
Tyrosol, at the Concentration Found in Maltese Extra Virgin Olive Oil, Induces HL-60 Differentiation towards the Monocyte lineage

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Phenolic Extraction and Identification
2.4. Induction of Differentiation
2.5. Evaluation of Differentiation
2.6. Assessment of the Specificity of the Anti-Proliferative Action
2.7. Leishman’s Stain for Visualization of Morphological Changes
2.8. Evaluation of the Expression of Differentiation Specific Cell Markers by Flow Cytometry
2.9. Cell Cycle Analysis by Flow Cytometry
2.10. Investigation of the Molecular Mechanism of Differentiation
2.11. RNA-Seq Data Analysis
2.12. Statistical Analysis
2.13. Summary of the Study
3. Results
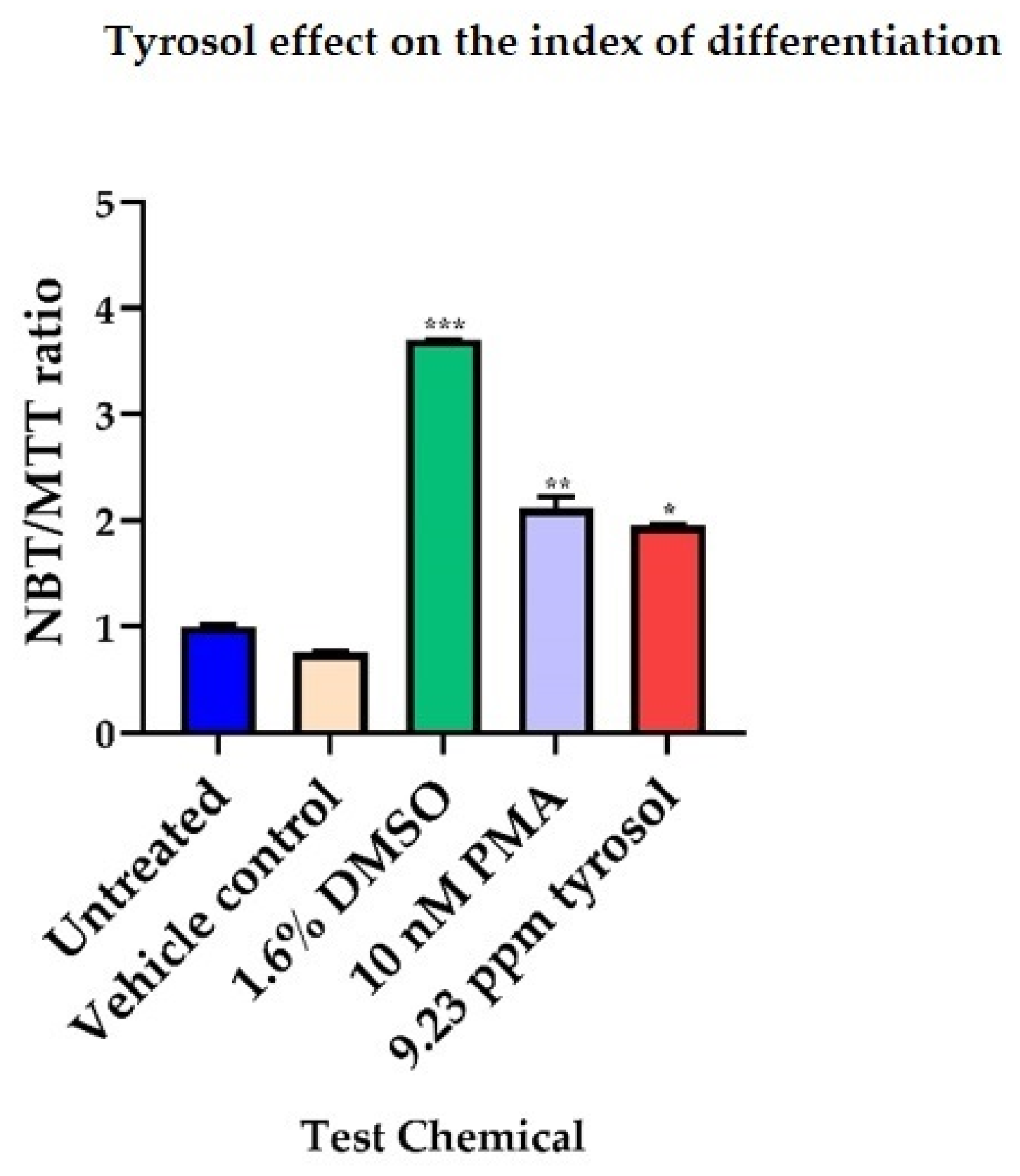
3.1. Tyrosol Shows Differentiating Activity in HL-60 Cells
3.2. Morphological Analysis of Tyrosol-Exposed HL-60 Cells Show Evidence of Monocytic Differentiation
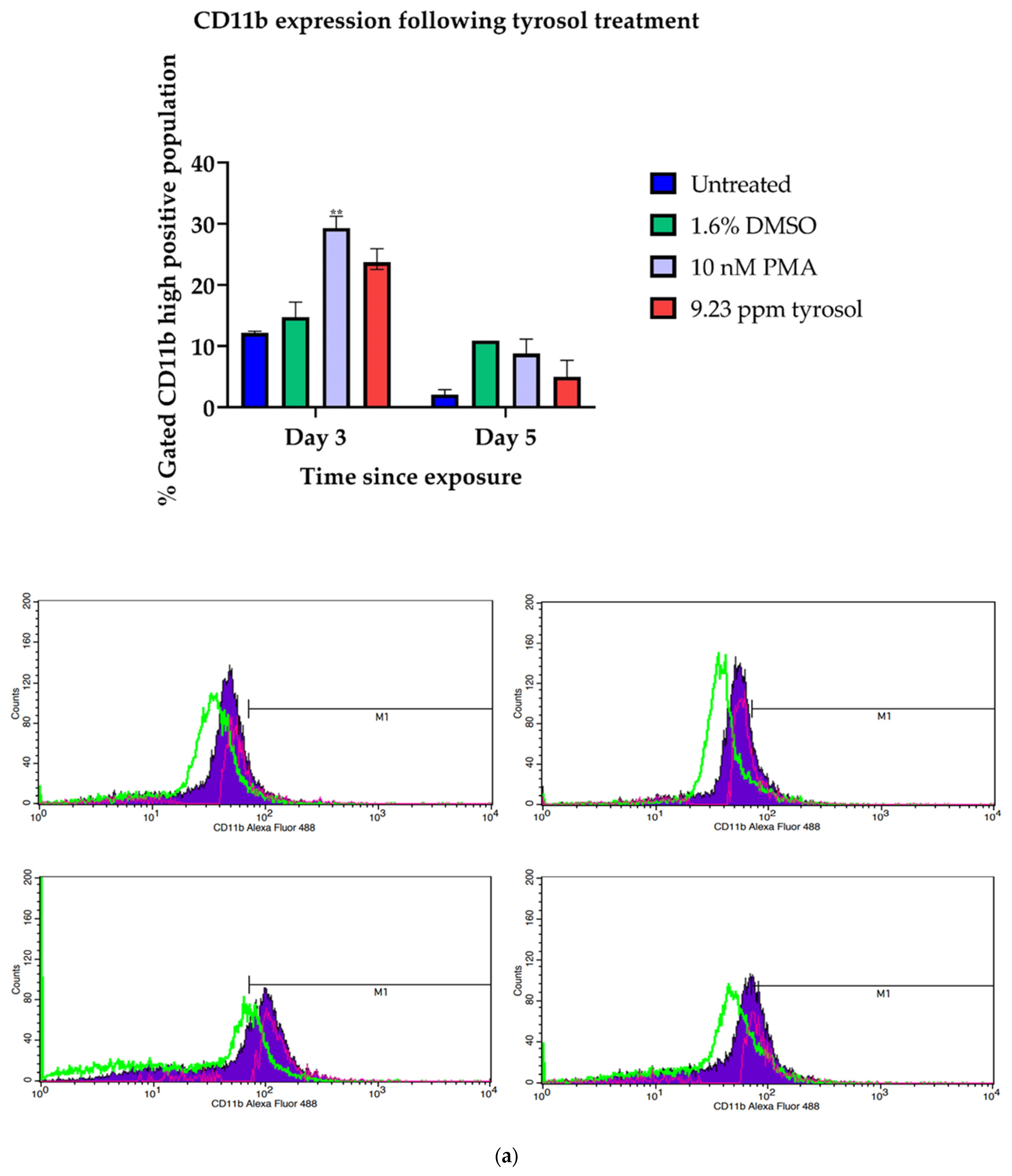
3.3. Tyrosol Stimulates the Expression of CD11b and CD14 Surface Antigens in Hl-60 Cells
3.4. Tyrosol Induces Apoptosis in Hl-60 Associated with a Reduction in Cells in the G1 and S Phases
3.5. Tyrosol Downregulates Neutrophil and Cholesterol Biosynthesis Genes and Upregulates Monocytic Differentiation Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Virchow, R. Weisses blut. Froriep’s Notizen. 1847, 36, 151–156. [Google Scholar]
- Estey, E.H. Acute myeloid leukemia: 2012 update on diagnosis, risk stratification, and management. Am. J. Hematol. 2012, 87, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Pierce, G.B.; Shikes, R.; Fink, L.M. Cancer: A Problem of Developmental Biology; Prentice-Hall Inc: Englewood Cliffs, NJ, USA, 1978; p. 242. [Google Scholar]
- Nowak, D.; Stewart, D.; Koeffler, H.P. Differentiation therapy of leukemia: 3 decades of development. Blood 2009, 113, 3655–3665. [Google Scholar] [CrossRef]
- Tallman, M.S.; Lefèbvre, P.; Baine, R.M.; Shoji, M.; Cohen, I.; Green, D.; Kwaan, H.C.; Paietta, E.; Rickles, F.R. Effects of all-trans retinoic acid or chemotherapy on the molecular regulation of systemic blood coagulation and fibrinolysis in patients with acute promyelocytic leukemia. J. Thromb. Haemost. 2004, 2, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Sell, S. Leukemia: Stem cells, maturation arrest and differentiation therapy. Stem Cell Rev. 2005, 1, 197–205. [Google Scholar] [CrossRef]
- Leszczyniecka, M.; Roberts, T.; Dent, P.; Grant, S.; Fisher, P.B. Differentiation therapy of human cancer: Basic science and clinical applications. Pharmacol. Ther. 2001, 90, 105–156. [Google Scholar] [CrossRef]
- Birnie, G.D. The HL60 cell line: A model system for studying human myeloid cell differentiation. Br. J. Cancer. Supplement. 1988, 9, 41–45. [Google Scholar]
- Salvi, H.Y.; Aalto, B.; Nagy, S.; Knuutila, S.; Pakkala, S. Gene expression analysis of 1,25(OH)2D3-dependent differentiation of HL-60 cells: A cDNA array study. Br. J. Hematol. 2002, 118, 1065–1070. [Google Scholar]
- Chang, H.H.; Oh, P.Y.; Ingber, D.E.; Huang, S. Multistable and multistep dynamics in neutrophil differentiation. BMC Cell Biol. 2006, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newburger, P.E.; Subrahmanyam, Y.V.; Weissman, S.M. Global analysis of neutrophil gene expression. Curr. Opin. Hematol. 2000, 7, 16–20. [Google Scholar] [CrossRef]
- Rincón, E.; Rocha-Gregg, B.L.; Collins, S.R. A map of gene expression in neutrophil-like cell lines. BMC Genom. 2018, 19, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naegelen, I.; Plançon, S.; Nicot, N.; Kaoma, T.; Muller, A.; Vallar, L.; Tschirhart, E.J.; Bréchard, S. An essential role of syntaxin 3 protein for granule exocytosis and secretion of IL-1α, IL-1β, IL-12b, and CCL4 from differentiated HL-60 cells. J. Leukoc. Biol. 2014, 97, 557–571. [Google Scholar] [CrossRef]
- Mark Welch, D.B.; Jauch, A.; Langowski, J.; Olins, A.L.; Olins, D.E. Transcriptomes reflect the phenotypes of undifferentiated, granulocyte and macrophage forms of HL-60/S4 cells. Nucleus 2017, 8, 222–237. [Google Scholar] [CrossRef] [Green Version]
- Santos-Beneit, A.M.; Mollinedo, F. Expression of genes involved in initiation, regulation, and execution of apoptosis in human neutrophils and during neutrophil differentiation of HL-60 cells. J. Leukoc Biol. 2000, 67, 712–724. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Gómez-Romero, M.; García-Villalba, R.; Carrasco-Pancorbo, A.; Fernández-Gutiérrez, A. Metabolism and Bioavailability of Olive Oil Polyphenols (2012). In Olive Oil—Constituents, Quality, Health Properties and Bioconversions; Dimitrios, B., Ed.; IntechOpen: London, UK; pp. 333–356.
- Shendi, E.G.; Ozay, D.S.; Ozkaya, M.T.; Ustunei, N.F. Changes occurring in chemical composition and oxidative stability of virgin olive oil during storage. Oilseeds Fats Crop. Lipids 2018, 25, 1–8. [Google Scholar]
- Lia, F.; Formosa, J.P.; Zammit-Mangion, M.; Farrugia, C. The First Identification of the Uniqueness and Authentication of Maltese Extra Virgin Olive Oil Using 3D-Fluorescence Spectroscopy Coupled with Multi-Way Data Analysis. Foods 2020, 9, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatt, L.; Lia, F.; Zammit-Mangion, M.; Thorpe, S.J.; Schembri-Wismayer, P. First Profile of Phenolic Compounds from Maltese Extra Virgin Olive Oils Using Liquid-Liquid Extraction and Liquid Chromatography-Mass Spectrometry. J. Oleo Sci. 2021, 70, 145–153. [Google Scholar] [CrossRef]
- Fabiani, R.; De Bartolomeo, A.; Rosignoli, P.; Servili, M.; Selvaggini, R.; Montedoro, G.F.; Di Saverio, C.; Morozzi, G. Virgin olive oil phenols inhibit proliferation of human promyelocytic leukemia cells (HL60) by inducing apoptosis and differentiation. J. Nutr. 2006, 136, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Abaza, L.; Talorete, T.P.N.; Yamada, P.; Kurita, Y.; Zarrouk, M.; Isoda, H. Induction of growth inhibition and differentiation of human leukemia HL-60 cells by a Tunisian gerboui olive leaf extract. Biosci. Biotechnol. Biochem. 2007, 71, 1306–1312. [Google Scholar] [CrossRef]
- Sepporta, M.V.; Mazza, T.; Morozzi, G.; Fabiani, R. Pinoresinol inhibits proliferation and induces differentiation on human HL60 leukemia cells. Nutr. Cancer 2013, 65, 1208–1218. [Google Scholar] [CrossRef]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Morozzi, G. Inhibition of cell cycle progression by hydroxytyrosol is associated with upregulation of cyclin-dependent protein kinase inhibitors p21(WAF1/Cip1) and p27(Kip1) and with induction of differentiation in HL60 cells. J. Nutr. 2008, 138, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, K.; Matsuki, N. Measurement of cellular 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction activity and lactate dehydrogenase release using MTT. Neurosci. Res. 2000, 38, 325–329. [Google Scholar] [CrossRef]
- Stoica, S.; Magoulas, G.E.; Antoniou, A.I.; Suleiman, S.; Cassar, A.; Gatt, L.; Papaioannou, D.; Athanassopoulos, C.M.; Schembri-Wismayer, P. Synthesis of minoxidil conjugates and their evaluation as HL-60 differentiation agents. Bioorganic Med. Chem. Lett. 2016, 26, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Heiniger, H.J.; Kandutsch, A.A. Relationship between sterol synthesis and DNA synthesis in phytohemagglutinin-stimulated mouse lymphocytes. Proc. Natl. Acad. Sci. USA 1975, 72, 1950–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mire-Sluis, A.R.; Wickremasinghe, R.G.; Hoffbrand, A.V.; Timms, A.M.; Francis, G.E. Human T lymphocytes stimulated by phytohaemagglutinin undergo a single round of cell division without a requirement for interleukin-2 or accessory cells. Immunology 1987, 60, 7–12. [Google Scholar]
- Hutchins, D.; Steel, C.M. Phytohaemagglutinin-induced proliferation of human T lymphocytes: Differences between neonate and adults in accessory cell requirements. Clin. Exp. Immunol. 1983, 52, 355–364. [Google Scholar]
- Enright, A.J.; Van Dongen, S.; Ouzounis, C.A.A. An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 2002, 30, 1575–1584. [Google Scholar] [CrossRef]
- Kawaii, S.; Lansky, E.P. Differentiation-promoting activity of pomegranate (Punica granatum) fruit extracts in HL-60 human promyelocytic leukemia cells. J. Med. Food 2004, 7, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Crescimanno, M.; Sepporta, M.V.; Tripoli, E.; Flandina, C.; Giammanco, M.; Tumminello, F.T.; Di Majo, D.; Tolomeo, M.; La Guardia, M.; Leto, G. Effects of extra virgin olive oil phenols on HL60 cell lines sensitive and resistant to anthracyclines. J. Biol. Res. 2009, 82, 34–37. [Google Scholar] [CrossRef]
- Baillie, J.K.; Arner, E.; Daub, C.; De Hoon, M.; Itoh, M.; Kawaji, H.; Lassmann, T.; Carninci, P.; Forrest, A.R.; Hayashizaki, Y.; et al. Analysis of the human monocyte-derived macrophage transcriptome and response to lipopolysaccharide provides new insights into genetic aetiology of inflammatory bowel disease. PLoS Genet. 2017, 13, e1006641. [Google Scholar] [CrossRef] [PubMed]
- Saliba, D.G.; Heger, A.; Eames, H.L.; Oikonomopoulos, S.; Teixeira, A.; Blazek, K.; Androulidaki, A.; Wong, D.; Goh, F.G.; Weiss, M.; et al. IRF5:RelA interaction targets inflammatory genes in macrophages. Cell Rep. 2014, 8, 1308–1317. [Google Scholar] [CrossRef] [Green Version]
- Birmachu, W.; Gleason, R.M.; Bulbulian, B.J.; Riter, C.L.; Vasilakos, J.P.; Lipson, K.E.; Nikolsky, Y. Transcriptional networks in plasmacytoid dendritic cells stimulated with synthetic TLR 7 agonists. BMC Immunol. 2007, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Ragione, F.D.; Cucciolla, V.; Borriello, A.; Pietra, V.D.; Pontoni, G.; Racioppi, L.; Manna, C.; Galletti, P.; Zappia, V. Hydroxytyrosol, a natural molecule occurring in olive oil, induces cytochrome c-dependent apoptosis. Biochem. Biophys. Res. Commun. 2000, 278, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Merck, E.; Gaillard, C.; Gorman, D.M.; Montero-Julian, F.; Durand, I.; Zurawski, S.M.; Menetrier-Caux, C.; Carra, G.; Lebecque, S.; Trinchieri, G.; et al. OSCAR is an FcRgamma-associated receptor that is expressed by myeloid cells and is involved in antigen presentation and activation of human dendritic cells. Blood 2004, 104, 1386–1395. [Google Scholar] [CrossRef] [Green Version]
- Goettsch, C.; Kliemt, S.; Sinningen, K.; von Bergen, M.; Hofbauer, L.C.; Kalkhof, S. Quantitative proteomics reveals novel functions of osteoclast-associated receptor in STAT signaling and cell adhesion in human endothelial cells. J. Mol. Cell. Cardiol. 2012, 53, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Rajappa, P.; Hu, W.; Hoffman, C.; Cisse, B.; Kim, J.H.; Gorge, E.; Yanowitch, R.; Cope, W.; Vartanian, E.; et al. A proangiogenic signaling axis in myeloid cells promotes malignant progression of glioma. J. Clin. Investig. 2017, 127, 1826–1838. [Google Scholar] [CrossRef] [Green Version]
- Czepluch, F.S.; Olieslagers, S.; van Hulten, R.; Vöö, S.A.; Waltenberger, J. VEGF-A-induced chemotaxis of CD16+ monocytes is decreased secondary to lower VEGFR-1 expression. Atherosclerosis 2011, 215, 331–338. [Google Scholar] [CrossRef]
- Fontana, M.F.; Baccarella, A.; Pancholi, N.; Pufall, M.A.; Herbert, D.R.; Kim, C.C. JUNB is a key transcriptional modulator of macrophage activation. J. Immunol. 2015, 194, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Chute, J.P.; Muramoto, G.G.; Whitesides, J.; Colvin, M.; Safi, R.; Chao, N.J.; McDonnell, D.P. Inhibition of aldehyde dehydrogenase and retinoid signaling induces the expansion of human hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 11707–11712. [Google Scholar] [CrossRef] [Green Version]
- Matikainen, S.; Ronni, T.; Lehtonen, A.; Sareneva, T.; Melén, K.; Nordling, S.; Levy, D.E.; Julkunen, I. Retinoic acid induces signal transducer and activator of transcription (STAT) 1, STAT2, and p48 expression in myeloid leukemia cells and enhances their responsiveness to interferons. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1997, 8, 687–698. [Google Scholar]
- Cohen, S.; Dovrat, S.; Sarid, R.; Huberman, E.; Salzberg, S. JAK-STAT signaling involved in phorbol 12-myristate 13-acetate- and dimethyl sulfoxide-induced 2’-5’ oligoadenylate synthetase expression in human HL-60 leukemia cells. Leuk. Res. 2005, 29, 923–931. [Google Scholar] [CrossRef]
- Matikainen, S.; Ronni, T.; Hurme, M.; Pine, R.; Julkunen, I. Retinoic acid activates interferon regulatory factor-1 gene expression in myeloid cells. Blood 1996, 88, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Gianni, M.; Terao, M.; Fortino, I.; LiCalzi, M.; Viggiano, V.; Barbui, T.; Rambaldi, A.; Garattini, E. Stat1 is induced and activated by all-trans retinoic acid in acute promyelocytic leukemia cells. Blood 1997, 89, 1001–1012. [Google Scholar] [CrossRef]
- Harada, H.; Kitagawa, M.; Tanaka, N.; Yamamoto, H.; Harada, K.; Ishihara, M.; Taniguchi, T. Anti-oncogenic and oncogenic potentials of interferon regulatory factors-1 and -2. Science 1993, 259, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.E.; Gray, D.A.; Berney, J.J.; Wing, M.A.; Guimaraes, J.E.; Hoffbrand, A.V. Role of ADP-ribosyl transferase in differentiation of human granulocyte-macrophage progenitors to the macrophage lineage. Blood 1983, 62, 1055–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, M.; Kirkland, J.B.; Meckling-Gill, K.A. Modulation of poly (ADP-ribose) polymerase during neutrophilic and monocytic differentiation of promyelocytic (NB4) and myelocytic (HL-60) leukaemia cells. Biochem. J. 1995, 308, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, H.; Goettsch, C.; Sharma, A.; Ricchiuto, P.; Goh, W.W.; Halu, A.; Yamada, I.; Yoshida, H.; Hara, T.; Wei, M.; et al. PARP9 and PARP14 cross-regulate macrophage activation via STAT1 ADP-ribosylation. Nat. Commun. 2016, 7, 12849. [Google Scholar] [CrossRef] [PubMed]
- Tieri, P.; Termanini, A.; Bellavista, E.; Salvioli, S.; Capri, M.; Franceschi, C. Charting the NF-κB pathway interactome map. PLoS ONE 2012, 7, e32678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-kappa B and Rel proteins: Evolutionarily conserved mediators of immune responses. Annu. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef]
- Holmes-McNary, M.; Baldwin, A.S., Jr. Chemopreventive properties of trans-resveratrol are associated with inhibition of activation of the IkappaB kinase. Cancer Res. 2000, 60, 3477–3483. [Google Scholar] [PubMed]
- Ren, Z.; Wang, L.; Cui, J.; Huoc, Z.; Xue, J.; Cui, H.; Mao, Q.; Yang, R. Resveratrol inhibits NF-kB signaling through suppression of p65 and IkappaB kinase activities. Die Pharm.-Int. J. Pharm. Sci. 2013, 68, 689–694. [Google Scholar]
- Tsai, S.H.; Lin-Shiau, S.Y.; Lin, J.K. Suppression of nitric oxide synthase and the down-regulation of the activation of NFkappaB in macrophages by resveratrol. Br. J. Pharmacol. 1999, 126, 673–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatt, L.; Saliba, D.G.; Schembri-Wismayer, P.; Zammit-Mangion, M. Tyrosol, at the Concentration Found in Maltese Extra Virgin Olive Oil, Induces HL-60 Differentiation towards the Monocyte lineage. Appl. Sci. 2021, 11, 10199. https://doi.org/10.3390/app112110199
Gatt L, Saliba DG, Schembri-Wismayer P, Zammit-Mangion M. Tyrosol, at the Concentration Found in Maltese Extra Virgin Olive Oil, Induces HL-60 Differentiation towards the Monocyte lineage. Applied Sciences. 2021; 11(21):10199. https://doi.org/10.3390/app112110199
Chicago/Turabian StyleGatt, Lucienne, David G. Saliba, Pierre Schembri-Wismayer, and Marion Zammit-Mangion. 2021. "Tyrosol, at the Concentration Found in Maltese Extra Virgin Olive Oil, Induces HL-60 Differentiation towards the Monocyte lineage" Applied Sciences 11, no. 21: 10199. https://doi.org/10.3390/app112110199