
The Influence of Nutrient Medium Composition on Escherichia coli Biofilm Development and Heterologous Protein Expression


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
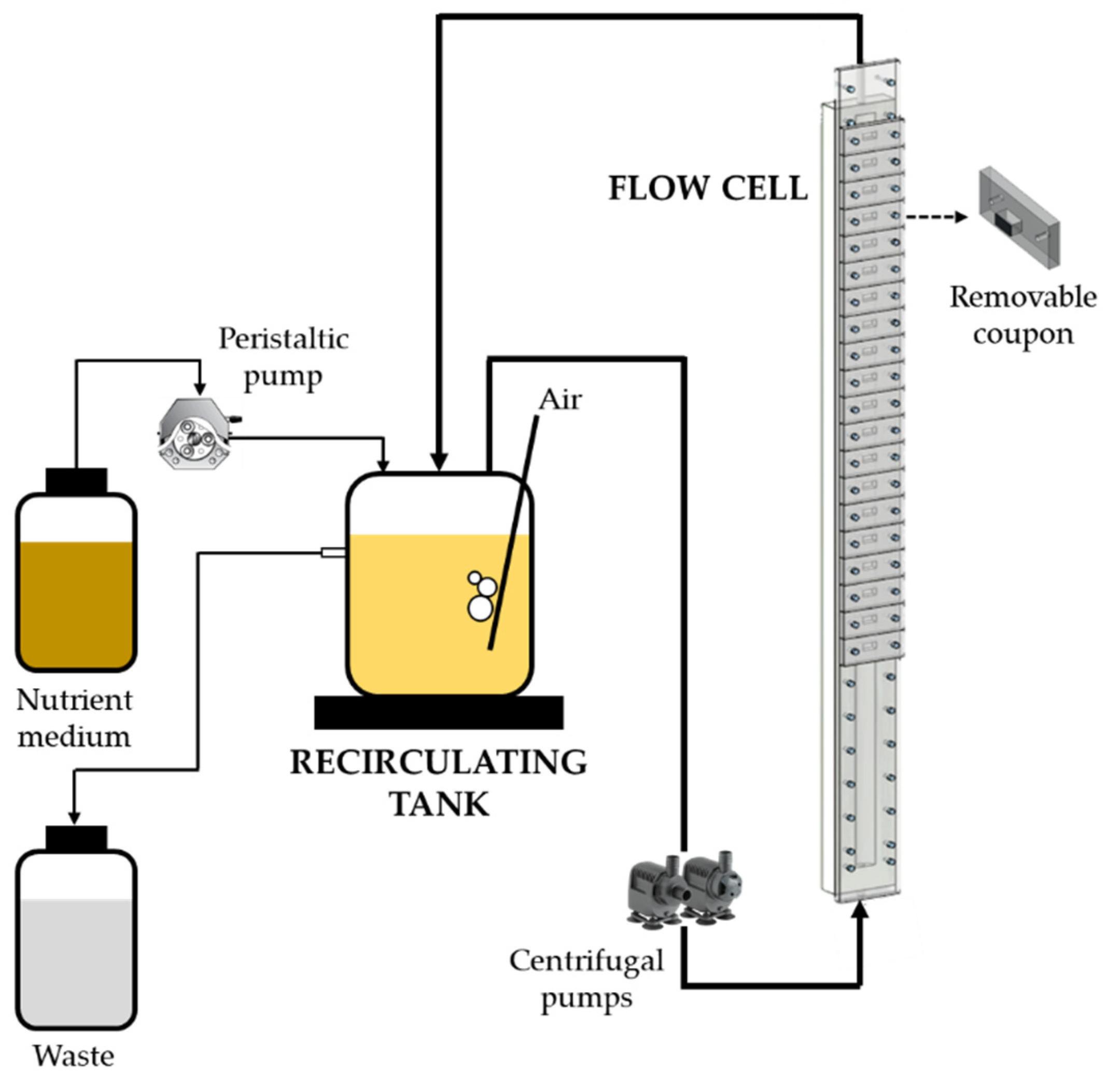
2.2. Flow System
2.3. Biofilm and Planktonic Analysis
2.3.1. Quantification of eGFP-Expressing Cells and Specific Protein Concentration
2.3.2. Plasmid Extraction and Quantification
2.4. Statistical Analysis
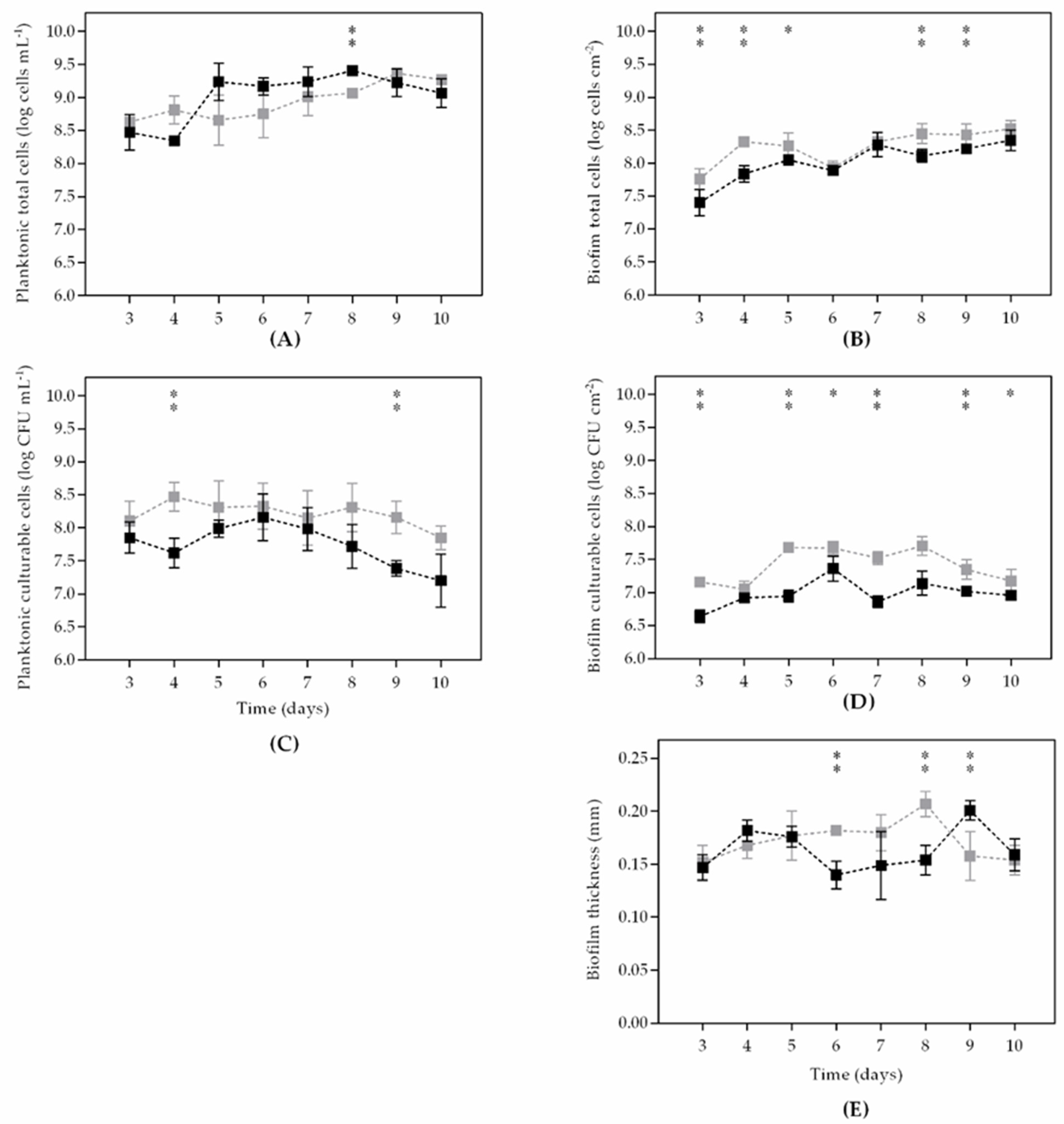
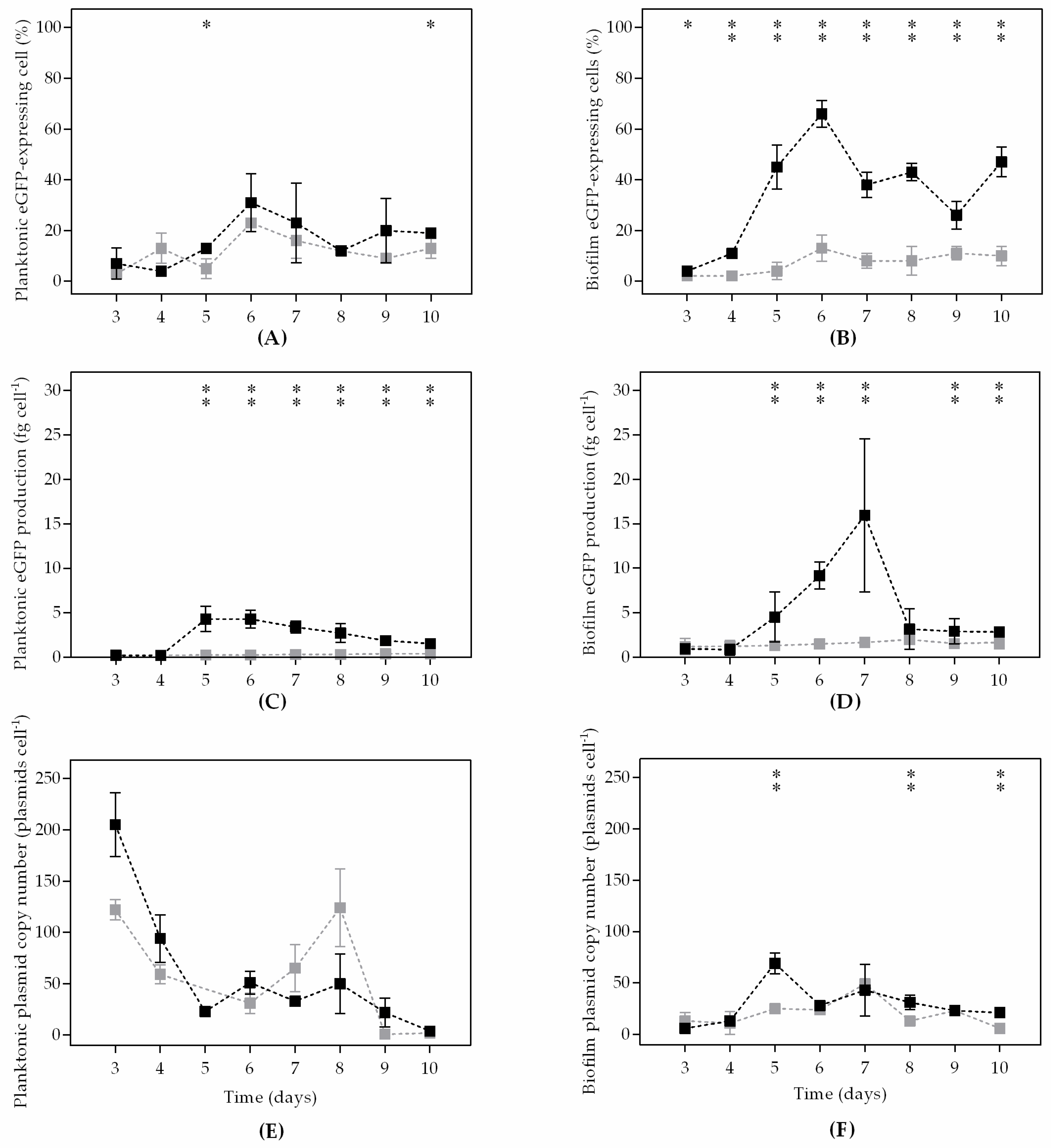
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demain, A.L.; Vaishnav, P. Production of recombinant proteins by microbes and higher organisms. Biotechnol. Adv. 2009, 27, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Gopal, G.J.; Kumar, A. Strategies for the production of recombinant protein in Escherichia coli. Protein J. 2013, 32, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Azevedo, A.; Gomes, L.C.; Mergulhão, F.J. Recombinant protein expression in biofilms. AIMS Microbiol. 2019, 5, 232. [Google Scholar] [CrossRef]
- Overton, T.W. Recombinant protein production in bacterial hosts. Drug Discov. Today 2014, 19, 590–601. [Google Scholar] [CrossRef]
- Mergulhão, F.J.; Monteiro, G.A.; Cabral, J.M.S.; Taipas, M.A. Design of bacterial vector systems for the production of recombinant proteins in Escherichia coli. J. Microbiol. Biotechnol. 2004, 14, 1–14. [Google Scholar]
- Gomes, L.C.; Monteiro, G.A.; Mergulhão, F.J. The Impact of IPTG Induction on Plasmid Stability and Heterologous Protein Expression by Escherichia coli Biofilms. Int. J. Mol. Sci. 2020, 21, 576. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.; Valdes, J.; Bentley, W. A comparative study of global stress gene regulation in response to overexpression of recombinant proteins in Escherichia coli. Metab. Eng. 2000, 2, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, W.; Wu, J.; Zhang, Y.; Zhu, Z.; Liu, J.; Hu, Z. Stability of plasmid and expression of a recombinant gonadotropin-releasing hormone (GnRH) vaccine in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 73, 780–788. [Google Scholar] [CrossRef]
- Gomes, L.C.; Carvalho, D.; Briandet, R.; Mergulhão, F.J. Temporal variation of recombinant protein expression in Escherichia coli biofilms analysed at single-cell level. Process Biochem. 2016, 51, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.C.; Mergulhão, F.J. Heterologous protein production in Escherichia coli biofilms: A non-conventional form of high cell density cultivation. Process Biochem. 2017, 57, 1–8. [Google Scholar] [CrossRef]
- Gomes, L.C.; Mergulhão, F.J. Effects of antibiotic concentration and nutrient medium composition on Escherichia coli biofilm formation and green fluorescent protein expression. FEMS Microbiol. Lett. 2017, 364, fnx042. [Google Scholar] [CrossRef] [Green Version]
- Wijeyekoon, S.; Mino, T.; Satoh, H.; Matsuo, T. Effects of substrate loading rate on biofilm structure. Water Res. 2004, 38, 2479–2488. [Google Scholar] [CrossRef]
- Bühler, T.; Ballestero, S.; Desai, M.; Brown, M.R.W. Generation of a reproducible nutrient-depleted biofilm of Escherichia coli and Burkholderia cepacia. J. Appl. Microbiol. 1998, 85, 457–462. [Google Scholar] [CrossRef]
- Domka, J.; Lee, J.; Bansal, T.; Wood, T.K. Temporal gene-expression in Escherichia coli K-12 biofilms. Environ. Microbiol. 2007, 9, 332–346. [Google Scholar] [CrossRef]
- Oh, Y.; Jo, W.; Yang, Y.; Park, S. Influence of culture conditions on Escherichia coli O157: H7 biofilm formation by atomic force microscopy. Ultramicroscopy 2007, 107, 869–874. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, H.A.; Niu, C.; Gilbert, E.S. Enhanced high copy number plasmid maintenance and heterologous protein production in an Escherichia coli biofilm. Biotechnol. Bioeng. 2007, 97, 439–446. [Google Scholar] [CrossRef]
- Rosano, G.L.; Morales, E.S.; Ceccarelli, E.A. New tools for recombinant protein production in Escherichia coli: A 5-year update. Protein Sci. 2019, 28, 1412–1422. [Google Scholar] [CrossRef]
- Terol, G.L.; Gallego-Jara, J.; Martínez, R.A.S.; Vivancos, A.M.; Díaz, M.C.; de Diego Puente, T. Impact of the expression system on recombinant protein production in Escherichia coli BL21. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Waegeman, H.; Soetaert, W. Increasing recombinant protein production in Escherichia coli through metabolic and genetic engineering. J. Ind. Microbiol. Biotechnol. 2011, 38, 1891–1910. [Google Scholar] [CrossRef] [PubMed]
- Bjergbæk, L.A.; Haagensen, J.A.J.; Reisner, A.; Molin, S.; Roslev, P. Effect of oxygen and growth medium on in vitro biofilm formation by Escherichia coli. Biofilms 2006, 3, 1. [Google Scholar] [CrossRef]
- Bowden, G.; Li, Y. Nutritional influences on biofilm development. J. Adv. Dent. Res. 1997, 11, 81–99. [Google Scholar] [CrossRef]
- Teodósio, J.S.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Flow cell hydrodynamics and their effects on E. coli biofilm formation under different nutrient conditions and turbulent flow. Biofouling 2011, 27, 1–11. [Google Scholar] [CrossRef]
- Bao, R.M.; Yang, H.M.; Yu, C.M.; Zhang, W.F.; Tang, J.B. An efficient protocol to enhance the extracellular production of recombinant protein from Escherichia coli by the synergistic effects of sucrose, glycine, and Triton X-100. Protein Expres. Purif. 2016, 126, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Khurana, J.; Pratibha, C.; Kaur, J. Studies on recombinant lipase production by E. coli: Effect of media and bacterial expression system optimization. Mol. Biol. 2017, 2, 00008. [Google Scholar]
- Ukkonen, K.; Veijola, J.; Vasala, A.; Neubauer, P. Effect of culture medium, host strain and oxygen transfer on recombinant Fab antibody fragment yield and leakage to medium in shaken E. coli cultures. Microb. Cell Factories 2013, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Goyal, D.; Sahni, G.; Sahoo, D.K. Enhanced production of recombinant streptokinase in Escherichia coli using fed-batch culture. Bioresour. Technol. 2009, 100, 4468–4474. [Google Scholar] [CrossRef] [PubMed]
- Kweon, D.H.; Han, N.S.; Park, K.M.; Seo, J.H. Overproduction of Phytolacca insularis protein in batch and fed-batch culture of recombinant Escherichia coli. Process. Biochem. 2001, 36, 537–542. [Google Scholar] [CrossRef]
- Matsui, T.; Sato, H.; Sato, S.; Mukataka, S.; Takahashi, J. Effects of nutritional conditions on plasmid stability and production of tryptophan synthase by a recombinant Escherichia coli. Agr. Biol. Chem. 1990, 54, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, F.; Rinas, U. Stress induced by recombinant protein production in Escherichia coli. In Physiological Stress Responses in Bioprocesses; Springer: Berlin/Heidelberg, Germany, 2004; pp. 73–92. [Google Scholar] [CrossRef]
- Novagen. pET System Manual, 11th ed.; EMD Biosciences: Darmstadt, Germany, 2005. [Google Scholar]
- Hahm, D.H.; Kim, S.H.; Pan, J.; Rhee, J.S. Maximum yield of foreign lipase in Escherichia coli HB101 limited by duration of protein expression. J. Ferment. Bioeng. 1995, 79, 236–241. [Google Scholar] [CrossRef]
- Mergulhão, F.J.; Monteiro, G.A. Analysis of factors affecting the periplasmic production of recombinant proteins in Escherichia coli. J. Microbiol. Biotechnol. 2007, 17, 1236. [Google Scholar]
- Teodósio, J.S.; Simões, M.; Mergulhão, F.J. The influence of nonconjugative Escherichia coli plasmids on biofilm formation and resistance. J. Appl. Microbiol. 2012, 113, 373–382. [Google Scholar] [CrossRef]
- Teodósio, J.S.; Silva, F.C.; Moreira, J.M.; Simões, M.; Melo, L.F.; Alves, M.A.; Mergulhão, F.J. Flow cells as quasi-ideal systems for biofouling simulation of industrial piping systems. Biofouling 2013, 29, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system. J. Mol. Biol. 1991, 219, 37–44. [Google Scholar] [CrossRef]
- Alves, C.P.; Šimčíková, M.; Brito, L.; Monteiro, G.A.; Prazeres, D.M.F. Production and purification of supercoiled minicircles by a combination of in vitro endonuclease nicking and hydrophobic interaction chromatography. Hum. Gene Ther. Methods 2018, 29, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Diogo, M.; Queiroz, J.; Prazeres, D.M.F. Assessment of purity and quantification of plasmid DNA in process solutions using high-performance hydrophobic interaction chromatography. J. Chromatogr. A 2003, 998, 109–117. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Soares, A.; Gomes, L.C.; Mergulhão, F.J. Comparing the recombinant protein production potential of planktonic and biofilm cells. Microorganisms 2018, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.C.; Silva, L.N.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Escherichia coli adhesion, biofilm development and antibiotic susceptibility on biomedical materials. J. Biomed. Mater. Res. A 2015, 103, 1414–1423. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, N.K.; Shrivastva, A.; Biswal, K.C.; Rao, P.V.L. METHODS: Optimization of culture medium for production of recombinant dengue protein in Escherichia coli. Ind. Biotechnol. 2009, 5, 179–183. [Google Scholar] [CrossRef]
- Prüß, B.M.; Verma, K.; Samanta, P.; Sule, P.; Kumar, S.; Wu, J.; Christianson, D.; Horne, S.M.; Stafslien, S.J.; Wolfe, A.J. Environmental and genetic factors that contribute to Escherichia coli K-12 biofilm formation. Arch. Microbiol. 2010, 192, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Seneviratne, C.J.; Yip, J.W.Y.; Chang, J.W.W.; Zhang, C.F.; Samaranayake, L.P. Effect of culture media and nutrients on biofilm growth kinetics of laboratory and clinical strains of Enterococcus faecalis. Arch. Oral Biol. 2013, 58, 1327–1334. [Google Scholar] [CrossRef]
- Dewanti, R.; Wong, A.C. Influence of culture conditions on biofilm formation by Escherichia coli O157: H7. Int. J. Food Microbiol. 1995, 26, 147–164. [Google Scholar] [CrossRef]
- Yoon, S.K.; Kang, W.K.; Park, T.H. Fed-batch operation of recombinant Escherichia coli containing trp promoter with controlled specific growth rate. Biotechnol. Bioeng. 1994, 43, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Grossman, T.H.; Kawasaki, E.S.; Punreddy, S.R.; Osburne, M.S. Spontaneous cAMP-dependent derepression of gene expression in stationary phase plays a role in recombinant expression instability. Gene 1998, 209, 95–103. [Google Scholar] [CrossRef]
- Jevševar, S.; Palčič, J.; Jalen, Š.; Pavko, A. Influence of the Media Composition on Behavior of pET Expression Systems. Acta Chim. Slov. 2007, 54, 360–365. [Google Scholar]
- Li, X.; Robbins, J.; Taylor, K. The production of recombinant beta-galactosidase in Escherichia coli in yeast extract enriched medium. J. Ind. Microbiol. Biotechnol. 1990, 5, 85–93. [Google Scholar] [CrossRef]
- Solaiman, D.K.; Somkuti, G.A. Expression of streptomycete cholesterol oxidase in Escherichia coli. J. Ind. Microbiol. 1991, 8, 253–258. [Google Scholar] [CrossRef]
- Singha, T.K.; Gulati, P.; Mohanty, A.; Khasa, Y.P.; Kapoor, R.K.; Kumar, S. Efficient genetic approaches for improvement of plasmid based expression of recombinant protein in Escherichia coli: A review. Process Biochem. 2017, 55, 17–31. [Google Scholar] [CrossRef]
- Williams, I.; Venables, W.A.; Lloyd, D.; Paul, F.; Critchley, I. The effects of adherence to silicone surfaces on antibiotic susceptibility in Staphylococcus aureus. Microbiology 1997, 143, 2407–2413. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, A.; Gomes, L.C.; Monteiro, G.A.; Mergulhão, F.J. The Influence of Nutrient Medium Composition on Escherichia coli Biofilm Development and Heterologous Protein Expression. Appl. Sci. 2021, 11, 8667. https://doi.org/10.3390/app11188667
Soares A, Gomes LC, Monteiro GA, Mergulhão FJ. The Influence of Nutrient Medium Composition on Escherichia coli Biofilm Development and Heterologous Protein Expression. Applied Sciences. 2021; 11(18):8667. https://doi.org/10.3390/app11188667
Chicago/Turabian StyleSoares, Alexandra, Luciana C. Gomes, Gabriel A. Monteiro, and Filipe J. Mergulhão. 2021. "The Influence of Nutrient Medium Composition on Escherichia coli Biofilm Development and Heterologous Protein Expression" Applied Sciences 11, no. 18: 8667. https://doi.org/10.3390/app11188667