Abstract
Cisplatin is one of the most effective anticancer agents used to treat colon cancer, which is the third malignancy between the most common human cancers in the world, but the resistance developed represents an obstacle against the full success of chemotherapy. An emerging interest appeared in finding other metallic compounds, such as ruthenium(III) complexes, for chemotherapeutic application in cancer. Our study focused on the anticancer activity of several ruthenium (Ru) complexes with quinolone antibiotics in colon tumor cell cultures. Real-time cell analysis and drug-mediated cytotoxicity tests monitored the inhibitory effects in the drug-treated LoVo colon cancer cells. Flow cytometry assays were performed to evaluate cell cycle phases distribution and apoptotic events. The obtained results showed dose-dependent increased levels of cell lysis and induction of apoptosis in LoVo cancer cells treated with the Ru(III) complexes. In addition, data showed a major decrease in cell proliferation, since the percentages of cells distributed in the S cell cycle phase diminished, and a G0/G1 cell arrest was observed. Therefore, our results strongly suggest that the newly synthesized Ru(III) complexes might play an important role in future chemotherapeutic approaches, since their activity is based on diminishing cell proliferation, induction of apoptosis, and modulation of cell cycle phases.
1. Introduction
Cancer is a major public health problem worldwide with an estimated 19.3 million new cancer cases and almost 10.0 million cancer deaths occurring in 2020. Moreover, in 2040, the global cancer burden is expected to be 28.4 million cases, a 47% rise from 2020, due to the increasing risk factors associated with globalization and a growing economy [1]. Colorectal cancer is the third malignancy between the most common human cancers in the world, and it is also among the most diagnosed cancers in the United States and Europe for both men and women, with high incidence and mortality (10% of cancer diagnosed female patients, and 9.4% of cancer diagnosed male patients) [1,2]. Many studies showed that cancer is a multi-step process consisting of several stages of development, such as the transformation of normal cells, tumor progression, and advanced metastasis [3], and it involves the loss of genomic stability that promotes the acquisition of further DNA alterations leading to cell transformation and tumor progression [4,5], aberrant progression of the cell cycle, resistance to growth inhibition, proliferation without dependence on growth factors, replication without limit, evasion of apoptosis, induction of angiogenesis, and modification of cell adhesion [3,6,7].
Although many types of cancers are initially susceptible to chemotherapy, over time, they can develop drug resistance that often leads to therapeutic failure. Resistance to chemotherapy and molecularly targeted therapies is a major problem facing current cancer research [8,9], and the number of drugs that have been shown to induce resistance in cancer cell killing is rapidly increasing, possibly through the modulation of survival cell components, such as proliferative or anti-apoptotic proteins [10,11,12]. The mechanisms of resistance to “classical” cytotoxic chemotherapeutics and to therapies that are designed to be selective for specific molecular targets share many features, such as alterations in the drug target, activation of pro-survival pathways, and ineffective induction of cell death [13,14,15].
The multi-drug combination chemotherapy was established to be a successful strategy in the management of colon cancer treatment, the reason being the potential additive or synergistic tumor cytotoxicity produced. Since the discovery of the cytotoxic properties of the cisplatin, platinum-based drugs have been investigated for cancer treatment and represent some of the most widely used anticancer chemotherapeutics, but their high toxicity and chemoresistance had spun the researchers to search for other novel chemotherapeutics based on metal compounds. In addition to cisplatin (CisPt), oxaliplatin and carboplatin are some of the most effective therapeutic agents commonly used in the treatment of colorectal cancers. [16,17]. The mechanism of action of CisPt has been linked to its ability to crosslink with the purine bases on DNA, and the CisPt-DNA adducts formed might interfere with normal DNA replication and transcription and subsequently promote the activation of cellular apoptosis [18,19].
Due to their potent cytotoxicity in cancer cells, ruthenium complexes have attracted great interest and recently emerged as potential candidates to replace platinum chemotherapy [19,20,21]. Over the last few years, research in colon cancer treatment has been focused on finding better chemotherapeutics. Therefore, various Ru complexes have been obtained and intensively studied due to their proper physicochemical and biochemical properties, such as selective bioactivity as well as the ability to overcome the resistance encountered with platinum-based drugs, which is associated with low toxicity profiles [21,22].
Ruthenium (III) complexes have shown remarkable antitumor activity with various advantages over traditional platinum drugs, including potent efficacy, low toxicity, and less drug resistance. Research in this area has shown over the last decade that subtle changes in the structure often have a significant effect on the pharmacological properties of metal complexes [20,23].
In the last years, many research studies were focused on obtaining complexes with remarkable activity using quinolones molecules as ligands [24]. Quinolones represent a class of synthetic antibacterial agents that is widely prescribed for the treatment of infections in humans or for veterinary use. The bactericidal activity of these agents arises from their capacity to inhibit two bacterial key enzymes: DNA gyrase (a type II topoisomerase) and DNA topoisomerase IV [25,26].
While quinolone-based drugs were developed to inhibit the prokaryiotic DNA gyrase [27], the antineoplastic drugs have eukaryotic type II topoisomerase as their primary target [28]. Moving the activity of quinolone from antibacterial to anticancer activity [29] represents a current challenge considering the increasing resistance to chemotherapy and the severe side effects of anti-neoplastic agents.
To accomplish this goal, different synthetic strategies were approached. The first of these directions involves chemical modifications on the quinolone nucleus, either by disruption of the zwitterionic properties of the compounds (which can be achieved by modifying the C7 basic substituent, or the C3 carboxyl group, or both) or by increasing the number of aromatic/condensed rings [30]. The most successful example is represented by vosaroxin (formely voreloxin), which is a first-in-class anticancer quinolone derivative that inhibits topoisomerase II and induces selective double-strand breaks, G2 arrest, and apoptosis [31,32]. Vosaroxin passed Phase 2 clinical trials in acute myeloid leukemia (AML) and platinum-resistant ovarian cancer, while in a Phase III trial, the addition of vosaroxin to cytarabine offered clinical benefit to some patients with relapsed or refractory AML [33]. Vosaroxin displays some valuable pharmacokinetic properties such as the minimal metabolization and a lack of free radical production; in addition, it is not a substrate for the glycoprotein efflux pump, and its antineoplastic activity is independent of P53 function [34].
A second direction aiming to extend the scale of biological activities for a molecular entity with coordinating properties is to obtain metal complexes with selected metal ions. In this regard, a starting point could be the complexes of quinolone antibiotics with Ru(III) ion based on the success of the already recognized NAMI-A [35] and KP1019 [36]. So far, only few Ru(III) complexes with quinolone antibiotics were reported. In these complexes, the quinolone molecules bind to the metal ions in two ways: as monoanionic bidentate generating metal chelates or as neutral unidentate ligand through the nitrogen atom of piperazin-1-yl moiety. The chelate complex of Ru(III) with norfloxacin of general formula [Ru(nor)3] (nor = norfloxacinate, C16H17FN3O3−) remarkably enhances the water solubility of norfloxacin and interacts with human serum albumin [37], while the chelate of ciprofloxacin [Ru(C17H17FN3O3)3]·4H2O is fully water soluble and stable under physiological pH for at least 48 h, and it interacts with ct-DNA [38]. The complexes of Ru(III) with unidentate ligands levofloxacin (LEV), gatifloxacin (GAT), and pipemidic acid (Pip-H: L) with formulas [Ru(LEVO)3(Cl)3]·8H2O [39], [Ru(GAT-NH4)(Cl)3(H2O)2] [40], and [M(L)3(Cl)3]·6H2O, respectively, were prepared and physicochemically characterized [41]. The biological evaluation of the Ru(III) complex with pipemidic acid revealed antibacterial efficiency against four kinds of bacteria—G(+) (Staphylococcus epidermidis and Staphylococcus aureus) and G(−) (Klebsiella and Escherichia coli)—and inhibitory activity against the human breast cancer (MCF-7) tumor cell line [41].
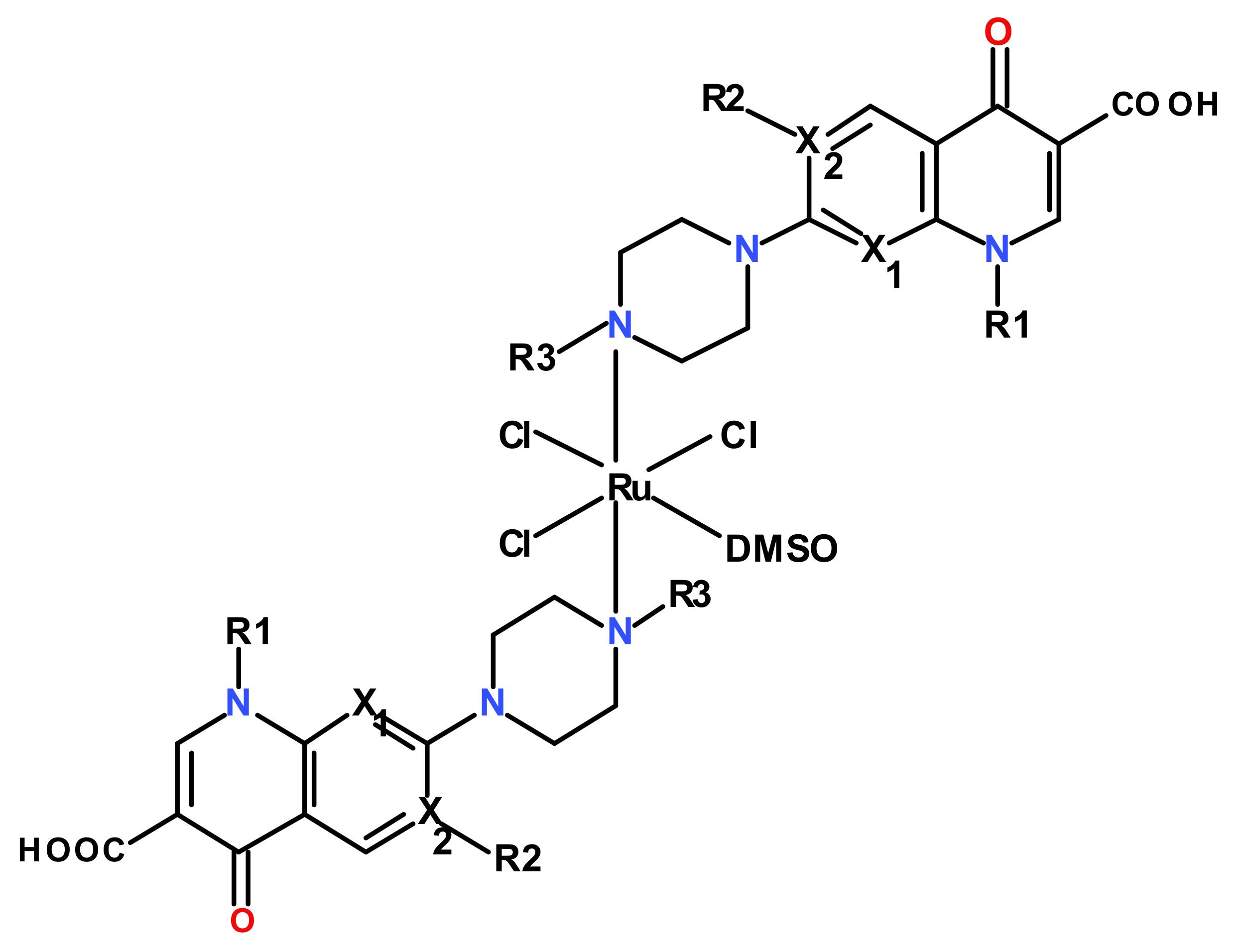
During previous research studies, our group also synthesized and characterized a series of some novel ruthenium (III) complexes with mixed ligands of the type RuCl3L2(DMSO)m·nH2O [42,43], as described in Table 1.

Table 1.
Ruthenium (III) complexes with mixed ligands of the type RuCl3L2(DMSO)m·nH2O.
All these complexes display an octahedral stereochemistry with the quinolone ligand acting as monodentate, coordinated through the N4 atom of the piperazinyl ring (Figure 1).
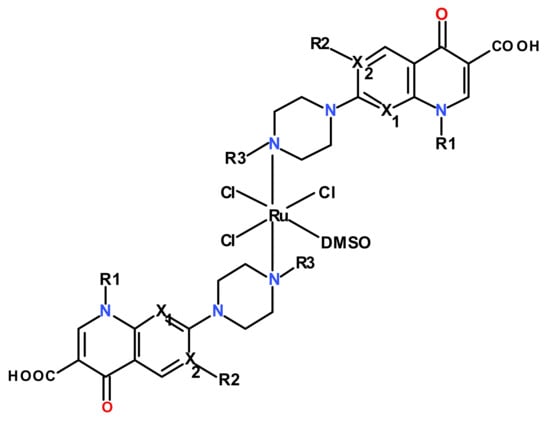
Figure 1.
Structures of the investigated Ru(III) complexes with quinolones.
Our previous studies showed that these Ru(III) complexes interacted in vitro with biomolecules such as calf thymus DNA [44] and human serum transferrin [45], and they presented antimicrobial activity in vitro [46] and antitumor activity in vivo [47]. In one previous study, the fluorescence intensity of the DNA-EtBr system was registered in the absence and in the presence of increasing amounts of complexes. The competitive study revealed the quenching of the fluorescence intensity of the DNA-EtBr system for all complexes, suggesting an intercalative mode for interaction with DNA [44]. The phenomenon is not singular, since other studies performed by our research group showed that organo-metal compounds induced cell lysis and apoptosis in several human cancer cell lines [48,49,50].
Apoptosis or “programmed cell death”, along with the cell cycle, represented important research directions addressed by our group, due to the implications that these biological processes could have in the evolution of neoplastic diseases and the therapeutic approaches in cancer. The evaluation of the apoptosis process and cell cycle phases, added by the characterization of related molecules, key regulators, in human cancer cells, was addressed by other previous in vitro studies performed by our group, which were carried out mainly in colon [7,11], renal [51], mammary [52], ovarian [53], or head and neck cells [54,55]. Moreover, due to the fact that often, the effect of cancer therapy could be amplified by combined treatments, in vitro approaches were made in testing the biological activity of several biocompounds as additional treatments to classical chemotherapy [7,11,52,55]. In addition, because the mode of administration of anticancer drugs and adjuvants contributes to increasing the therapeutic effect and diminishing the unwanted side effects, we studied the capacity of nanoparticles to better sensitize the tumor cells as well as modulate apoptosis and proliferation [56,57]. The last but not least research direction aimed to decipher the mechanisms underlying the drug resistance and find the means to reverse it in various cancer cell lines [7,55].
Therefore, all these together with the anticancer potential showed by Ru(III) complexes prompted us to extend our studies on the investigation of the in vitro anticancer biological activity of the Ru(III) complexes with some quinolone antibiotics in the human colon adenocarcinoma LoVo cell line, using as positive control the therapeutic drug cisplatin (CisPt).
2. Materials and Methods
2.1. Reagents
The Ru(III) complexes studied were previously synthesized and characterized [39,40]. The stock solutions were prepared by dissolving the Ru(III) complexes in a minimum amount of DMSO, which was diluted in the cosolvent system H2O:Polyethyleneglycol 400 (PEG400) 3.8:6.2 (V/V) and preserved at −20 °C.
Cisplatin (Cis-diammineplatinum(II) dichloride, DDP) was purchased from Sigma Aldrich (St. Louis, MO, USA), and stock solutions of 5 mM were prepared as recommended, in ultrapure sterile water, and preserved at −20 °C. Working solutions were prepared from the frozen stocks in culture medium before each experiment.
Ribonuclease A (RNase A) from bovine pancreas, propidium iodide (PI), ethylenediaminetetraacetic acid (EDTA), dimethyl sulfoxide (DMSO), and paraformaldehyde (PFA) were obtained from Sigma Aldrich (St. Louis, MO, USA). Annexin V-FITC Apoptosis Detection kit was purchased from Becton Dickinson (BD) Biosciences (Mountain View, CA, USA).
2.2. Cell Cultures and Treatments
The LoVo human colon cancer cell line was derived from a metastatic tumor in the left supraclavicular region of a 56-year-old male with Dukes’ type C, grade IV, colorectal adenocarcinoma, and it was purchased from the American Type Culture Collection (ATCC). Cells were routinely maintained in culture in DMEM/F12 medium added with 2 mM l-glutamine and 10% fetal calf serum in culture flasks and incubated at 37 °C in 5% CO2 humidified atmosphere [11]. After 24 h, when cells achieved around 60% confluence, they were treated for various periods of time with different concentrations of Ru(III) complexes or CisPt, as positive control. Then, cells from flasks were detached with a non-enzymatic solution of PBS/1 mM EDTA, washed twice in PBS, and immediately used for proliferation/cytotoxicity assays or evaluation of apoptotic events by flow cytometry; alternatively, cells were fixed in ice-cold ethanol/PBS (70:30) and kept until use at 4 °C for cell cycle analysis by flow-cytometry technique. Non-treated cells were used as controls throughout all experiments [11,52,55].
2.3. Real-Time Cell Analysis (RTCA) of Drug-Treated Cells
To analyze the proliferation profiles of treated LoVo cells, we continuously monitored cell growth by using real-time cells analysis (RTCA) assay and an xCELLigence DP-System, which allows cell-based, label-free in vitro assays and real-time monitoring of cellular processes such as cell viability and cytotoxicity [52,58]. Changes in a cell status, such as cell morphology, cell adhesion, or cell viability led to a change in cell index (CI), which is a quantitative measure of the cell number present in a well [7,48]. Real-time impedance data were obtained and used to generate compound-specific profiles that are dependent on the biological mechanisms of action of each Ru(III) complex. Briefly, LoVo human colon cancer cells were cultured in DMEM culture medium added by 2 mM l-glutamine and 10% FCS, and seeded in 100 μL culture medium in 16 cell E-Plates (ACEA Biosciences, San Diego, CA, USA). Growth curves started to be automatically recorded in real time on the xCELLigence System with a DP device [11]. After cells proliferated until a cellular index (CI) over 1.0, usually after 24 h, scalar concentrations of Ru(III) complexes or CisPt were added, and growth curves were registered in real time on a computer using RTCA 2.1.2. Software [7,52,57].
2.4. Assessment of Drug Cytotoxicity by MTS Assay
All assays were performed in triplicate in 96-well microtiter plates with flat-bottom (Falcon), using a CellTiter 96 Aqueous One Solution Cell Proliferation Assay (Promega), which is a MTS colorimetric assay. The method is based on the ability of metabolically active cells to reduce MTS, which is a yellow tetrazolium salt to the colored formazan that is soluble in the culture medium. Briefly, 1.5 × 104 cells/well were cultured in 100 μL for 24 h, culture supernatants were discarded; then, cells were treated for an additional 24 h with increasing concentrations of Ru(III) complexes or CisPt. After the end of the incubation time, 20 μL of reagent mixture were added in each well: (a) MTS[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt] and (b) PES (phenazine ethosulfate), which has a high chemical stability, and might be combined with MTS to form a stable solution.
After adding the coloring mixture, plates were incubated for 4 h at 37 °C with mild agitation every 20 min. The color that developed during incubation was spectrophotometrically quantified at λ = 492 nm, using a Dynex ELISA reader (DYNEX Technologies—MRS, USA) [48,50,54,57].
The percentages of cell lysis were calculated using with the equation below, and compared to untreated cells (considered 100% viable):
where T = absorbance of treated cells, U = absorbance of untreated cells, and B = absorbance of culture medium (blank), for λ = 492 nm.
Cell lysis (%) = 100 − (T − B)/(U − B) × 100
Cell lysis data were expressed as the mean values ± standard deviations (SD) of three different experiments. A parallel assay for the evaluation of DMSO cytotoxicity was determined in the same experimental conditions, but no cell cytotoxicity was observed for concentrations lower than 1% (data not shown).
2.5. Apoptosis Analysis by Flow Cytometry
The apoptosis assay was carried out using the Annexin V-FITC Apoptosis Detection Kit and the manufacturer’s protocol from Becton Dickinson (BD) Biosciences. The percentages of apoptotic cells were evaluated by double staining with Annexin V-FITC/propidium iodide (PI), followed by sample analysis by flow cytometry. As negative control of the test, the unmarked cells were used [11,52]. Briefly, 1 × 105 cells per tube were resuspended in 100 μL binding buffer, added with 5 μL Annexin V/FITC and 5 μL PI solution, and then incubated for 15 min/RT at dark. Then, 400 μL binding buffer were added and percentages of apoptotic events were measured by using data acquisition by flow cytometry using a FACScan or FACS CantoII cytometers (Becton Dickinson, Immunocytometry System, Mountain View, CA, USA). Evaluation of apoptotic events was performed using WinMDI 2.9 and Kaluza softwares [11,52].
2.6. Cell Cycle Evaluation Using Flow Cytometry
Previously, ethanol fixed cells (1 × 106) were washed twice in PBS, and cell pellets were resuspended in 350 μL of PBS and then added with 50 μL of 10 mg/mL RNase A (1 mg/mL final concentration). After 10 min incubation at 37 °C, 100 μL of 100 mg/mL PI solution were added (20 μg/mL final concentration) and further incubated at 37 °C for 10 min. The probes were kept in the dark at 4 °C until data acquisition by flow cytometry using a FACS CantoII flow-cytometer (Becton Dickinson (BD) Immunocytometry System, Mountain View, CA, USA). Then, cell cycle analyses were performed using ModFIT LT software to estimate the nuclear DNA content in 3 × 104 nuclei and progression through cell cycle phases [11,55].
2.7. Statistical Analysis
All measurements were performed in triplicates. Statistical analyses were performed using one-way analysis of variance (ANOVA) and GraphPad Prism 7 software (GraphPad Software Inc., La Jolla, CA, USA).
3. Results and Discussion
3.1. Real-Time Cell Analysis of Proliferation of LoVo Cells Treated with Ru(III) Complexes
The xCELLigence System of Real-Time Cell Analyzer (RTCA) allows label-free and real-time monitoring of cellular processes such as cell proliferation, cytotoxicity, adhesion, viability, invasion, and migration, using electronic cell sensor array technology. Cellular events are monitored in real time without the incorporation of labels by measuring the electrical impedance across inter-digitated micro-electrodes integrated on the bottom of special tissue culture plates. The actual kinetic response of the cells within an assay provided important information regarding the biological status of the cell. Changes in cell status, such as cell morphology, adhesion, or viability led to a change in cell index (CI), which is a quantitative measure of the cell number present in a well [58].
The capacity of cisplatin (CisPt), a metallic anticancer drug that has been used for many years to treat various human cancers, including colon cancer, and Ru(III) complexes to inhibit cell proliferation was investigated in a LoVo colon cancer cell line in order to modulate the chemosensitivity of colon cancer cells to drug treatments and overcome the chemoresistance. Therefore, we monitored compound-induced cytotoxicity to produce continuous compound- and concentration-dependent cell impedance profiles using the xCELLigence System and LoVo cancer cell line as our in vitro model.
To assess the optimal number of cells to be used in further assays, in a first line of experiments, LoVo colon cancer cells were seeded at scalar densities from 103 to 106/well in 16-well E-Plates for a dynamic monitoring of adherent cell proliferation. The cell growth curves were automatically recorded on the xCELLigence System in real time. RTCA growth curves of LoVo cells and CI values prompted us to establish the optimal cell density to be used in further experiments and choose the 15 × 103 cells/well for the Ry(III) compound-mediated cell cytotoxicity assays (data not shown).
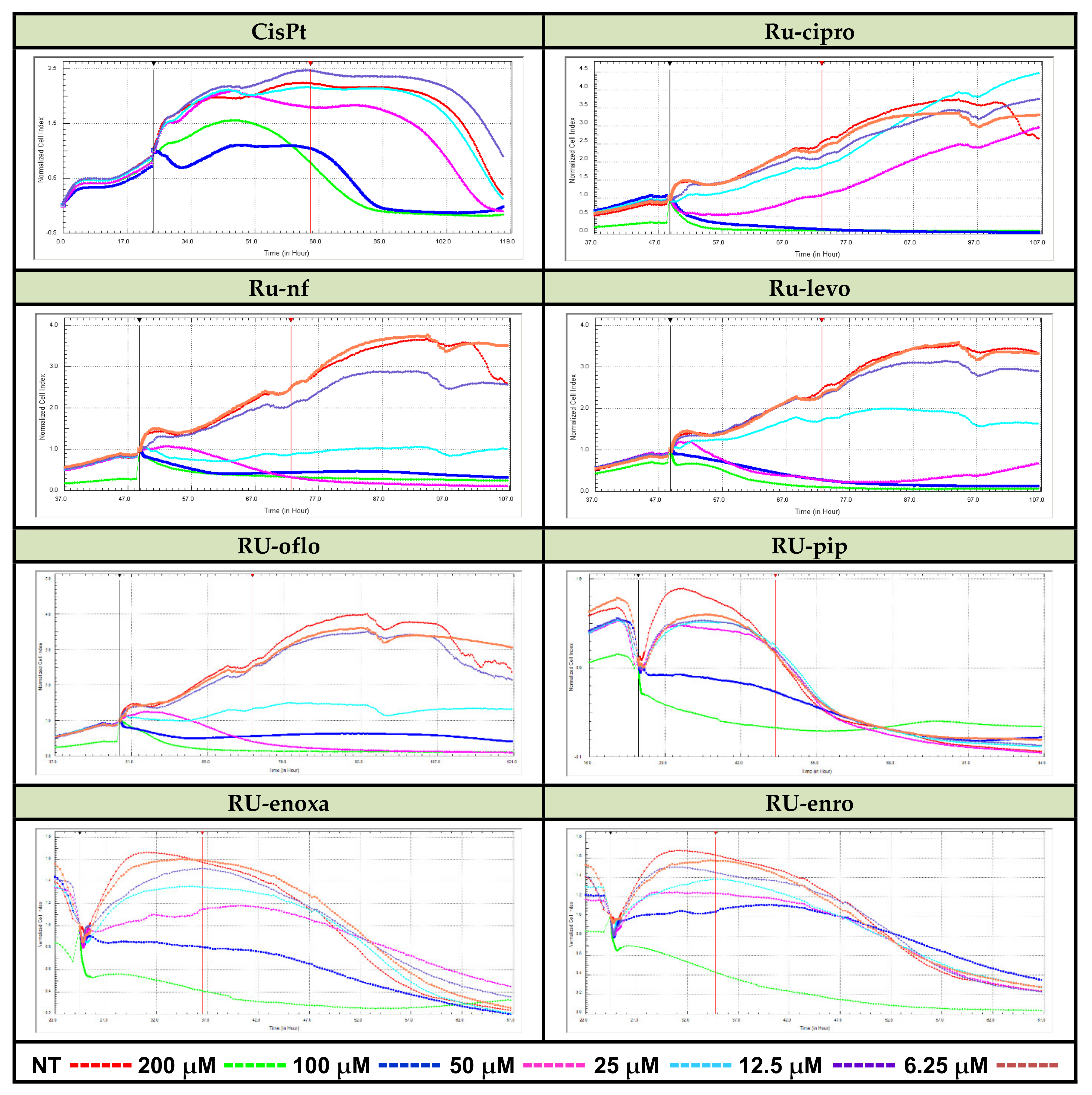
LoVo cancer cells were seeded in E-plates 16 in complete culture medium (1.5 × 104 cells/well/100 μL), plates were put in the xCELLigence DP device and cultured at 37 °C in a 5% CO2 humified atmosphere. After 24 h, cells were added by increasing concentrations of Ru(III) complexes or CisPt (from 12.5 to 200 μM), and growth curves were registered in real time on a computer by using RTCA 2.1.2. Software. Then, the cell growth, proliferation, and viability of LoVo cells were continuously monitored. Results are expressed as normalized cell index (CI) after automatic comparison between the curves of viability for treated vs. non-treated (NT) cells (Figure 2).
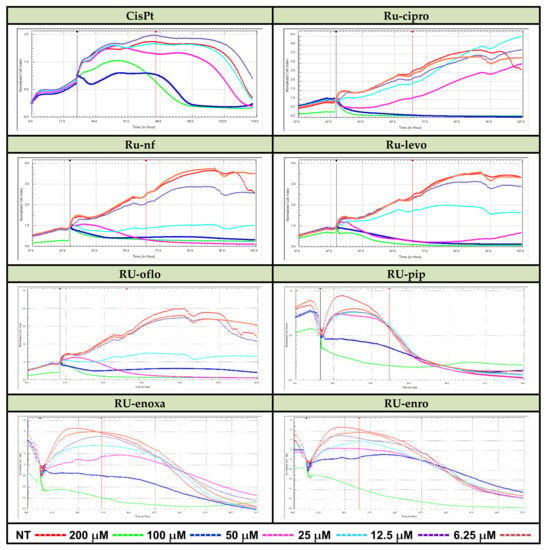
Figure 2.
Real-time cell analysis of cytotoxic effects of Ru(III) complexes vs. CisPt. LoVo human colon cancer cells were treated with the reported drug concentrations in 16-well E-plates. Monitoring of Ru(III) complexes-mediated cytolysis was performed by measuring the cell index for several days using the xCELLigence DP-System and compared to CisPt-induced inhibition of proliferation vs. cell lysis.
Some active ruthenium complexes could manifest cytotoxic action against various cancer cell lines, including colon cancer cell lines [16,17,20]. Therefore, we investigated the proliferative vs. cytotoxic action of Ru(III) complexes on the LoVo colon cancer cell line, and results varied depending on the type of compound, concentration, and treatment time. RTCA analysis of the LoVo cell line proliferation curves for cells treated with concentrations between 6.25 and 200 μM of Ru(III) complexes, and normalized vs. control (the proliferation curve for untreated cells), demonstrated a pronounced cytotoxicity decrease in viability for almost all the concentrations used in the assay, mostly for 25, 50, 100, or 200 μM (Figure 2).
Among the tested Ru(III) complexes, Ru-nf, Ru-oflo, and Ru-levo seem to induce the highest inhibition of LoVo cell proliferation during the first 24 h of treatments with 50, 100, or 200 μM of drugs. Ru-pip, Ru-enro, and Ru-enoxa showed also inhibitory effects on cell proliferation, the inhibitory effect becoming stronger with increasing treatment time.
Real-time impedance data obtained were used to generate compound-dependent cell impedance profiles that are dependent on the biological mechanisms of action of each compound. The proper concentrations of compounds with cytotoxic or cytostatic potential to inhibit the growth of LoVo adherent tumor cells were further used in end-point assays such as the evaluation of cell proliferation by colorimetric MTS assay or flow-cytometry approaches to investigate both apoptosis induction and DNA progression through cell cycle phases.
3.2. Cell Lysis Induced by Ru(III) Complexes Is Dose-Dependent
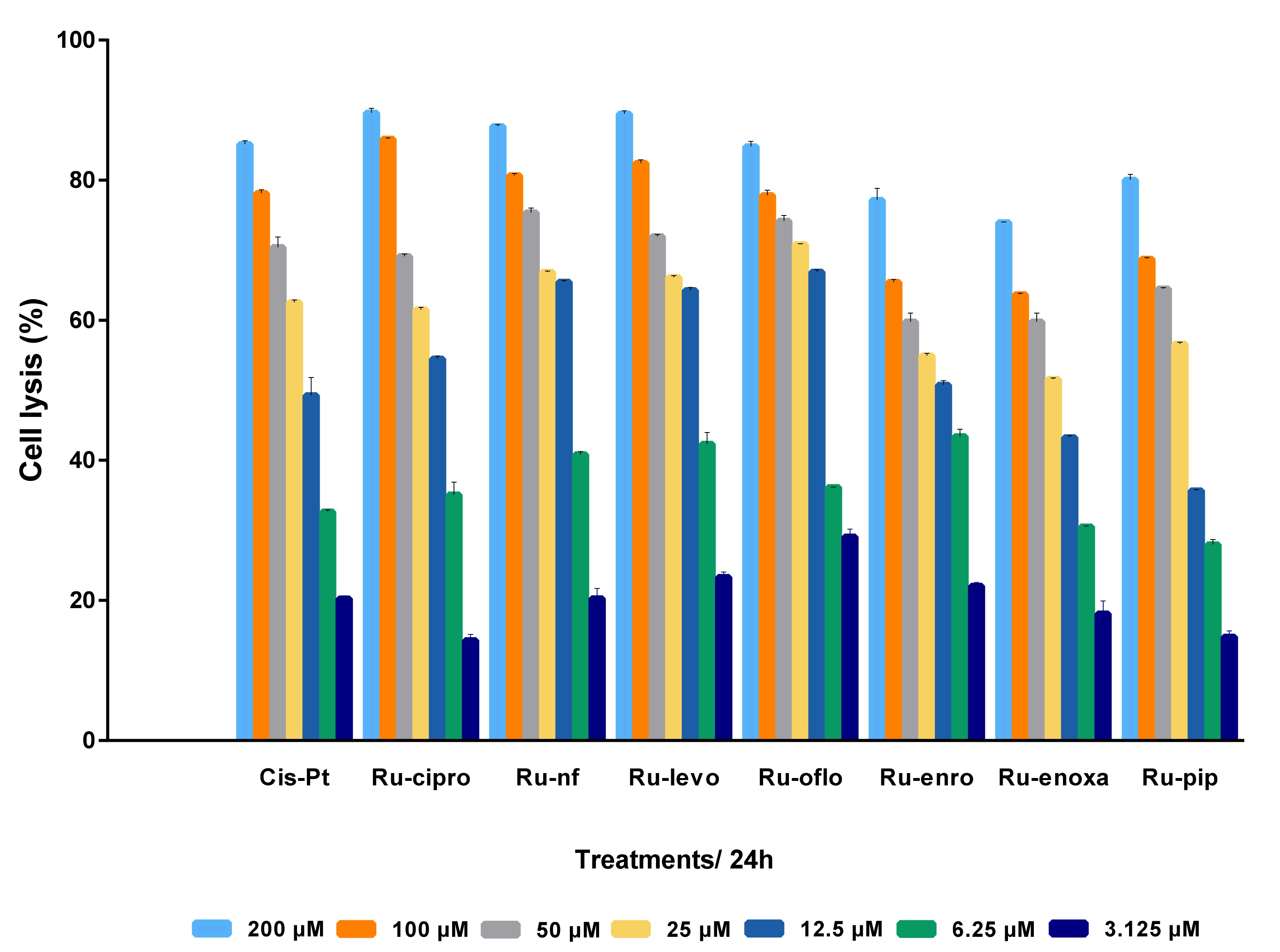
After analyzing the effects of various concentrations of Ru(III) complexes on LoVo human colon cancer cell proliferation, and compared to cisplatin, in the real-time cell analysis assay using the xCELLigence DP-System, cells were subjected to an end-point, colorimetric drug-mediated cytotoxicity method. MTS assay was used in order to better assess the levels of cell lysis induced by drug treatments and discriminate between the Ru(III) complexes under study regarding their ability to kill the LoVo colon tumor cells. Increasing concentrations of Ru(III) complexes with quinolones or CisPt ranging from 3.125 to 200 μM were used to treat LoVo cultured cells for 24 h. Percentages of cell lysis induced by drug treatments in the LoVo cell line are shown in Figure 3. Untreated cells were used as control and considered to have 100% viability. Results were expressed as mean values of three different experiments ± standard deviation (SD).
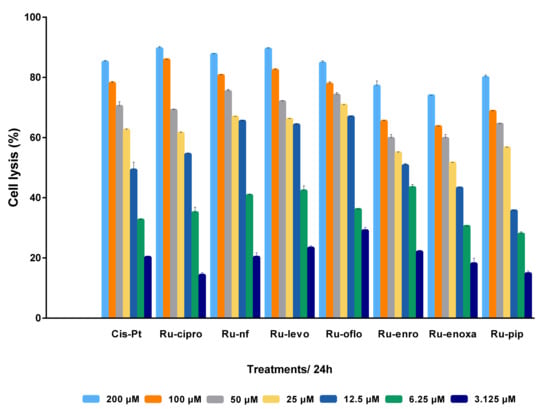
Figure 3.
Antitumor effect of Ru(III) complexes against LoVo colon cancer cells. Cells lysis was induced by 24 h treatment with scalar concentrations of Ru(III) complexes or CisPt, and compared to untreated control LoVo cells. Data shown are expressed as mean values ± standard deviations (SD) of three different experiments (n = 3).
Analyzing the cell response to drug treatments, a cytotoxic dose-dependent effect was observed for all Ru(III) complexes and CisPt. Treatments for 24 h with Ru(III) complexes in cosolvents resulted in increased cell lysis rates and implicitly decreased cell viability. Thus, more than 75% of the cells were killed when the LoVo cell line was incubated either with 200 μM or 100 μM of Ru-nf, Ru-levo, Ru-oflo, Ru-cipro, or CisPt. Treatments with 25 or 50 μM drugs induced more than 50% of cell lysis for all the Ru(III) complexes under study as well as CisPt. Even for 6.25 μM or 12.5 μM treatments, the percentages of cell lysis were higher than 25% for all drugs tested, while 3.125 μM induced more than 14% cell lysis (Figure 3).
In addition, the inhibitory concentration values (IC) of Ru(III) complexes or CisPt were calculated using the linear regression analysis from the cytotoxicity data of each compound and for different percentages of cell lysis corresponding to drug concentrations.
Therefore, the corresponding drug concentrations required to induce the lysis of 50% (IC50), 75% (IC75), or 90% (IC90) of the cells were calculated when the cytotoxicity of Ru(III) complexes was assessed by MTS assay after 24 h of their incubation with LoVo cells compared to cisplatin as positive control (Table 2).
Table 2.
Values (μM) of IC50, IC75, and IC90, for Ru(III) complexes and CisPt-mediated lysis.
In terms of IC50 values, several Ru(III) complexes with quinolones displayed similar activities to the positive control, cisplatin, on LoVo cells. The best cytotoxic activity seemed to have Ru-cipro, Ru-nf, Ru-levo, and Ru-oflo since IC50, IC75, and IC90 values were the lowest (Table 2), and therefore, the toxicity profile is lower compared to cisplatin, which might allow a wide modulation of concentrations to be further used in therapeutic approaches with an improved control of the non-target cell toxicity.
The results are similar to those obtained by in vitro treatments with several other ruthenium compounds such as dinuclear Ru(III) complexes, whose effect was studied on different types of cancer cell lines, and when a reduction of cell proliferation was observed after 24 h of exposure to 100 μM of the compounds under study [16]. In addition, some ruthenium organometallic compounds might present collateral sensitivity for cisplatin-resistant cancer cells [59].
Data analysis following the MTS colorimetric assay confirms the results obtained by RTCA analysis. Thus, in subsequent end-point assays, such as apoptosis and DNA progression through cell cycle phases, LoVo cells will be treated with fixed concentrations of 10 μM and 50 μM of Ru(III) complexes.
3.3. Promotion of Apoptosis by Ru(III) Complexes in LoVo-Treated Cells
Apoptosis, known also as “programmed cell death”, is a cellular “suicide” mechanism responsible for the control of the cell number in tissues and elimination of those predisposed to malignant transformation that present DNA mutations or have an aberrant cell cycle [60,61,62,63]. During tumorigenesis, apoptosis is undermined through the systematic loss of regulatory control mechanisms, finally resulting in the generation of a malignant phenotype, and resistance to chemo- and radio-therapy [8,64,65]. The mechanisms of apoptosis might be of great importance for carcinogenesis, tumor evasion, and have practical implications for anticancer therapy, since many anticancer drugs act during the physiological pathways of apoptosis, in the end leading to tumor cell destruction [66,67,68,69,70].
Along with the inhibition of cellular proliferation, apoptosis is frequently evaluated in the biological response to treatment with various chemotherapeutic agents. After automatic data acquisition from the homogenous assays of real time of the compound-specific influence, and further of the end-point test of drug-mediated cytotoxicity, we calculated IC50 values and determined the optimal doses to be used in further end-point experiments, such as evaluation of apoptosis induction.
In order to study the potential role of Ru(III) complexes in the induction of apoptosis in colon cancer cells, the levels of apoptotic events were evaluated by flow cytometry techniques. LoVo colon cancer cells were cultured in complete medium for 24 h; then, the culture medium was changed, and cells were treated for an additional 24 h with 10 or 50 μM of Ru(III) complexes. Then, LoVo cells were detached with PBS/1 mM EDTA, sequentially washed with PBS and washing buffer, and centrifuged at 300× g/5 min. Pellets were suspended in 400 μL binding buffer, and 100 μL was distributed in flow tubes and stained with 5 μL of Annexin-V/FITC and/or PI for 15 min/RT in the dark [7,55]). As positive controls, cells treated with 10, 25, or 50 μM cisplatin were used, while the negative control (CTRL) was represented by untreated cells. The green and red fluorescences were measured by using a FACScan flow-cytometer (BD Biosciences) after acquiring 10,000 events. Then, the acquired data were analyzed using WinMDI2.9 and Kaluza softwares, and apoptotic events were expressed as cell percentages.
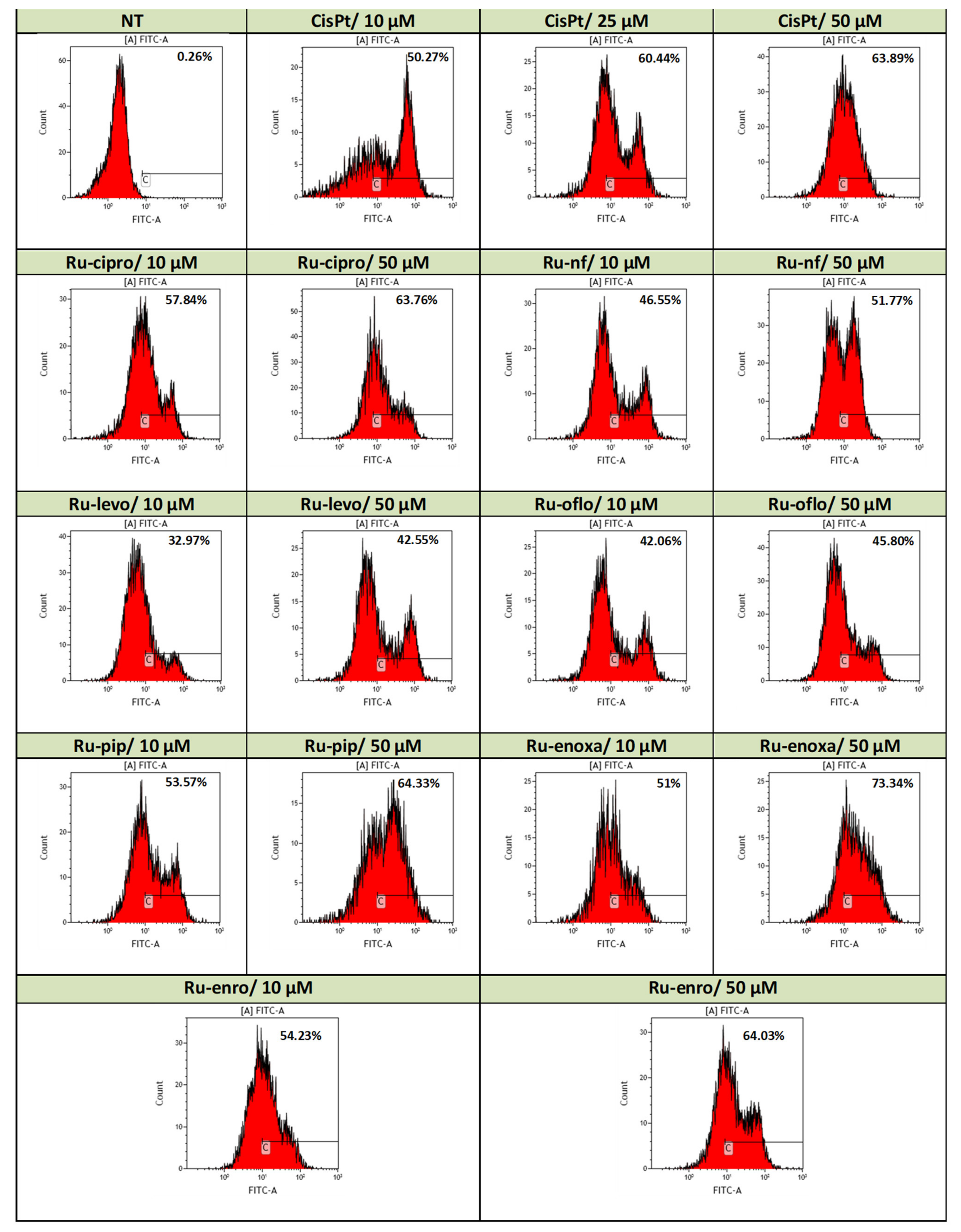
Figure 4 shows data from a significant experiment of induced apoptosis in LoVo cells by 24 h treatments with either Ru(III) complexes or CisPt compared to untreated cells. Total apoptosis took into account the labeled Annexin-V/FITC cells, and data are presented as histograms, the assay marker of positivity being set on non-treated cell champion.
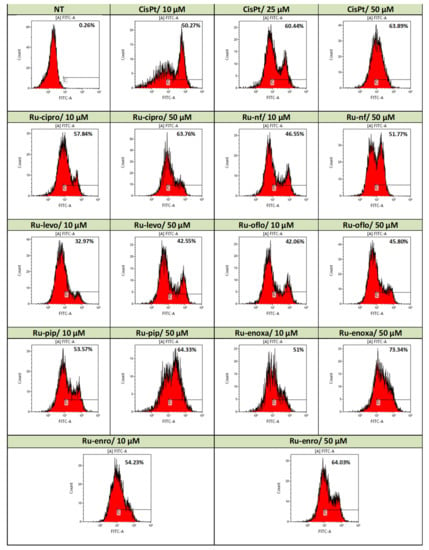
Figure 4.
Induction of apoptosis in the LoVo cell line by Ru(III) complexes vs. CisPt. Effects of drug treatments on apoptosis induction in the LoVo human colon adenocarcinoma cell line were evaluated by Annexin-V/FITC and PI double-staining assay. Levels of apoptotic events were assessed by flow cytometry acquisition using a FACScan cytometer and analysis with WinMDI2.9 and Kaluza softwares. Data are presented as flow cytometry histograms.
Annexin V belongs to a family of calcium-dependent phospholipid-binding proteins that bind to phosphatidylserine (PS) and allow identifying the apoptotic cells found in early and late stages, the last process preceding necrosis; propidium iodide (PI) is a dye used for assessing cell viability and the exclusion of non-viable cells that binds to double-stranded DNA but is excluded from cells with intact plasma membranes. It is used in combination with Annexin V/FITC to assess the cell viability and discriminate between apoptotic and necrotic cells; live cells do not bind Annexin or PI.
Use of 24 h treatments with 10 μM of all Ru(III) complexes with quinolones induced higher levels of apoptosis compared to control cells with over 32% of apoptosis being found in the treated cells (Figure 4). Ru-cipro, Ru-pip, Ru-enoxa, and Ru-enro induced levels of apoptosis of 57.84%, 53.57%, 51%, and 54.23%, respectively, higher than the apoptotic events induced by 10 μM CisPt treatment (50.27%). The lowest apoptosis was induced by Ru-levo (32.97%), while Ru-oflo and Ru-nf induced more than 40% of apoptosis, 42.06% and 46.55%, respectively.
When drug doses of cell treatments were increased to 50 μM, stronger effects were observed both for Ru(III) complexes and CisPt treatments (Figure 4). Treatments with Ru-cipro, Ru-pip, and Ru enro induced levels of apoptosis comparable with those induced by CisPt treatment (63.76%, 64.33%, and 64.03%, respectively, as compared to 63.89%) (Figure 4). The strongest effect of 73.34% apoptosis was induced by Ru-enoxa treatment. However, a high increase in the total apoptosis was also induced by Ru-nf (51.77%), Ru-levo (42.55%), and Ru-oflo (45.80%) (Figure 4).
Induction of apoptosis was also achieved by cancer cell treatments with several other ruthenium (II) and (III) compounds, but levels of apoptotic events depended not only on ligands but also on the doses used and the sensitivity of the cancer cell lines tested [16,22].
3.4. Cell Cycle Progression Changes in LoVo Cells Treated with Ru(III) Complexes
The cell cycle is the ordered sequence of events that occur to enable DNA replication and cell division, and it can be divided in two phases (interphase and mitosis) and four stages. During these stages, the cell increases in size (gap 1/G1 stage), copies its DNA (synthesis, or S stage), prepares to divide (gap 2, or G2 stage), and then divides (mitosis or M stage). The stages G1, S, and G2 belong to the interphase, which accounts for the span between cell divisions. Some cells exit the G1 stage and enter in a resting state called G0. Based on stimulatory and inhibitory messages received by a cell, it “decides” whether it should enter into the cell cycle and divide. Regulation of the cell cycle machinery, resulting in an arrest of cells in different phases of the cell cycle, might be altered by anticancer agents, and thereby, the growth and proliferation of cancerous cells could be reduced [16,19,71].
To reveal the anti-carcinogenic effects of Ru(III) complexes with quinolones that may have an inhibitory potential, cells were cultured and treated as in the experiment described above, and the end-point assay of LoVo cell progression through cell cycle phases was studied by flow cytometry. Perturbations in cell cycle progression may account for the anti-carcinogenic effects. Flow cytometry assays are useful techniques that became standard tools to analyze the DNA content of cells and show the cell cycle distribution.
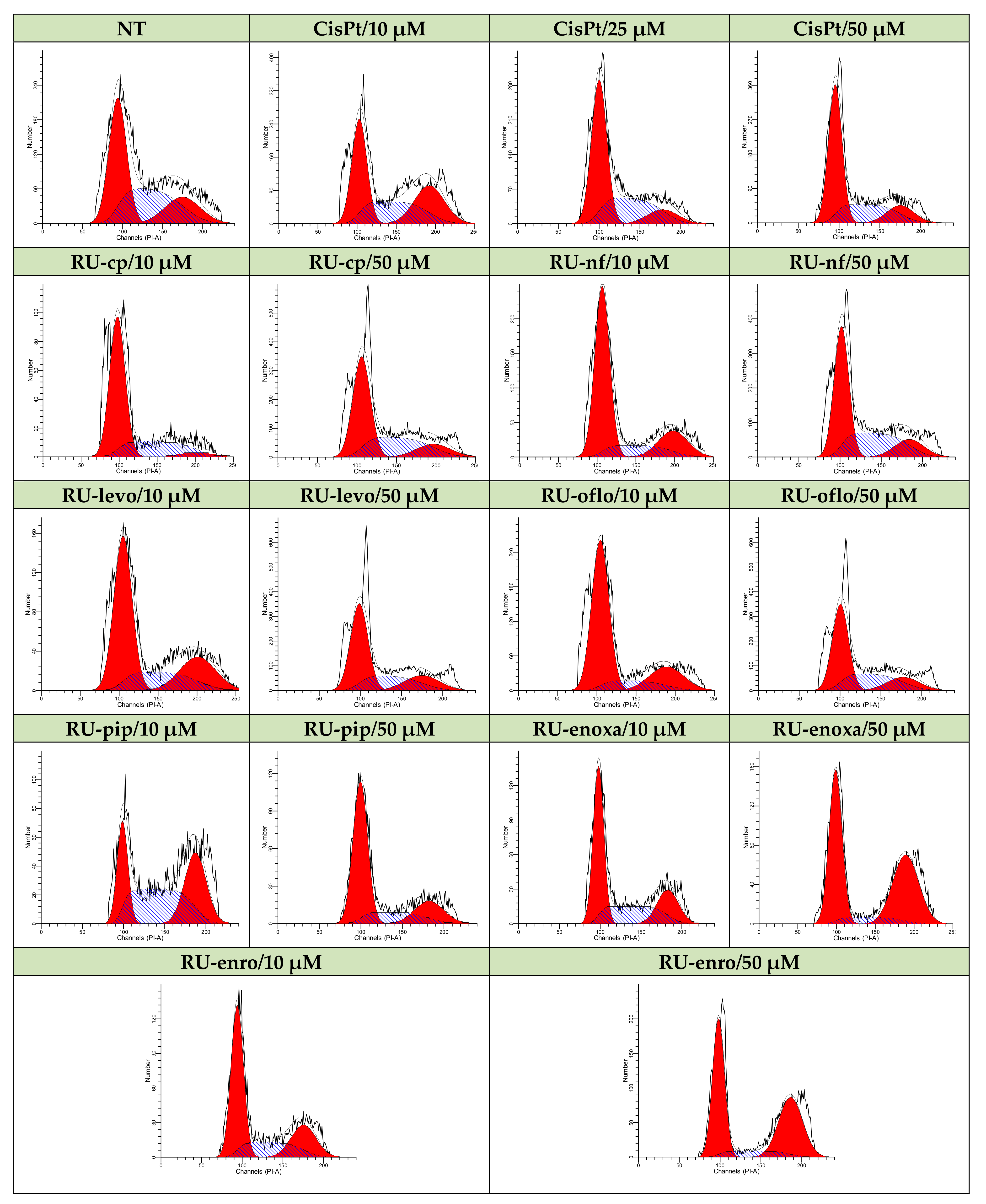
Therefore, non-treated control LoVo cells or cells treated either with Ru(III) complexes or CisPt were further subjected to RNase treatment, PI labeling, followed by data acquisition and analysis by flow cytometry. DNA progression through cell cycle phases is shown in Figure 5 for all drug-treated samples vs. control, which is represented by non-treated cells. The nuclear DNA content and progression through cell cycle phases were evaluated in acquired 3 × 104 nuclei.
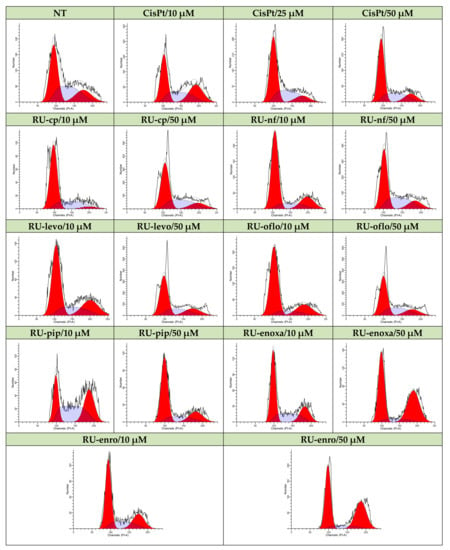
Figure 5.
Modulation of LoVo cell cycle progression by Ru(III) complexes vs. CisPt. Effects of treatments with Ru(III) complexes on the modulation of cell cycle phases was studied in the LoVo human colon adenocarcinoma cell line and compared to CisPt-treated or non-treated cells. DNA progression through cell cycle phases was evaluated by PI labeling, which was followed by flow cytometry acquisition and analysis using a FACS CantoII flow-cytometer (BD Biosciences) and ModFit software.
4. Discussion
The control cells displayed a cell distribution of 44.55% in G0/G1, 36.79% in S, and 18.66% in G2M cell cycle phases. Both 10 μM and 50 μM CisPt treatments induced a decrease in LoVo cell proliferation, since cell distribution in the S phase diminished to 33.2% and 28.99%, respectively, and induced a cell cycle arrest in the G0/G1 phase, where 55.84% and 56.66%, respectively, of the nuclei were found (Figure 6, Table 3).
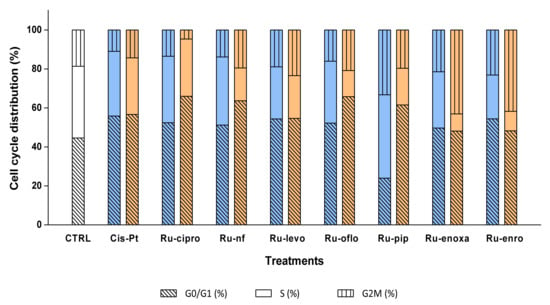
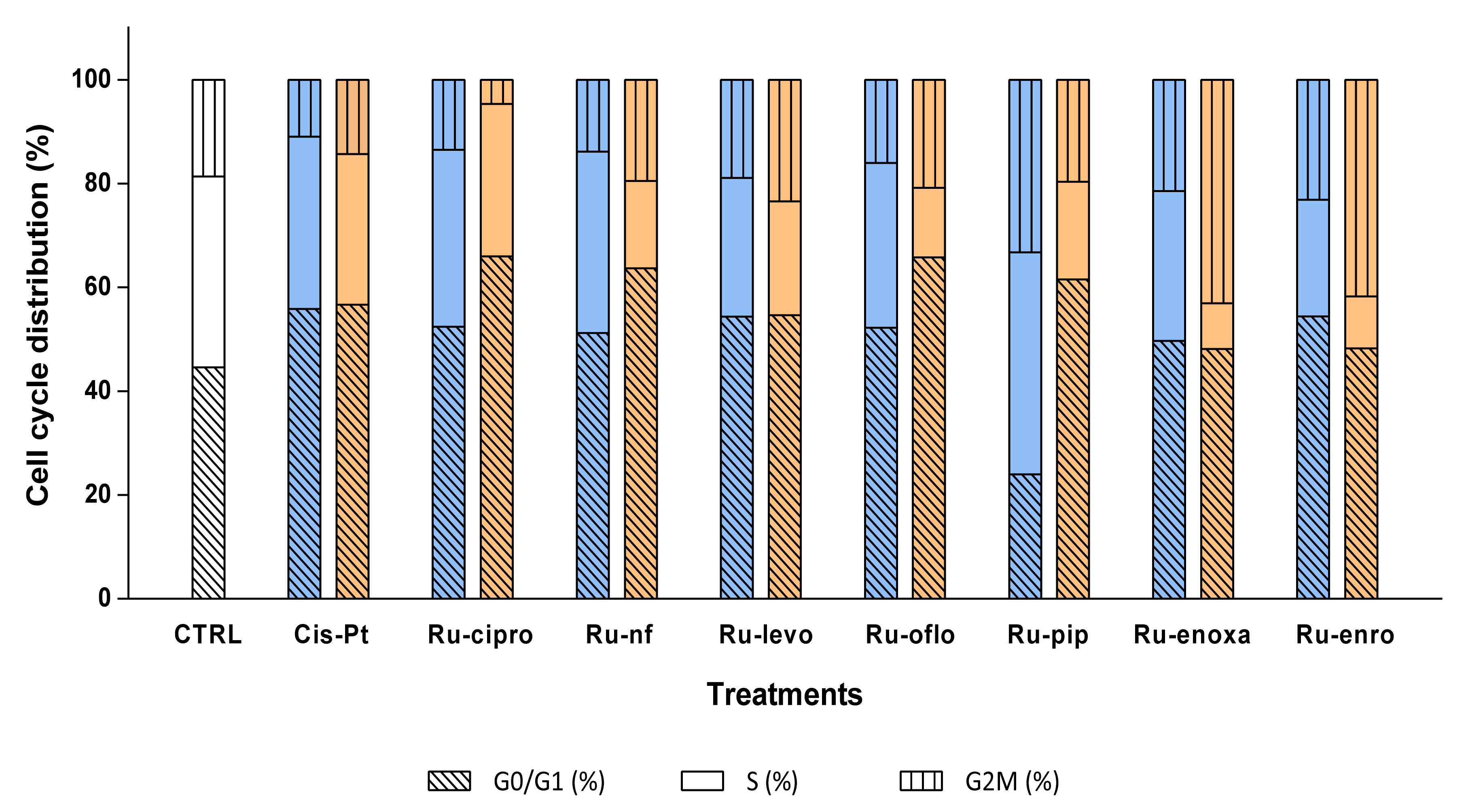
Figure 6.
Effects of Ru(III) complexes on DNA progression through LoVo cell cycle phases. Cell cycle phases were analyzed in LoVo human colon adenocarcinoma cell line after cell treatments with 10 μM (□) or 50 μM (□) Ru(III) complexes or CisPt. DNA progression through cell cycle phases was evaluated after RNase treatment and PI labeling, which was followed by flow cytometry acquisition using a FACS CantoII flow cytometer, and analysis with ModFit software. Negative control (CTRL) was represented by untreated LoVo cells.
Table 3.
Evaluation of DNA progression through LoVo cell cycle phases treated either with Ru(III) complexes or CisPt.
Cell treatments with 10 μM of Ru-cipro and Ru-nf induced similar decreases in the S phase as CisPt (34.1%, and 34.94%), while Ru-oflo, Ru-levo, Ru-enoxa, and Ru-enro induced a decrease in cell distribution in the S phase between 31.7% and 22.47%. At the same time, percentages of nuclei blocked in the G0/G1 phase were between 49.7% for Ru-enoxa and 54.40% for Ru-enro (Table 3, Figure 6).
When cells were treated with 50 μM of Ru(III) complexes, a higher decrease in the S phase distribution was observed for all treated samples. So, the highest effect was observed for Ru-pip (18.82%), Ru-nf (16.77%), Ru-oflo (13.43%), Ru-enro (9.97%), and Ru-enoxa (8.81%) treated cells. Treatments with 50 μM of Ru-cipro, Ru-nf, Ru-oflo, and Ru-pip increased also the cell distribution of over 60% in the G0/G1 phase, while Ru-enoxa and Ru-enro induced also an increase in the G2M phase, to 43.09% and 41.75%, respectively (Table 3, Figure 6).
When the proliferation indexes were calculated as the sum of the percentages of cells distributed in the S and G2M cell phases, the highest decreases were found for 50 μM treatments with Ru-cipro (33.98%), Ru-nf (36.31%), Ru-oflo (34.24%), and Ru-pip (38.49%) as compared to the non-treated cells (55.45%) or CisPt (43.34%). In addition, 10 μM treatments induced significant decreases of the proliferation index, which was mainly for Ru-cipro (47.59%), Ru-nf (48.82%), Ru-levo (45.62%), Ru-oflo (47.76%), or Ru-enro (45.61%). Both 10 μM and 50 μM of Ru-levo treatments showed similar proliferation indexes (Table 3, Figure 6).
In conclusion, increased concentrations of Ru(III) complexes seem to induce a dose-dependent decrease in cell distribution found in the S cell cycle phase. The results suggested that cell cycle arrest mostly took place in the G0/G1 phase and in a lower proportion also in the G2M phase, which might represent an important cause for the anti-proliferative effect of Ru(III) complexes, similar to CisPt, but hopefully less toxic.
The arrest in G0/G1 and G2-M phases are some cell cycle-regulating mechanisms related to cellular damage and, in particular, to DNA damage. In that case, DNA repair mechanisms are activated to ensure the replication of an intact DNA. When the reparation is successful, cells start again cycling; otherwise, apoptosis mechanisms are activated [16].
The results obtained during the study showed increased levels of cell lysis and apoptotic events, which are both dose-dependent in LoVo cancer cells treated with the Ru(III) complexes with quinolone antibiotics. In addition, data showed a major decrease in cell proliferation, since percentages of cells distributed in the S cell cycle phase diminished, and a G0/G1 cell arrest was observed. Therefore, our results strongly suggest that the newly synthesized Ru(III) complexes might play an important role in future chemotherapeutic approaches, since their activity is based on diminishing cell proliferation, induction of apoptosis, and modulation of cell cycle phases.
5. Conclusions
Intrinsic (de novo) or acquired tumor cell resistance remains a major challenge for different types of chemotherapeutical approaches. Despite the recent advances in surgery, radiotherapy, and especially in chemotherapy that have led to major achievements in cancer treatment, the development of tumor cell resistance to chemotherapeutic agents is still an important problem, reducing the effectiveness of available therapies. A major goal of early approaches in individualized therapy was to develop in vitro tests to predict tumors’ drug responsiveness. Since the chemosensitivity of tumors could not be predicted with high reliability, the evaluation of drug resistance is reliable independent of tumor type, test assay, and drug used in in vitro tests [72,73].
One of the most effective anticancer agents used to treat colon cancer is the platinum-based drugs, such as cisplatin, oxaliplatin, or carboplatin, but the insurgence of drug resistance and the high systemic toxicity developed are among the important obstacles against the success of chemotherapy. Ruthenium compounds have drawn great attention for their potential clinical application as anticancer drugs. Contrast data are available on their anticancer effects in colon cancer. The emerging interest appeared in developing new pharmaceutical compounds, such as Ru(III) complexes, with a high potential for chemotherapeutic use in cancer, and the improvement in their delivery prompted us to investigate their role in the modulation of colon cell proliferation vs. cytotoxicity, induction of apoptosis, and cell cycle regulation, in order to further find ways to improve their biological effects.
The Ru(III) complexes with quinolones developed by our group manifested a strong drug-mediated cytotoxicity against the LoVo colon cancer cell line, as assessed both by real-time cell analysis (RTCA) or end-point cytotoxicity assay, and results varied depending on the compound and concentration used. The registered RTCA proliferation curves of treated LoVo cells and the calculated IC50 values showed that Ru(III)-nf, -oflo, -levo, and -cipro were the most effective in killing colon cells at lower doses. Their toxicity profiles seemed to be lower compared to cisplatin; therefore, a wide modulation of concentrations might be further used in therapeutic approaches, and the control of non-target cell-toxicity could be improved.
Since apoptosis started to be considered as an ideal way to eliminate precancerous and/or cancer cells, and chemopreventive agents might modulate apoptosis, our experimental results obtained after evaluation of the apoptotic events induced by Ru(III) complexes with quinolones showed that treatments of colon cancer cells with the drugs under study induced dose-dependent effects, which could be useful in the management and therapy of cancer, since they proved to be at least as efficient as cisplatin. Treatments with 10 μM of all Ru(III) complexes with quinolones induced over 32% of apoptosis, while Ru-cipro, Ru-pip, Ru-enoxa, and Ru-enro induced higher levels of apoptotic events than CisPt. When drug doses of cell treatments were increased to 50 μM, stronger effects were observed both for Ru(III) complexes and CisPt treatments, the highest apoptosis being induced by Ru-enoxa treatment (73.34% apoptotic events).
Ru(III) complexes with quinolones seemed also to regulate the colon cell proliferation through cell cycle phases in a dose-dependent manner: both 10 and 50 μM treatments induced a decrease in the cells found in the synthesis (S) phase; in addition, we observed an increase in the cell cycle arrest mainly in the G0/G1 phase, the highest effect being obtained for Ru-pip, Ru-nf, Ru-oflo, Ru-enro, and Ru-enoxa treated cells.
Our results, consistent with other scientific reports highlighting the cytotoxic effects of ruthenium-based drugs in colon cells, showed that Ru(III) complexes with quinolone antibiotics might be further used to elaborate new therapeutic approaches for future personalized cancer treatments.
Author Contributions
Conceptualization, M.M. and V.U.; methodology, M.M., C.M.H., M.B., A.C.M., L.I.B., V.U.; investigation, M.M., C.M.H., M.B., L.I.B.; resources, M.M., L.I.B., V.U.; data curation, M.M., C.M.H., M.B., A.C.M., L.I.B., V.U.; writing—original draft preparation, M.M., C.M.H., M.B., A.C.M., I.A.V., L.I.B., V.U.; writing—review and editing, M.M., C.M.H., A.C.M., L.I.B., V.U.; supervision, L.I.B. and V.U.; project administration, M.M., L.I.B. and V.U.; funding acquisition, M.M., and V.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Research, the Executive Unit for Financing Higher Education, Research, Development and Innovation (UEFISCDI) through grants: PCCA PN-II 136/2012, and PN-III-P2-2.1-PED-2019-5143 (Contract PED 383/2020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Cirri, D.; Bartoli, F.; Pratesi, A.; Baglini, E.; Barresi, E.; Marzo, T. Strategies for the improvement of metal-based chemotherapeutic treatments. Biomedicines 2021, 9, 504. [Google Scholar] [CrossRef]
- Weber, G.F. Molecular Mechanisms of Cancer; Springer: Dordrecht, The Netherlands, 2007; Volume 39, pp. 45–54, 93–99. [Google Scholar]
- Solier, S.; Zhang, Y.W.; Ballestrero, A.; Pommier, Y.; Zoppoli, G. DNA damage response pathways and cell cycle checkpoints in colorectal cancer: Current concepts and future perspectives for targeted treatment. Curr. Cancer Drug Targets 2012, 12, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Temraz, S.; Mukherji, D.; Shamseddine, A. Potential targets for colorectal cancer prevention. Int. J. Mol. Sci. 2013, 14, 17279–17303. [Google Scholar] [CrossRef] [Green Version]
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T. Cancer statistics. CA Cancer J. Clin. 2008, 58, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Hotnog, C.M.; Mihaila, M.; Puiu, L.; Botezatu, A.; Roman, V.; Popescu, I.D.; Bostan, M.; Brasoveanu, L.I. Modulation of the interplay between p53, ICAM-1 and VEGF in drug-treated LoVo colon cancer cells. Rom. Biotechnol. Lett. 2019, 24, 261–270. [Google Scholar] [CrossRef]
- Gottesman, M.M. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [Green Version]
- Izar, B.; Rotow, J.; Gainor, J.; Clark, J.; Chabner, B. Pharmacokinetics, clinical indications, and resistance mechanisms in molecular targeted therapies in cancer. Pharmacol. Rev. 2013, 65, 1351–1395. [Google Scholar] [CrossRef] [Green Version]
- Mattern, J. Drug resistance in cancer: A multifactorial problem. Anticancer Res. 2003, 23, 1769–1772. [Google Scholar]
- Hotnog, D.; Mihaila, M.; Botezatu, A.; Matei, G.G.; Hotnog, C.; Anton, G.; Bostan, M.; Brasoveanu, L.I. Genistein potentiates the apoptotic effect of 5-fluorouracyl in colon cancer cell lines. Rom. Biotechnol. Lett 2013, 18, 8751–8760. [Google Scholar]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [Green Version]
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer 2012, 12, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving Paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Ke, X.; Shen, L. Molecular targeted therapy of cancer: The progress and future prospect. Front. Lab. Med. 2017, 1, 69–75. [Google Scholar] [CrossRef]
- Bergamo, A.; Stocco, G.; Gava, B.; Cocchietto, M.; Alessio, E.; Serli, B.; Iengo, E.; Sava, G. Distinct effects of dinuclear ruthenium(III) complexes on cell proliferation and on cell cycle regulation in human and murine tumor cell lines. JPET 2003, 305, 725–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhireksan, Z.; Davey, G.E.; Campomanes, P.; Groess, M.; Clave, C.M.; Yu, H.; Nazarov, A.A.; Yeo, C.H.F.; Ang, W.H.; Dröge, P.; et al. Ligand substitutions between ruthenium–cymene compounds can control protein versus DNA targeting and anticancer activity. Nat. Commun. 2014, 5, 3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, C.R.R.; Silva, M.M.; Quinet, A.; Cabral-Neto, J.B.; Menck, C.F.M. DNA repair pathways and cisplatin resistance: An intimate relationship. Clinics 2018, 73, e478s. [Google Scholar] [CrossRef] [PubMed]
- Bostan, M.; Mihaila, M.; Petrica-Matei, G.G.; Radu, N.; Hainarosie, R.; Stefanescu, C.G.; Roman, V.; Diaconu, C.C. Resveratrol Modulation of Apoptosis and Cell Cycle Response to Cisplatin in Head and Neck Cancer Cell Lines. Int. J. Mol. Sci. 2021, 22, 6322. [Google Scholar] [CrossRef]
- Zeng, L.; Chen, Y.; Liu, J.; Huang, H.; Guan, R.; Ji, L.; Chao, H. Ruthenium(II) complexes with 2-phenylimidazo[4,5-f][1,10]phenanthroline derivatives that strongly combat cisplatin-resistant tumor cells. Sci. Rep. 2016, 6, 19449. [Google Scholar] [CrossRef]
- Silvestri, S.; Cirilli, I.; Marcheggiani, F.; Dludla, P.; Lupidi, P.; Pettinari, R.; Marchetti, F.; Di Nicola, C.; Falcioni, G.; Marchini, C.; et al. Evaluation of the anticancer role of a novel ruthenium(II)-based compound compared to NAMI-A and cisplatin in impairing mitochondrial functionality and promoting oxidative stress in triple negative breast cancer models. Mitocondrion 2021, 56, 25–34. [Google Scholar] [CrossRef]
- Xu, L.; Zhong, N.J.; Xie, Y.Y.; Huang, H.L.; Jiang, G.B.; Liu, Y.J. Synthesis, characterization, in vitro cytotoxicity, and apoptosis-inducing properties of ruthenium(III) complexes. PLoS ONE 2014, 9, e96082. [Google Scholar] [CrossRef]
- Malecka, M.; Skocynska, A.; Goodman, D.M.; Hartinger, C.G.; Budzisz, E. Biological properties of ruthenium(II)/(III) complexes with flavonoids as ligands. Coord. Chem. Rev. 2021, 436, 213849. [Google Scholar] [CrossRef]
- Uivarosi, V. Metal Complexes of Quinolone Antibiotics and Their Applications: An Update. Molecules 2013, 18, 11153–11197. [Google Scholar] [CrossRef]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol. Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar] [CrossRef]
- Shen, L.L.; Chu, D.T.W. Type II DNA topoisomerases as antibacterial targets. Curr. Pharm. Des. 1996, 2, 195–208. [Google Scholar]
- Collin, F.; Karkare, S.; Maxwell, A. Exploiting bacterial DNA gyrase as a drug target: Current state and perspectives. Appl. Microbiol. Biotechnol. 2011, 92, 479–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, V.E.; Osheroff, N. Type II topoisomerases as targets for quinolone antibacterials turning Dr. Jekyll into Mr. Hyde. Curr. Pharm. Des. 2001, 7, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Talwar, P. Repositioning of fluoroquinolones from antibiotic to anti-cancer agents: An underestimated truth. Biomed. Pharmacother. 2019, 111, 934–946. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Daneshtala, M. Nonclassical biological activities of quinolone derivatives. J. Pharm. Pharmaceut. Sci. 2012, 15, 52–72. [Google Scholar]
- Hawtin, R.E.; Stockett, D.E.; Byl, J.A.W.; McDowell, R.S.; Tan, N.; Arkin, M.R.; Conroy, A.; Yang, W.; Osheroff, N.; Fox, J.A. Voreloxin Is an Anticancer Quinolone Derivative that Intercalates DNA and Poisons Topoisomerase II. PLoS ONE 2010, 5, e10186. [Google Scholar] [CrossRef]
- Advani, R.H.; Hurwitz, H.I.; Gordon, M.S.; Ebbinghaus, S.W.; Mendelson, D.S.; Wakelee, H.A.; Hoch, U.; Silverman, J.A.; Havrilla, N.A.; Berman, C.J.; et al. Voreloxin, a first-in-class anticancer quinolone derivative, in Relapsed/Refractory solid tumors: A report on two dosing schedules. Clin. Canc. Res. 2010, 16, 2167–2175. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, G.C.; Fox, J.A.; Poi, M.; Strickland, S.A. Molecular and pharmacological properties of the anticancer quinolone derivative vosaroxin: A new therapeutic agent for acute myeloid leukemia. Drugs 2016, 76, 1245–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, K.R.; Kantarjian, H.; Ravandi, F. Vosaroxin: Innovative anticancer quinolone for the treatment of acute myelogenous leukemia. Expert Opin. Orphan Drugs 2016, 4, 885–892. [Google Scholar] [CrossRef]
- Sava, G.; Zorzet, S.; Turrin, C.; Vita, F.; Soranzo, M.; Zabucchi, G.; Cocchietto, M.; Bergamo, A.; DiGiovine, S.; Pezzoni, G.; et al. Dual action of NAMI-A in inhibition of solid tumor metastasis: Selective targeting of metastatic cells and binding to collagen. Clin. Cancer Res. 2003, 9, 1898–1905. [Google Scholar]
- Jakupec, M.A.; Arion, V.B.; Kapitza, S.; Reisner, E.; Eichinger, A.; Pongratz, M.; Marian, B.; Graf von Keyserlingk, N.; Keppler, B.K. KP1019 (FFC14A) from bench to bedside: Preclinical and early clinical development—An overview. Int. J. Clin. Pharmacol. Ther. 2005, 43, 595–596. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.C.C.; Coelho, J.S.V.; Pazin, W.M.; Nikolaou, S. A newhomoleptic coordination compound of ruthenium and norfloxacin and its interaction with human serum albumin. Inorg. Chem. Commun. 2016, 63, 96–100. [Google Scholar] [CrossRef]
- Tanimoto, M.K.; Dias, K.; Dovidauskas, S.; Nikolaou, S. Tuning the reaction products of ruthenium and ciprofloxacin for studies of DNA interactions. J. Coord. Chem. 2012, 65, 1504–1517. [Google Scholar] [CrossRef]
- Alghamdi, M.T.; Alsibai, A.A.; Shahawi, M.S.; Refat, M.S. Synthesis and spectroscopic studies of levofloxacin uni-dentate complexes of Ru(II), Pt(IV) and Ir(III): Third generation of quinolone antibiotic drug complexes. J. Mol. Liq. 2016, 224, 571–579. [Google Scholar] [CrossRef]
- Alghamdi, M.T.; Alsibai, A.A.; Shahawi, M.S.; Refat, M.S. Structural and chelation behaviors of new Ru(II), Pt(IV) and Ir(III) gatifloxacin drug complexes: Spectroscopic characterizations. J. Mol. Struct. 2017, 1130, 264–275. [Google Scholar] [CrossRef]
- Alibrahim, K.A.; Al-Saif, F.A.; Alghamdi, M.T.; El-Shahawi, M.S.; Moustafa, Y.M.; Refat, M.S. Synthesis, spectroscopic, thermal, antimicrobial and electrochemical characterization of some novel Ru(III), Pt(IV) and Ir(III) complexes of pipemidic acid. RSC Adv. 2018, 8, 22515–22529. [Google Scholar] [CrossRef] [Green Version]
- Badea, M.; Olar, R.; Marinescu, D.; Uivarosi, V.; Iacob, D. Thermal decomposition of some biologically active complexes of ruthenium (III) with quinolone derivatives. J. Therm. Anal. Calorim. 2009, 97, 735–739. [Google Scholar] [CrossRef]
- Badea, M.; Olar, R.; Marinescu, D.; Uivarosi, V.; Nicolescu, T.O.; Iacob, D. Thermal study of some new quinolone ruthenium (III) complexes with potential cytostatic activity. J. Therm. Anal. Calorim. 2009, 99, 829–834. [Google Scholar] [CrossRef]
- Arsene, A.L.; Uivarosi, V.; Mitrea, N.; Dragoi, C.; Nicolae, A. In vitro studies regarding the interactions of some novel ruthenium (III) complexes with double stranded calf thymus deoxyribonucleic acid (DNA). Farmacia 2016, 64, 712–716. [Google Scholar]
- Arsene, A.L.; Uivarosi, V.; Mitrea, N.; Dragoi, C.; Nicolae, A. The binding properties of some novel ruthenium (III) complexes with human serum transferrin. Biopolym. Cell 2011, 27, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Uivarosi, V.; Olar, R.; Badea, M.; Chifiriuc, M.C. Antimicrobial activity of some new Ru(III) complexes with quinolone derivatives. Farmacia 2017, 65, 972–977. [Google Scholar]
- Gruia, M.I.; Negoita, V.; Vasilescu, M.; Panait, M.; Gruia, I.; Velescu, B.S.; Uivarosi, V. Biochemical action of some new complexes of ruthenium with quinolones as potential antitumoral agents. Anticancer Res. 2015, 35, 3371–3378. [Google Scholar]
- Munteanu, A.C.; Badea, M.; Olar, R.; Silvestro, L.; Mihaila, M.; Brasoveanu, L.I.; Musat, M.G.; Andries, A.; Uivarosi, V. Cytotoxicity studies, DNA interaction and protein binding of new Al(III), Ga(III) and In(III) complexes with 5-hydroxyflavone. Appl. Organomet. Chem. 2018, 32, e4579. [Google Scholar] [CrossRef] [Green Version]
- Maciuca, A.M.; Munteanu, A.C.; Mihaila, M.; Badea, M.; Olar, R.; Nitulescu, G.M.; Munteanu, C.V.A.; Bostan, M.; Uivarosi, V. Rare-Earth Metal Complexes of the Antibacterial Drug Oxolinic Acid: Synthesis, Characterization, DNA/Protein Binding and Cytotoxicity Studies. Molecules 2020, 25, 5418. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, A.C.; Musat, M.G.; Mihaila, M.; Badea, M.; Olar, R.; Nitulescu, G.M.; Radulescu, F.S.; Brasoveanu, L.I.; Uivarosi, V. New heteroleptic lanthanide complexes as multimodal drugs: Cytotoxicity studies, apoptosis, cell cycle analysis, DNA interactions, and protein binding. Appl. Organomet. Chem. 2020, 35, e6062. [Google Scholar] [CrossRef]
- Petrusca, D.N.; Petrescu, A.; Vrabie, C.; Jinga, V.; Diaconu, C.C.; Brasoveanu, L.I. Cell cycle regulatory factors in juxtatumoral renal parenchyma. Rom. J. Morphol. Embriol. 2005, 46, 117–122. [Google Scholar]
- Mihaila, M.; Bostan, M.; Hotnog, D.; Ferdes, M.; Brasoveanu, L.I. Real-time analysis of quercetin, resveratrol and/or doxorubicin effects in MCF-7 cells. Rom. Biotechnol. Lett. 2013, 18, 8106–8114. [Google Scholar]
- Hotnog, C.; Mihaila, M.; Panait, M.; Petrica-Matei, G.G.; Roman, V.; Gruia, M.I.; Bostan, M.; Brasoveanu, L.I. Resveratrol modulates oxidative stress as well as apoptosis in drug-treated SW1116 colon cancer cells. Rom. Arch. Microbiol. Immunol. 2017, 76, 41–49. [Google Scholar]
- Petrica-Matei, G.G.; Roman, V.; Mihaila, M.; Hotnog, C.; Brasoveanu, L.I.; Bostan, M. Role of p38-mitogen-activated protein kinase in modulation of the response to therapy in FaDu human pharyngeal carcinoma cell line. Rom. Biotechnol. Lett. 2019, 24, 118–128. [Google Scholar] [CrossRef]
- Bostan, M.; Petrica-Matei, G.G.; Ion, G.; Radu, N.; Mihaila, M.; Hainarosie, R.; Brasoveanu, L.I.; Roman, V.; Constantin, C.; Neagu, M.T. Cisplatin effect on head and neck squamous cell carcinoma cells is modulated by ERK1/2 protein kinases. Exp. Ther. Med. 2019, 18, 5041–5051. [Google Scholar] [CrossRef] [Green Version]
- Lacatusu, I.; Badea, N.; Badea, G.; Oprea, O.; Mihaila, M.A.; Kaya, D.A.; Stan, R.; Meghea, A. Lipid nanocarriers based on natural oils with high activity against oxygen free radicals and tumor cell proliferation. Mater. Sci. Eng. C 2015, 56, 88–94. [Google Scholar] [CrossRef]
- Lacatusu, I.; Badea, N.; Badea, G.; Brasoveanu, L.; Stan, R.; Ott, C.; Oprea, O.; Meghea, A. Ivy leaves extract based—Lipid nanocarriers and their bioefficacy on antioxidant and antitumor activities. RSC Adv. 2016, 6, 77243–77255. [Google Scholar] [CrossRef]
- Knop, C.; Putnik, J.; Scheuermann, M.; Schmitz, M. Cutting Edge Technologies: Cell Analysis; Ziebolz, B., Ed.; Springer Medizin: Heidelberg, Germany, 2010; pp. 4–13, 58–68, 137–141. [Google Scholar]
- Teixeira, R.G.; Belisario, D.C.; Fontrodona, X.; Romero, I.; Tomaz, A.I.; Garcia, M.H.; Rigantib, C.; Valente, A. Unprecedented collateral sensitivity for cisplatin resistant lung cancer cells presented by new ruthenium organometallic compounds. Inorg. Chem. Front. 2021, 8, 1983–1996. [Google Scholar] [CrossRef]
- Zimmermann, K.C.; Bonzon, C.; Green, D.R. The machinery of programmed cell death. Pharmacol. Ther. 2001, 92, 57–70. [Google Scholar] [CrossRef]
- Townson, J.L.; Naumov, G.N.; Chambers, A.F. The role of apoptosis in tumor progression and metastasis. Curr. Mol. Med. 2003, 3, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Desoize, B.; Jardillier, J. Multicellular resistance: A paradigm for clinical resistance? Crit. Rev. Oncol. Hematol. 2000, 36, 193–207. [Google Scholar] [CrossRef]
- Gillison, T.L.; Gurkamal, S.C. Cancer chemotherapy in the elderly patient. Oncology 2010, 24, 76–85. [Google Scholar] [PubMed]
- Zhang, L.; Yu, J. Role of apoptosis in colon cancer biology, therapy, and prevention. Curr. Colorect. Cancer Rep. 2013, 9, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Bostan, M.; Mihaila, M.; Hotnog, C.; Bleotu, C.; Anton, G.; Roman, V.; Brasoveanu, L.I. Chapter Modulation of Apoptosis in Colon Cancer Cells by Bioactive Compounds. In Colorectal Cancer—From Pathogenesis to Treatment; Rodrigo, L., Ed.; Intech: Rijeka, Croatia, 2016. [Google Scholar]
- Ucker, D.S.; Levine, J.S. Exploitation of apoptotic regulation in cancer. Front. Immunol. 2018, 9, 241. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Castillo Ferrer, C.; Berthenet, K.; Ichim, G. Apoptosis—Fueling the oncogenic fire. FEBS J. 2021, 288, 4445–4463. [Google Scholar] [CrossRef]
- Kaufmann, S.H.; Vaux, D.L. Alterations in the apoptotic machinery and their potential role in anticancer drug resistance. Oncogene 2003, 22, 7414–7430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volm, M.; Efferth, T. Prediction of cancer drug resistance and implications for personalized medicine. Front. Oncol. 2015, 5, 282. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).