with benzene (Figure 1).
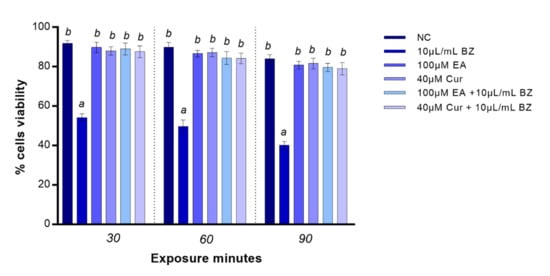
Figure 1.
Percentage of live (y-axis) zebrafish blood cells at different exposure timepoints (x-axis). Benzene—BZ; ellagic acid—EA; curcumin—Cur; negative control—NC. Letters correspond to diverse statistical significances (ANOVA); a: p < 0.05 in comparison with NC; b: p < 0.05 in comparison with BZ exposure.
3.2. Comet Assay
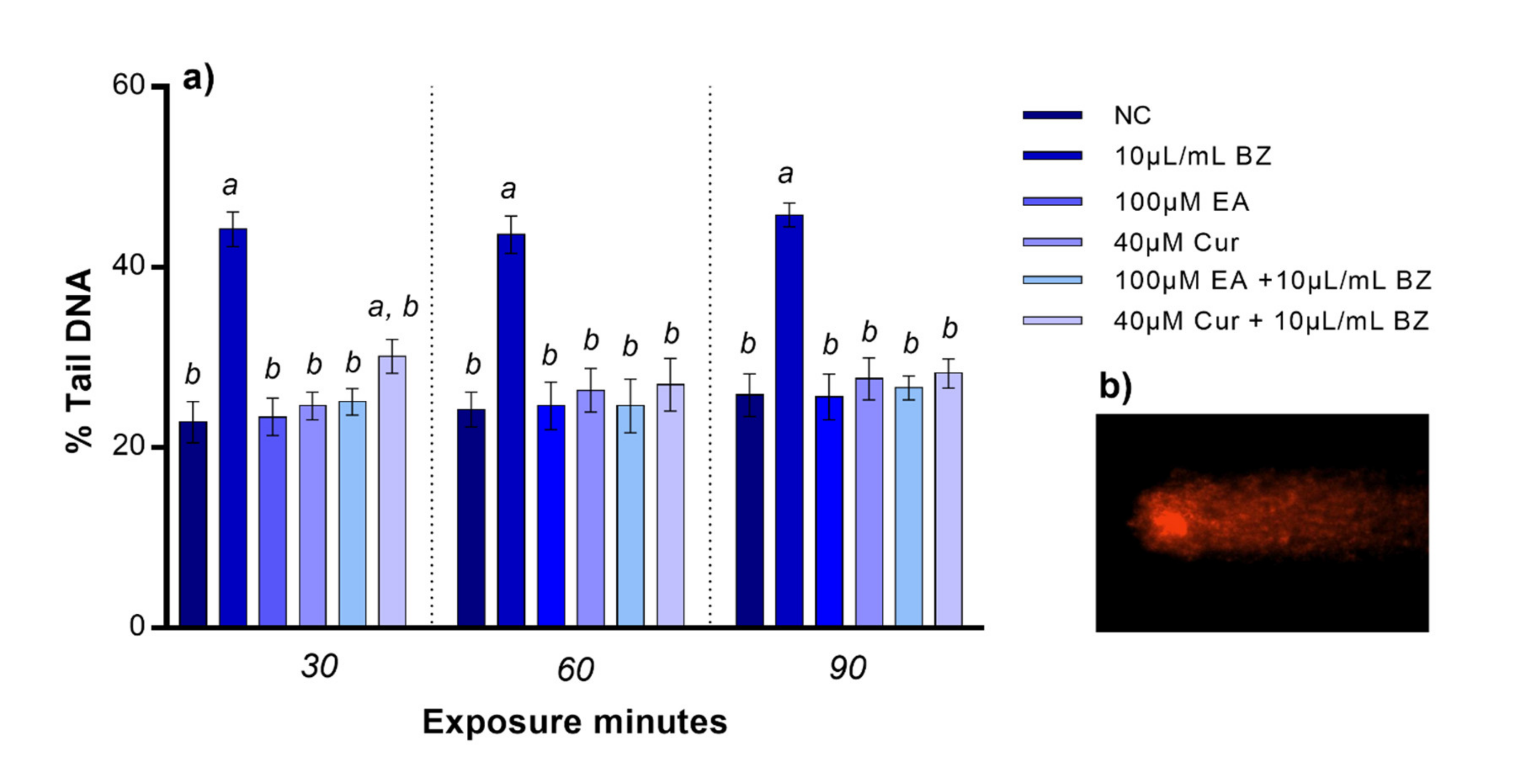
Following EA or Cur treatment, we did not observe any statistical variation in DNA damage at 30, 60, and 90 min. Benzene exposure resulted in reduced zebrafish blood cells DNA integrity (p < 0.05), starting at 30 min. When zebrafish blood cells were co-exposed to EA and benzene, DNA integrity was comparable to the negative control at all timepoints, while DNA damage was statistically significant after Cur and benzene co-exposure at 30 min, but not at 60 and 90 min, compared to the negative control (Figure 2).
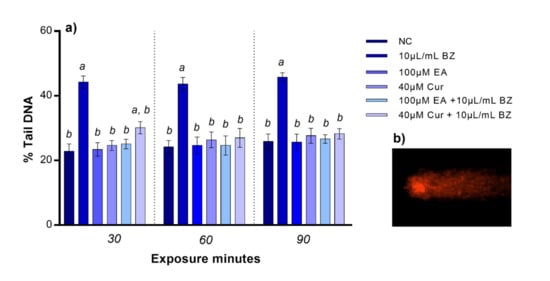
Figure 2.
(a) Percentage of DNA in the comet tail (y-axis) in zebrafish blood cells at different exposure timepoints (x-axis). Benzene—BZ; Ellagic acid—EA; curcumin—Cur; negative controls—NC. Letters correspond to diverse statistical significances (ANOVA); a: p < 0.05 in comparison with NC; b: p < 0.05 in comparison with BZ exposure. (b) Comet tail DNA in zebrafish blood cell analyzed using Komet 5.5 software.
3.3. Diffusion Assay
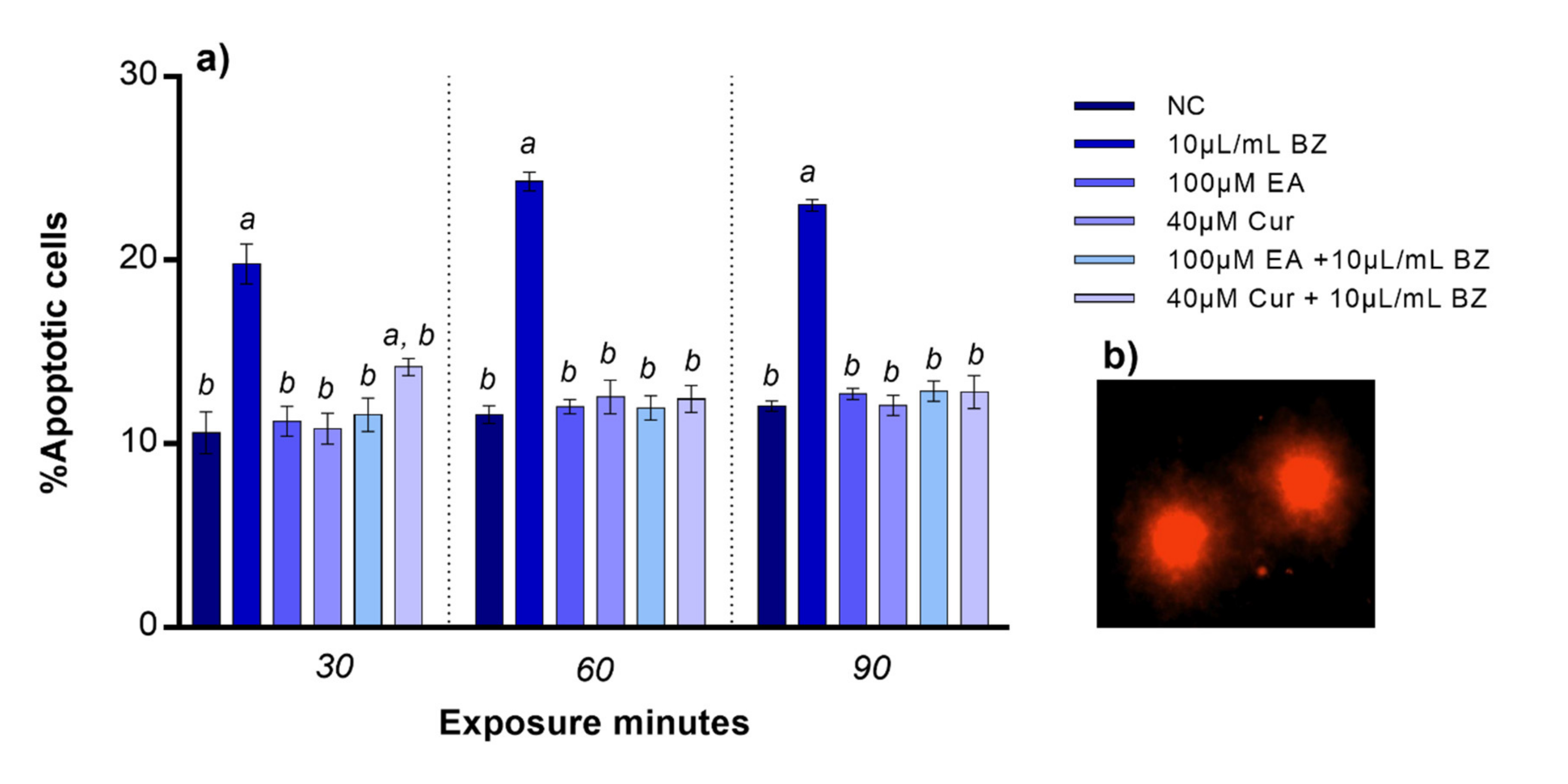
Apoptotic cells percentage was comparable to the negative control when zebrafish blood cells were exposed to EA or Cur. The percentage of apoptotic cells observed after benzene treatment significantly increased already at 30 min (p < 0.05). No variation in percentage of apoptotic blood cells was observed after co-exposure to EA and benzene at all exposure timepoints, while apoptosis was statistically significant after Cur and benzene co-exposure at 30 min, but not at 60 and 90 min, compared to the negative control (Figure 3).
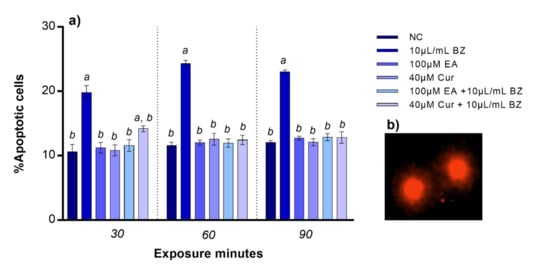
Figure 3.
(a) Percentage of apoptotic zebrafish blood cells (y-axis) at different exposure timepoints (x-axis). Benzene—BZ; Ellagic acid—EA; curcumin—Cur; negative controls—NC. Letters correspond to diverse statistical significances (ANOVA); a: p < 0.05 in comparison with NC; b: p < 0.05 in comparison with BZ exposure. (b) Apoptotic zebrafish blood cells analyzed with Nikon Eclipse E-600 microscope.
3.4. RAPD-PCR Technique
The electrophoretic DNA profile of zebrafish blood cells after benzene treatment resulted in appearance and disappearance of several bands at all three timepoints in comparison with the negative control. The exposure to EA did not result in any variation of bands while a band of 450 bp appeared after exposure to Cur at 90 min. Furthermore, co-exposure to EA and benzene determined the appearance of one new band of 750 bp at 90 min, while two bands and one band variation was detected at 30 and 60 min, respectively, after co-exposure to Cur and benzene (Table 1).

Table 1.
Molecular sizes (bp) of appeared and disappeared bands after amplification with primer 6 in zebrafish blood cells DNA exposed to different substances. EA: ellagic acid; Cur: curcumin.
3.5. Genomic Template Stability (% GTS)
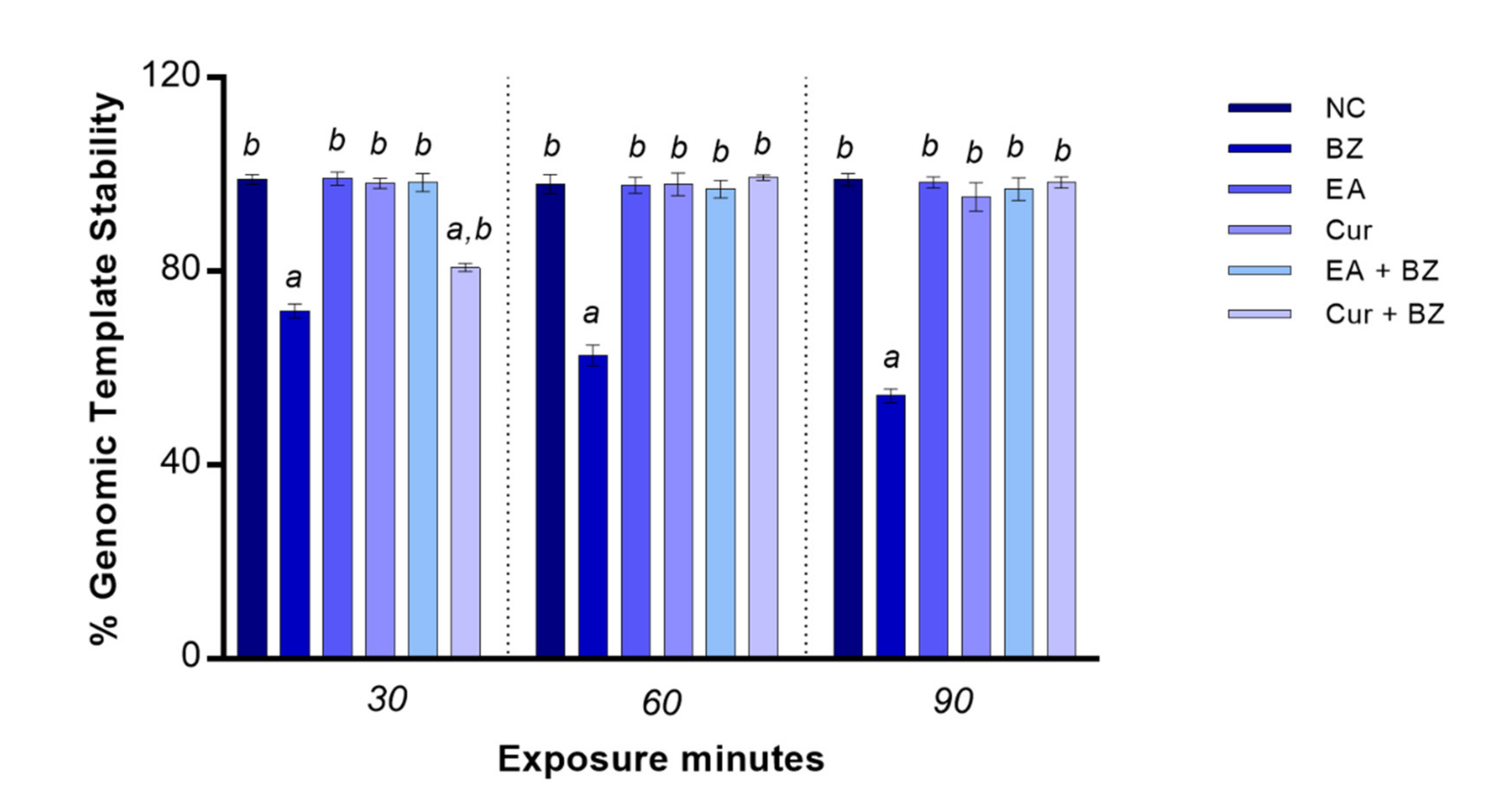
Exposure to benzene significantly reduced genome stability starting at 30 min (p < 0.05). When zebrafish blood cells were exposed to EA or Cur, the rate of genomic DNA stability was identical as per the negative control at all exposure timepoints. GTS% did not change when blood cells were co-exposed to EA and benzene in comparison with the negative control, while co-exposure to Cur and benzene determined a significant reduction of genome stability only at 30 min compared to the negative control (Figure 4).
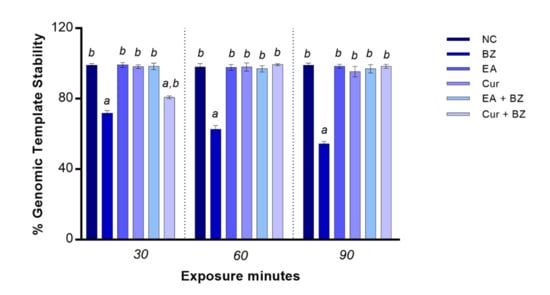
Figure 4.
Changes in percentage of genome template stability (y-axis) in zebrafish blood cells DNA at different exposure times (x-axis). Benzene—BZ; Ellagic acid—EA; curcumin—Cur; negative controls—NC. Letters correspond to diverse statistical significances (ANOVA); a: p < 0.05 in comparison with NC; b: p < 0.05 in comparison with BZ exposure.
4. Discussion
The antioxidant power of EA and Cur is well known, so the novelty of our research was to assess the compounds’ ability to counteract DNA damage caused by a known genotoxic agent (benzene), by using different methodologies in an experimental in vitro model of zebrafish blood cells. Our results confirm the genotoxicity of benzene and highlight EA and Cur as anti-genotoxic molecules, able to counteract ROS-mediated DNA damage.
We observed that DNA damage and apoptotic rate were increased when zebrafish blood cells were incubated with benzene. Benzene is a toxic chemical whose mutagenic effect has been studied for decades [32,33,34,35]. Benzene and its metabolites, such as 1,4-benzoquinone and hydroquinone, were described as cytotoxic agents able to induce DNA strand breaks in hamster ovary cells [34]. In human sperm cells, exposure to benzene resulted in reduced sperm motility and vitality, along with increased DNA denaturation [36]. In 2015, Huang et al. showed that hydroquinone (HQ), an important metabolite of benzene which is abundant in cigarette smoking, significantly reduced the expression of multidrug resistance mutation (MDR1) gene, encoding a membrane transporter, and inhibited the activation and nuclear translocation of the transcription factor NF-kB in mesenchymal stem cells derived from bone marrow [32]. This resulted in the induction of apoptotic pathways and reduced cellular vitality. Particularly, benzene cytotoxicity is mediated by the induction of OS [36]. Nishikawa et al. demonstrated that cells incubation in benzene resulted in increased ROS generation and apoptotic rate, along with upregulated expression of OS-related genes [35].
In the presence of noxious oxygen species, antioxidants play an important role in counterbalancing such effect and preserving the physiological redox balance, exerting an anti-genotoxic effect [10,37]. In this regard, several antioxidant compounds have been identified in plants or food as having the power to counteract the impact of genotoxic agents on DNA [38,39].
In this study, Cur was chosen for investigation as it acts as antioxidant but also upregulates the expression of transcription factors involved in the response to OS and other endogenous antioxidants, such as superoxide dismutase (SOD), catalase and glutathione (GSH) [40,41,42,43]. EA was selected because its antioxidant and anti-genotoxic potential has been pointed out by several studies, along with its anti-inflammatory and anti-pathogens properties [16,44,45,46,47]. EA and Cur tested concentrations were chosen based on preliminary tests and previous literature data [13,37,48,49].
Our data showed that EA and Cur were not harmful for zebrafish blood cells in vitro for the time and concentration investigated. Specifically, no variation was observed in comparison with the negative control at all timepoints analyzed in terms of cytotoxicity by trypan blue, DNA integrity by comet assay and RAPD-PCR and apoptotic rate by diffusion assay. However, the comet assay measures strand breaks, which are not always specific of OS, so in future experiments we plan to incorporate a formamidopyrimidine DNA glycosylase into the incubation assay, which converts oxidized purines into breaks, and is therefore more specific for oxidative damage [50].
We observed that incubation of zebrafish blood cells in EA nullifies the impact of benzene on all the above-mentioned outcomes at all timepoints analyzed, exerting a relevant antioxidant and anti-genotoxic effect. EA is a phenolic molecule, which is abundant in specific plants, mushrooms and food sources [51,52].
The molecular functions elicited by EA include scavenging of free radicals, regulation of lipid metabolism, and inhibition of fibrogenesis response-mediating proteins. Moreover, it restrains hepatic viral replication, facilitates suppression of growth factors, regulates transcription factors, the expression of proinflammatory cytokines, and augments the liver immune response [47]. Lately, EA has sparked considerable interest as a potential chemotherapeutic compound as it can induce apoptosis and inhibit the proliferation of various types of cancer cells in vitro and in vivo through the regulation of apoptosis and energy metabolism. However, EA does not have a single target of action, but has effects that are highly dependent on cell type and cell metabolism. This is because EA can be transported into the cell through multiple mechanisms that influence its intracellular accumulation, especially in tumor cells where the transporters are differentially expressed (organic anion transporters (OATP), SLC22A (OAT1 and OAT4) families, sodium-dependent glucose transporter (SGLT1)). Thus, the usefulness of this compound as a cancer therapeutic is limited [53].
EA reportedly acts as a scavenger for free radicals and prevents the OS-related induction of apoptosis [54]. Its antioxidant properties are exerted through the regulation of OS-related pathways, such as Nrf2, cyclooxygenase 2, and NF-kB [55,56,57,58], which result in higher expression and activity of endogenous antioxidants [59], and in the inhibition of NADPH oxidase activity and ROS generation [60,61]. Moreover, it is reported to inhibit the proliferation of cervical cancer by regulating the expression of IGBPF7 in a dose-dependent manner [62]. Our current study proved that the ability of Cur to counteract DNA damage provoked by benzene is expressed for intermediate and prolonged times of exposure (60 and 90 min) on zebrafish blood cells in vitro. This could be explained by the fact that Cur exhibits poor bioavailability due to poor absorption [63]. The anti-inflammatory properties of Cur have been described as comparable as of steroidal and nonsteroidal drugs, by inhibiting inflammatory enzymes (cyclooxygenases, lipoxygenases, nitric oxide synthase) and production of cytokines [64,65,66,67,68]. Furthermore, Cur has been reported to reduce the activation of the NF-kB pathway, hyperactivated in conditions of OS, by inhibiting IKB kinase and downregulating the expression of other genes involved in this pathway [69]. Also, it regulates the expression of tumor necrosis factor (TNF), leading to reduced cell proliferation [70].
The mechanism of action of Cur depends both on its chemical structure characterized by the presence of functional antioxidant groups, such as β-diket groups, carbon-carbon double bonds and phenylic rings, and on its ability to stimulate the production of antioxidant enzymes. In fact, it is able to regulate the expression of transcription factors and antioxidant enzymes, such as heme oxygenase-1 (HO-1), factor 2 related to nuclear erythroid factor 2 (Nrf2), SOD and GSH), and inhibits cytokine production (interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), interleukin 12 (IL-12)) [71]. Its antitumor action occurs through the regulation of caspases-3 and -4 activities, intracellular calcium ion, ATM levels and cell cycle arrest in the G2/M phase, while the ability to protect the nervous system comes from its effect on Akt, Wnt/β catenin and Aβ peptide phosphorylation. Moreover, Cur prevents gastrointestinal diseases and supports cardio-protection by acting on Treg and PKCδ cells and through the effect on DKK-3 [72]. Recently, a randomized, double-blind, placebo-controlled clinical trial was conducted to investigate the effects of Cur in patients with non-alcoholic fatty liver disease [73]. Investigators observed no changes in 8-hydroxy-2′-deoxyguanosine (8-OHdG) levels, a DNA marker of OS [73]. However, they reported reduced methylation in MutL homolog 1 (MLH1) and MutS homolog 2 (MSH2) genes, which are involved in DNA mismatch repair mechanism. This study suggested for the first time that Cur may exert an antigenotoxic action through the regulation of epigenetic mechanisms.
Our results support the use of Cur and EA as antigenotoxic agents to be considered for the prevention of OS-associated genotoxicity. A therapeutic use of Cur has been investigated in several pathological conditions where OS plays a contributive role. In cancer, it exerts an antiproliferative effect, acting as a chemo-preventive agent [74], while, in atherosclerosis, it prevents platelet aggregation by inhibiting thromboxane [75]. Its antioxidant properties have been reported to be beneficial also in neurodegenerative pathologies [76], liver fibrosis [77], diabetes [78], and autoimmune diseases [79]. Importantly, animal studies revealed that Cur supplementation can reduce chronic alcohol and nicotine-induced toxicity [80,81]. Similarly, an in vitro study revealed that EA protects against nicotine-induced cellular DNA damage and lipid peroxidation [82]. Animal studies investigated the use of EA to counterbalance the negative effects of chemotherapeutic agents for cancer therapy. Both Ceribase et al. and Rehman et al. reported normalized semen parameters, antioxidants concentration and levels of lipid peroxidation, when adriamycin and cyclophosphamide treatments were associated with EA, respectively [83,84]. In a placebo-controlled double blind trial, cyclists drinking beverages supplemented with EA showed lower levels of carbonyls and 8-OHdG, both markers of OS, after intensive exercise [85].
Despite encouraging results, our study is not free of limitations. Although an in vitro approach is useful to understand the mechanism of actions, it is difficult to forecast effects due to molecular interactions between cells in different tissues or long-term exposure, as in an in vivo system. Moreover, it is difficult to compare the concentration tested with those likely to be introduced through normal nutrition (curcuma longa 1–2 μg/g), as well as to establish the optimal concentrations to be used for therapeutic purposes starting from in vitro experiments. In fact, in an in vivo system, Cur is subjected to low absorption, poor bioavailability and rapid metabolism. Therefore, the determination of the pharmacokinetic profile of Cur appears difficult due to low circulating concentrations [86]. However, the safety of the substance is maintained in vivo even after the administration of very high doses [87]; in addition, other in vitro studies have demonstrated its efficacy at concentrations ranging from 5 to 50 μM [49,88]. Similarly, Ea is poorly bioavailable and non-absorbable, despite being a very small molecule. In fact, it passes unaltered to the intestinal tract, where it undergoes extensive metabolism by the intestinal microbiota providing urolithins (UROs), that are more bioavailable and are much better absorbed than EA. Even when consumed in very large quantities (800 mg of extract), the maximum concentration of EA in plasma was reported to be very low and not sufficient to produce significant beneficial effects [89]. The activity of the human microbiota is difficult to reproduce in animal models and cannot be easily studied and controlled in vitro. Hence, we had the same difficulties with EA to compare the concentration tested with those likely to be present through normal nutrition and used therapeutically.
In this study we used only one concentration of EA and Cur, but a variety of assays with the primary goal of studying different antigenotoxicity endpoints. Therefore, our future goal will be to evaluate the effects of different EA and Cur concentrations for dose-response experiment, including at increasing exposure times. We cannot exclude that higher concentrations may have a negative influence on zebrafish physiology. Also, it has been shown that antioxidants exert a higher effect when used in combination [90,91,92,93]. Pavan et al. demonstrated a synergic antioxidant effect of resveratrol and Cur for treating cancer [94]. The synergistic effect of EA and Cur was investigated in vitro on cervical carcinoma cells [95]. In this study, authors showed EA and Cur to have anticancer properties by inhibiting ROS generation and inducing apoptosis through the p53 pathway. Moreover, the antigenotoxic effect of antioxidants was enhanced when they were used in combination [37], laying the foundation for further studies investigating their combined use as anti-genotoxic compounds. Hence, the next step in our research will include the investigation of the synergistic anti-genotoxic effect of Cur and EA.
5. Conclusions
This study highlighted the activity of EA and Cur as anti-genotoxic molecules due to their positive effects on cytotoxicity, DNA integrity and apoptotic rate in an in vitro model of zebrafish blood cells treated with benzene. Besides their anti-genotoxic, anti-pathogenic, and antioxidant potential, EA and Cur arouse great interest in the scientific community because of their natural origin. Hence, they may be integrated into a diet for the prevention of several diseases and disorders.
Author Contributions
Conceptualization, F.M.; methodology, M.S., C.I. and M.C.; validation, F.M. and L.R.; data curation, R.F.; writing—original draft preparation, R.F.; writing—review and editing, R.F. and F.M.; supervision, L.R.; project administration, L.R.; funding acquisition, L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially financed by the University Research Funds (University of Campania “Luigi Vanvitelli”, grant n. 2020).
Institutional Review Board Statement
The study was conducted according to the recommendations reported in the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes No animals were sacrificed.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included; any additional information is available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Turkez, H.; Arslan, M.E.; Ozdemir, O. Genotoxicity testing: Progress and prospects for the next decade. Expert Opin. Drug Metab. Toxicol. 2017, 13, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxidative Med. Cell. Longev. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nourazarian, A.; Kangari, P.; Salmaninejad, A. Roles of Oxidative Stress in the Development and Progression of Breast Cancer. Asian Pac. J. Cancer Prev. 2014, 15, 4745–4751. [Google Scholar] [CrossRef]
- Turner, A. Cadmium pigments in consumer products and their health risks. Sci. Total Environ. 2019, 657, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Mottola, F.; Santonastaso, M.; Iovine, C.; Feola, V.; Pacifico, S.; Rocco, L. Adsorption of Cd to TiO2-NPs Forms Low Genotoxic AGGREGATES in Zebrafish Cells. Cells 2021, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Handl, J.; Čapek, J.; Majtnerová, P.; Petira, F.; Hauschke, M.; Roušarová, E.; Roušar, T. Transient increase in cellular dehydrogenase activity after cadmium treatment precedes enhanced production of reactive oxygen species in human proximal tubular kidney cells. Physiol. Res. 2019, 68, 481–490. [Google Scholar] [CrossRef]
- Santonastaso, M.; Mottola, F.; Iovine, C.; Cesaroni, F.; Colacurci, N.; Rocco, L. In Vitro Effects of Titanium Dioxide Nanoparticles (TiO2NPs) on Cadmium Chloride (CdCl2) Genotoxicity in Human Sperm Cells. Nanomaterials 2020, 10, 1118. [Google Scholar] [CrossRef]
- Goswami, P.; Banerjee, R.; Mukherjee, A. Potential antigenotoxicity assessment of Ziziphus jujuba fruit. Heliyon 2019, 5, e01768. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.M.; Figueiredo, C.C.; Gomes, A.C.; Ferreira, P.C.; Granero, F.O.; Santiago, P.S.; Silva, L.P. Evaluation of the antigenotoxic and antioxidant activity induced by Croton antisyphiliticus. An. Acad. Bras. Cienc. 2020, 92, e20190047. [Google Scholar] [CrossRef]
- Rocco, L.; Mottola, F.; Santonastaso, M.; Saputo, V.; Cusano, E.; Costagliola, D.; Suero, T.; Pacifico, S.; Stingo, V. Anti-genotoxic ability of α-tocopherol and Anthocyanin to counteract fish DNA damage induced by musk xylene. Ecotoxicology 2015, 24, 2026–2035. [Google Scholar] [CrossRef]
- McLaughlin, P.J.; Weihrauch, J.L. Vitamin E content of foods. J. Am. Diet. Assoc. 1979, 75, 647–665. [Google Scholar]
- Mazza, G.; Miniati, E. Anthocyanins in Fruits, Vegetables and Grains, 1st ed.; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Iovine, C.; Mottola, F.; Santonastaso, M.; Finelli, R.; Agarwal, A.; Rocco, L. In vitro ameliorative effects of ellagic acid on vitality, motility and DNA quality in human spermatozoa. Mol. Reprod. Dev. 2021, 88, 167–174. [Google Scholar] [CrossRef]
- Cao, J.; Jiang, L.-P.; Liu, Y.; Yang, G.; Yao, X.-F.; Zhong, L.-F. Curcumin-induced genotoxicity and antigenotoxicity in HepG2 cells. Toxicon 2007, 49, 1219–1222. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.H.; Zhao, W.W.; Chen, X.P. Research progress on the anti-cancer effect and mechanisms of the natural tanshinones. Chin. Pharmacol. Bull. 2014, 30, 1041–1044. [Google Scholar] [CrossRef]
- Baek, B.; Lee, S.H.; Kim, K.; Lim, H.-W.; Lim, C.-J. Ellagic acid plays a protective role against UV-B-induced oxidative stress by up-regulating antioxidant components in human dermal fibroblasts. Korean, J. Physiol. Pharmacol. 2016, 20, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Galano, A.; Marquez, M.F.; Pérez-González, A. Ellagic Acid: An Unusually Versatile Protector against Oxidative Stress. Chem. Res. Toxicol. 2014, 27, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Tang, S.-N.; Marsh, J.L.; Shankar, S.; Srivastava, R.K. Ellagic acid inhibits human pancreatic cancer growth in Balb c nude mice. Cancer Lett. 2013, 337, 210–217. [Google Scholar] [CrossRef]
- Mottola, F.; Scudiero, N.; Iovine, C.; Santonastaso, M.; Rocco, L. Protective activity of ellagic acid in counteract oxidative stress damage in zebrafish embryonic development. Ecotoxicol. Environ. Saf. 2020, 197, 110642. [Google Scholar] [CrossRef] [PubMed]
- El-Hack, M.E.A.; El-Saadony, M.T.; Swelum, A.A.; Arif, M.; Ghanima, M.M.A.; Shukry, M.; Noreldin, A.; Taha, A.E.; El-Tarabily, K.A. Curcumin, the active substance of turmeric: Its effects on health and ways to improve its bioavailability. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Alalwan, H.K.A.; Rajendran, R.; Lappin, D.F.; Combet, E.; Shahzad, M.; Robertson, D.; Nile, C.; Williams, C.; Ramage, G. The Anti-Adhesive Effect of Curcumin on Candida albicans Biofilms on Denture Materials. Front. Microbiol. 2017, 8, 659. [Google Scholar] [CrossRef]
- Panda, A.K.; Chakraborty, D.; Sarkar, I.; Khan, T.; Sa, G. New insights into therapeutic activity and anticancer properties of curcumin. J. Exp. Pharmacol. 2017, 9, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, M.; Kang, M.-C.; Lee, H.; Cho, C.; Choi, I.; Park, Y.; Lee, S.-H. Antioxidant Effects of Turmeric Leaf Extract against Hydrogen Peroxide-Induced Oxidative Stress In Vitro in Vero Cells and In Vivo in Zebrafish. Antioxidants 2021, 10, 112. [Google Scholar] [CrossRef]
- Whysner, J.; Reddy, M.V.; Ross, P.M.; Mohan, M.; Lax, E.A. Genotoxicity of benzene and its metabolites. Mutat. Res. Mutat. Res. 2003, 566, 99–130. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J. Making Waves: New Developments in Toxicology with the Zebrafish. Toxicol. Sci. 2018, 163, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Directive 2010/63/EU Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; 2010; Volume 276, pp. 1–47. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 1 September 2021).
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21, A–3B. [Google Scholar]
- Mottola, F.; Iovine, C.; Santonastaso, M.; Romeo, M.L.; Pacifico, S.; Cobellis, L.; Rocco, L. NPs-TiO2 and lincomycin coexposure induces DNA damage in cultured human amniotic cells. Nanomaterials 2019, 9, 1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyori, B.M.; Venkatachalam, G.; Thiagarajan, P.; Hsu, D.; Clement, M.-V. OpenComet: An automated tool for comet assay image analysis. Redox Biol. 2014, 2, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.P. A Simple Method for Accurate Estimation of Apoptotic Cells. Exp. Cell Res. 2000, 256, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Cantafora, E.; Giorgi, F.S.; Frenzilli, G.; Scarcelli, V.; Busceti, C.L.; Nigro, M.; Bernardeschi, M.; Fornai, F. Region-specific DNA alterations in focally induced seizures. J. Neural Transm. 2014, 121, 1399–1403. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, M.; Li, X.; Ma, L.; Zhang, J.; Shi, J.; Li, B.; Fan, W.; Zhou, Y. The Cytotoxic Effect of the Benzene Metabolite Hydroquinone is Mediated by the Modulation of MDR1 Expression via the NF-κB Signaling Pathway. Cell. Physiol. Biochem. 2015, 37, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Spanò, M.; Pacchierotti, F.; Uccelli, R.; Amendola, R.; Bartoleschi, C. Cytotoxic effects of benzene on mouse germ cells determined by flow cytometry. J. Toxicol. Environ. Health Part A 1989, 26, 361–372. [Google Scholar] [CrossRef]
- Sze, C.; Shi, C.; Ong, C. Cytotoxicity and DNA strand breaks induced by benzene and its metabolites in Chinese hamster ovary cells. J. Appl. Toxicol. 1996, 16, 259–264. [Google Scholar] [CrossRef]
- Nishikawa, T.; Izumo, K.; Miyahara, E.; Horiuchi, M.; Okamoto, Y.; Kawano, Y.; Takeuchi, T. Benzene Induces Cytotoxicity without Metabolic Activation. J. Occup. Heal. 2011, 53, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandani, P.; Desai, K.; Highland, H. Cytotoxic Effects of Benzene Metabolites on Human Sperm Function: An In Vitro Study. ISRN Toxicol. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mottola, F.; Santonastaso, M.; Iovine, C.; Rossetti, C.; Ronga, V.; Rocco, L. DNA Damage in Human Amniotic Cells: Antigenotoxic Potential of Curcumin and α-Lipoic Acid. Antioxidants 2021, 10, 1137. [Google Scholar] [CrossRef]
- Romero-Jiménez, M.; Campos-Sánchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, A. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 585, 147–155. [Google Scholar] [CrossRef]
- Iddir, M.; Brito, A.; Dingeo, G.; Del Campo, S.S.F.; Samouda, H.; La Frano, M.R.; Bohn, T. Strengthening the Immune System and Reducing Inflammation and Oxidative Stress through Diet and Nutrition: Considerations during the COVID-19 Crisis. Nutrients 2020, 12, 1562. [Google Scholar] [CrossRef]
- Santonastaso, M.; Mottola, F.; Iovine, C.; Colacurci, N.; Rocco, L. Protective Effects of Curcumin on the Outcome of Cryopreservation in Human Sperm. Reprod. Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Ak, T.; Gülçin, I. Antioxidant and radical scavenging properties of curcumin. Chem. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef]
- Balogun, E.; Hoque, M.; Gong, P.; Killeen, E.; Green, C.J.; Foresti, R.; Alam, J.; Motterlini, R. Curcumin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive element. Biochem. J. 2003, 371, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meshkibaf, M.H.; Maleknia, M.; Noroozi, S. Effect of curcumin on gene expression and protein level of methionine sulfoxide reductase A (MSRA) SOD, CAT and GPx in Freund’s adjuvant inflammation-induced male rats. J. Inflamm. Res. 2019, 12, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Zeb, A. Ellagic acid in suppressing in vivo and in vitro oxidative stresses. Mol. Cell. Biochem. 2018, 448, 27–41. [Google Scholar] [CrossRef]
- Ceci, C.; Lacal, P.M.; Tentori, L.; De Martino, M.G.; Miano, R.; Graziani, G. Experimental Evidence of the Antitumor, Antimetastatic and Antiangiogenic Activity of Ellagic Acid. Nutrients 2018, 10, 1756. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, V.B.; Ghadiri, M.; Ramezani, M.; Askari, V.R. Antiinflammatory and anti-cancer activities of pomegranate and its constituent, ellagic acid: Evidence from cellular, animal, and clinical studies. Phytotherapy Res. 2020, 34, 685–720. [Google Scholar] [CrossRef] [PubMed]
- Aishwarya, V.; Solaipriya, S.; Sivaramakrishnan, V. Role of ellagic acid for the prevention and treatment of liver diseases. Phytother. Res. 2020, 35, 2925–2944. [Google Scholar] [CrossRef] [PubMed]
- Kharat, P.; Sarkar, P.; Mouliganesh, S.; Tiwary, V.; Priya, V.R.; Sree, N.Y.; Annapoorna, H.V.; Saikia, D.K.; Mahanta, K.; Thirumurugan, K. Ellagic acid prolongs the lifespan of Drosophila melanogaster. GeroScience 2019, 42, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.L.; Luis, P.B.; Varuzza, P.V.; Joseph, A.I.; Presley, S.H.; Chaturvedi, R.; Schneider, C. The anti-inflammatory activity of curcumin is mediated by its oxidative metabolites. J. Biol. Chem. 2017, 292, 21243–21252. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. Measuring oxidative damage to DNA and its repair with the comet assay. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Daniel, E.M.; Krupnick, A.S.; Heur, Y.-H.; Blinzler, J.A.; Nims, R.W.; Stoner, G.D. Extraction, stability, and quantitation of ellagic acid in various fruits and nuts. J. Food Compos. Anal. 1989, 2, 338–349. [Google Scholar] [CrossRef]
- Zeb, A. A reversed phase HPLC-DAD method for the determination of phenolic compounds in plant leaves. Anal. Methods 2015, 7, 7753–7757. [Google Scholar] [CrossRef]
- Boehning, A.L.; Essien, S.A.; Underwood, E.L.; Dash, P.K.; Boehning, D. Cell type-dependent effects of ellagic acid on cellular metabolism. Biomed. Pharmacother. 2018, 106, 411–418. [Google Scholar] [CrossRef]
- Dalvi, L.T.; Moreira, D.C.; Andrade, R.; Ginani, J.; Alonso, A.; Hermes-Lima, M. Ellagic acid inhibits iron-mediated free radical formation. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 173, 910–917. [Google Scholar] [CrossRef]
- Wang, H.R.; Sui, H.C.; Zhu, B.T. Ellagic acid, a plant phenolic compound, activates cyclooxygenase-mediated prostaglandin production. Exp. Ther. Med. 2019, 18, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.C.; Teuber, S.S. Ellagic acid and polyphenolics present in walnut kernels inhibit in vitro human peripheral blood mononuclear cell proliferation and alter cytokine production. Ann. N. Y. Acad. Sci. 2010, 1190, 86–96. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, B.; Zhou, K.; Chen, M.; Wang, M.; Jia, Y.; Song, Y.; Li, Y.; Wen, A. Dietary ellagic acid improves oxidant-induced endothelial dysfunction and atherosclerosis: Role of Nrf2 activation. Int. J. Cardiol. 2014, 175, 508–514. [Google Scholar] [CrossRef]
- Chatterjee, A.; Chatterjee, S.; Das, S.; Saha, A.; Chattopadhyay, S.; Bandyopadhyay, S.K. Ellagic acid facilitates indomethacin-induced gastric ulcer healing via COX-2 up-regulation. Acta Biochim. Biophys. Sin. 2012, 44, 565–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.H.; Lee, M.J.; Kim, J.H. Antioxidant and apoptosis-inducing activities of ellagic acid. Anticancer. Res. 2006, 26, 3601–3606. [Google Scholar] [PubMed]
- Lee, W.-J.; Ou, H.-C.; Hsu, W.-C.; Chou, M.-M.; Tseng, J.-J.; Hsu, S.-L.; Tsai, K.-L.; Sheu, W.H.-H. Ellagic acid inhibits oxidized LDL-mediated LOX-1 expression, ROS generation, and inflammation in human endothelial cells. J. Vasc. Surg. 2010, 52, 1290–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.M.; Cho, J.S.; Kim, T.H.; Lee, Y.I. Ellagic acid protects hepatocytes from damage by inhibiting mitochondrial production of reactive oxygen species. Biomed. Pharmacother. 2010, 64, 264–270. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, D.; Fu, Q. Inhibition of Cervical Cancer by Promoting IGFBP7 Expression Using Ellagic Acid from Pomegranate Peel. Med. Sci. Monit. 2016, 22, 4881–4886. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Huang, M.T.; Newmark, H.L.; Frenkel, K. Inhibitory effects of curcumin on tumorigenesis in mice. J. Cell. Biochem. Suppl. 1997, 67, 26–34. [Google Scholar] [CrossRef]
- Brouet, I.; Ohshima, H. Curcumin, an Anti-tumor Promoter and Anti-inflammatory Agent, Inhibits Induction of Nitric Oxide Synthase in Activated Macrophages. Biochem. Biophys. Res. Commun. 1995, 206, 533–540. [Google Scholar] [CrossRef]
- Bernd, A.; Theilig, C.; Kippenberger, S.; Ramirez-Bosca, A.; Diaz, J.; Miquel, J.; Kaufmann, R. Effect of curcuma longa extract on the expression of proinflammatory cytokines. Skin Pharm. Apll Skin Physiol. 2000, 13, 226–234. [Google Scholar]
- Chan, M.M.-Y.; Huang, H.-I.; Fenton, M.R.; Fong, D. In Vivo Inhibition of Nitric Oxide Synthase Gene Expression by Curcumin, a Cancer Preventive Natural Product with Anti-Inflammatory Properties. Biochem. Pharmacol. 1998, 55, 1955–1962. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and Anti-Inflammatory Properties of Curcumin. Adv. Exp. Med. Biol. 2007, 595, 105–125. [Google Scholar] [CrossRef]
- Sarkar, F.H.; Li, Y. Cell signaling pathways altered by natural chemopreventive agents. Mutat. Res. Mol. Mech. Mutagen. 2004, 555, 53–64. [Google Scholar] [CrossRef]
- Shishodia, S.; Amin, H.M.; Lai, R.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive NF-κB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem. Pharmacol. 2005, 70, 700–713. [Google Scholar] [CrossRef]
- Wright, J.S. Predicting the antioxidant activity of curcumin and curcuminoids. J. Mol. Struct. THEOCHEM 2002, 591, 207–217. [Google Scholar] [CrossRef]
- Liczbiński, P.; Michałowicz, J.; Bukowska, B. Molecular mechanism of curcumin action in signaling pathways: Review of the latest research. Phytotherapy Res. 2020, 34, 1992–2005. [Google Scholar] [CrossRef] [PubMed]
- Hariri, M.; Gholami, A.; Mirhafez, S.R.; Bidkhori, M.; Sahebkar, A. A pilot study of the effect of curcumin on epigenetic changes and DNA damage among patients with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled, clinical trial. Complement. Ther. Med. 2020, 51, 102447. [Google Scholar] [CrossRef] [PubMed]
- Nagabhushan, M.; Bhide, S. Curcumin as an Inhibitor of Cancer. J. Am. Coll. Nutr. 1992, 11, 192–198. [Google Scholar] [CrossRef]
- Srivastava, R.; Dikshit, M.; Srimal, R.; Dhawan, B. Anti-thrombotic effect of curcumin. Thromb. Res. 1985, 40, 413–417. [Google Scholar] [CrossRef]
- Grundman, M.; Delaney, P. Antioxidant strategies for Alzheimer’s disease. Proc. Nutr. Soc. 2002, 61, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akila, G.; Rajakrishnan, V.; Viswanathan, P.; Rajashekaran, K.N.; Menon, V.P. Effects of curcumin on lipid profile and lipid peroxidation status in experimental hepatic fibrosis. Hepatol. Res. 1998, 11, 147–157. [Google Scholar] [CrossRef]
- Sajithlal, G.; Chithra, P.; Chandrakasan, G. Effect of curcumin on the advanced glycation and cross-linking of collagen in diabetic rats. Biochem. Pharmacol. 1998, 56, 1607–1614. [Google Scholar] [CrossRef]
- Bosman, B. Testing of Lipoxygenase Inhibitors, Cyclooxygenase Inhibitors, Drugs with Immunomodulating Properties and Some Reference Antipsoriatic Drugs in the Modified Mouse Tail Test, an Animal Model of Psoriasis. Skin Pharmacol. Physiol. 1994, 7, 324–334. [Google Scholar] [CrossRef]
- Baliga, M.S.; Rao, S.; Rao, P.; Pais, M.L.J.; Naik, T.S.; Adnan, M.; Palatty, P.L. Hepatoprotective effects of curcumin in alcohol-induced hepatotoxicity: A memoir on the preclinical studies. In Polyphenols: Prevention and Treatment of Human Disease; Watson, R., Preedy, V., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 313–317. ISBN 9780128130087. [Google Scholar]
- Polasa, K.; Naidu, A.; Ravindranath, I.; Krishnaswamy, K. Inhibition of B(a)P induced strand breaks in presence of curcumin. Mutat. Res. Toxicol. Environ. Mutagen. 2003, 557, 203–213. [Google Scholar] [CrossRef]
- Sudheer, A.R.; Muthukumaran, S.; Devipriya, N.; Menon, V.P. Ellagic acid, a natural polyphenol protects rat peripheral blood lymphocytes against nicotine-induced cellular and DNA damage in vitro: With the comparison of N-acetylcysteine. Toxicology 2007, 230, 11–21. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Ali, F.; Qamar, W.; Lateef, A.; Khan, R.; Quaiyoom, A.; Hamiza, O.O.; Sultana, S. Cyclophosphamide-induced nephrotoxicity, genotoxicity, and damage in kidney genomic DNA of Swiss albino mice: The protective effect of Ellagic acid. Mol. Cell. Biochem. 2012, 365, 119–127. [Google Scholar] [CrossRef]
- Çeribaşı, A.O.; Sakin, F.; Türk, G.; Sönmez, M.; Ateşşahin, A. Impact of ellagic acid on adriamycin-induced testicular histopathological lesions, apoptosis, lipid peroxidation and sperm damages. Exp. Toxicol. Pathol. 2012, 64, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morillas-Ruiz, J.; Zafrilla, P.; Almar, M.; Cuevas, M.J.; López, F.J.; Abellán, P.; Villegas, J.A.; González-Gallego, J. The effects of an antioxidant-supplemented beverage on exercise-induced oxidative stress: Results from a placebo-controlled double-blind study in cyclists. Graefe’s Arch. Clin. Exp. Ophthalmol. 2005, 95, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Cas, M.D.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [Green Version]
- Vollono, L.; Falconi, M.; Gaziano, R.; Iacovelli, F.; Dika, E.; Terracciano, C.; Bianchi, L.; Campione, E. Potential of curcumin in skin disorders. Nutrients 2019, 11, 2169. [Google Scholar]
- Wang, X.; Gao, J.; Wang, Y.; Zhao, B.; Zhang, Y.; Han, F.; Zheng, Z.; Hu, D. Curcumin pretreatment prevents hydrogen peroxide-induced oxidative stress through enhanced mitochondrial function and deactivation of Akt/Erk signaling pathways in rat bone marrow mesenchymal stem cells. Mol. Cell. Biochem. 2017, 443, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative stress, antioxidant capabilities, and bioavailability: Ellagic acid or urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liu, S.; Chen, M.; Li, J.; Huang, D.; Zhu, S. Synergistic effects of ascorbic acid and plant-derived ceramide to enhance storability and boost antioxidant systems of postharvest strawberries. J. Sci. Food Agric. 2019, 99, 6562–6571. [Google Scholar] [CrossRef] [PubMed]
- Crespo, Y.A.; Sánchez, L.R.B.; Quintana, Y.G.; Cabrera, A.S.T.; del Sol, A.B.; Mayancha, D.M.G. Evaluation of the synergistic effects of antioxidant activity on mixtures of the essential oil from Apium graveolens L., Thymus vulgaris L. and Coriandrum sativum L. using simplex-lattice design. Heliyon 2019, 5, e01942. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.; Wang, K.; Lei, Y.; Yao, S.; Lu, B.; Huang, W. Antioxidant synergistic effects of Osmanthus fragrans flowers with green tea and their major contributed antioxidant compounds. Sci. Rep. 2017, 7, 46501. [Google Scholar] [CrossRef] [Green Version]
- Abdulkhaleq, F.M.; Alhussainy, T.M.; Badr, M.M.; Abu Khalil, A.A.; Gammoh, O.; Ghanim, B.Y.; Qinna, N.A. Antioxidative stress effects of vitamins C, E, and B12, and their combination can protect the liver against acetaminophen-induced hepatotoxicity in rats. Drug Des. Dev. Ther. 2018, 12, 3525–3533. [Google Scholar] [CrossRef] [Green Version]
- Pavan, A.R.; Da Silva, G.D.B.; Jornada, D.H.; Chiba, D.E.; Fernandes, G.F.D.S.; Chin, C.M.; Dos Santos, J.L. Unraveling the Anticancer Effect of Curcumin and Resveratrol. Nutrients 2016, 8, 628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Basu, S.; Parija, L.; Rout, D.; Manna, S.; Dandapat, J.; Debata, P.R. Curcumin and Ellagic acid synergistically induce ROS generation, DNA damage, p53 accumulation and apoptosis in HeLa cervical carcinoma cells. Biomed. Pharmacother. 2016, 81, 31–37. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).