
Drought Stress during Anthesis Alters Grain Protein Composition and Improves Bread Quality in Field-Grown Iranian and German Wheat Genotypes


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genotype Characteristics
2.2. Trial Location, Experimental Design and Growing Conditions
2.3. Yield and Total Protein Concentration
2.4. Extraction of Cereal Proteins
2.5. Quantitative Analysis of the Protein Fractions
2.6. Qualitative Analysis of the Protein Fractions by SDS-PAGE
2.7. Evaluation of SDS Gels
2.8. Flour and Baking Test
2.9. Statistical Methods
3. Results
3.1. Yield and Total Protein Concentration of the Grain
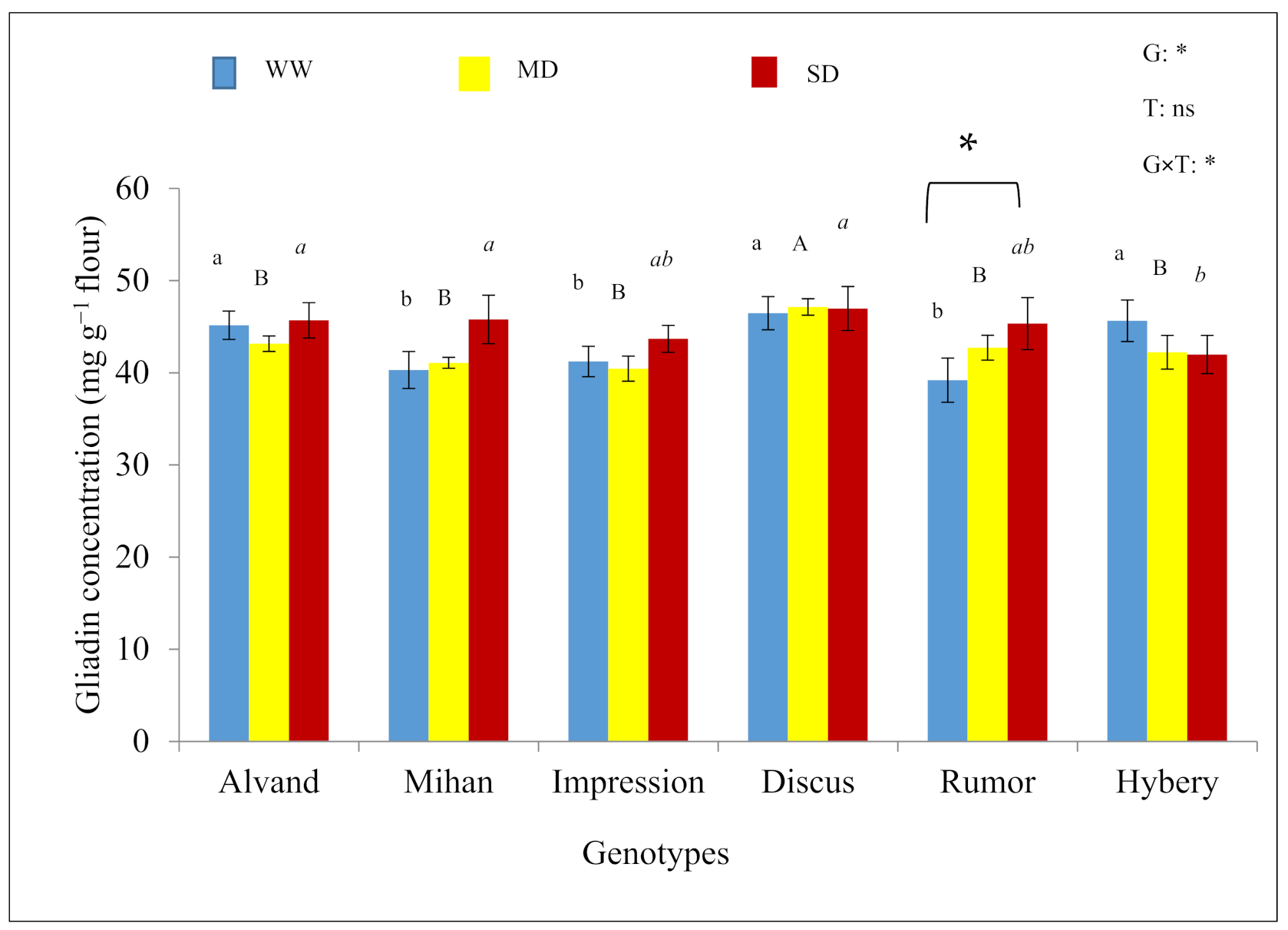
3.2. Concentration of Different Fractions
3.3. Composition of the Grain Protein Fractions
3.4. Baking Quality
3.5. Principal Component Analysis
4. Discussion
4.1. Drought-Induced Improved Baking Quality Is Related to Changes in Protein Composition Rather Than Total Protein Content
4.2. Baking Quality Might Be Somewhat Negatively Related to Drought-Induced Increases in γ-Gliadins and LMW-D Glutenins
4.3. Iranian Genotypes Are Not Generally More Tolerant to Severe Drought Stress Than German Genotypes
4.4. Drought Tolerant Iranian Genotypes Have a High Baking Quality and Might Be Suitable Candidates for Drought-Prone Areas in Germany
4.5. Conclusions and Outlook for Further Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatments | ||
|---|---|---|---|
| WW | MD | SD | |
| ω-Gliadin (% of the total gliadin content) | |||
| Alvand | 14.9 b | 15.5 b | 17.0 ab |
| Mihan | 12.4 d | 14.6 c | 15.5 c |
| Impression | 16.4 a | 17.2 a | 17.4 a |
| Discus | 11.3 e | 12.8 d | 14.4 d |
| Rumor | 10.7 f | 12.0 e | 13.1 e |
| Hybery | 13.3 c | 14.4 c | 16.5 b |
| HMW Glutenin (% of the total glutenin content) | |||
| Alvand | 27.3 d | 27.2 e | 28.7 e |
| Mihan | 33.3 b | 35.2 b | 35.2 c |
| Impression | 41.3 a | 41.4 a | 48.0 a |
| Discus | 27.3 d | 29.2 d | 32.6 d |
| Rumor | 32.0 bc | 32.4 c | 36.0 c |
| Hybery | 31.2 c | 30.9 cd | 41.2 b |
| LMW-C Glutenin (% of the total glutenin content) | |||
| Alvand | 37.2 a | 37.9 a | 35.3 a |
| Mihan | 27.5 c | 29.2 cd | 28.8 b |
| Impression | 30.6 b | 31.9 b | 22.8 d |
| Discus | 31.4 b | 31.0 bc | 25.4 c |
| Rumor | 28.5 c | 27.8 d | 22.7 d |
| Hybery | 30.7 b | 30.8 bc | 23.2 d |
| Specific volume (mL/g) | |||
| Alvand | 3.3 b | 3.4 b | 3.7 b |
| Mihan | 2.9 d | 3.3 bc | 3.4 c |
| Impression | 3.5 a | 3.6 a | 3.8 a |
| Discus | 2.8 e | 3.0 d | 3.3 d |
| Rumor | 2.8 e | 2.9 e | 3.0 e |
| Hybery | 3.2 c | 3.2 c | 3.4 c |

References
- Klingler, R.W. Grundlagen der Getreidetechnologie, 2nd ed.; Behr’s GmbH: Hamburg/Berlin, Germany, 2010. [Google Scholar]
- Osborne, T.B. The Proteins of the Wheat Kernel; Nabu Press: Washington, DC, USA, 1907; Available online: https://archive.org/details/proteinsofwheatk00osborich/page/n5/mode/2up (accessed on 13 October 2021).
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeranz, Y. Composition and functionally of wheat flour components. In Wheat: Chemistry and Technology, 2nd ed.; Pomeranz, Y., Ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 1988; Volume 2, pp. 219–370. [Google Scholar]
- Bietz, J.A.; Wall, J.S. Wheat Gluten Subunits: Molecular Weights Determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Cereal Chem. 1972, 49, 416–430. [Google Scholar]
- Jackson, E.A.; Holt, L.M.; Payne, P.I. Characterisation of high molecular weight gliadin and low-molecular-weight glutenin subunits of wheat endosperm by two-dimensional electrophoresis and the chromosomal localisation of their controlling genes. Theor. Appl. Genet. 1983, 66, 29–37. [Google Scholar] [CrossRef] [PubMed]
- DuPont, F.M.; Altenbach, S.B. Molecular and biochemical impacts of environmental factors on wheat grain development and protein synthesis. J. Cereal Sci. 2003, 38, 133–146. [Google Scholar] [CrossRef]
- Rekowski, A.; Wimmer, M.A.; Henkelmann, G.; Zörb, C. Is a change of protein composition after late application of nitrogen sufficient to improve the baking quality of winter wheat? Agriculture 2019, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Souza, E.J.; Martin, J.M.; Guttieri, M.J.; O’Brien, K.M.; Habernicht, D.K.; Lanning, S.P.; McLean, R.; Carlson, G.R.; Talbert, L.E. Influence of genotype, environment, and nitrogen management on spring wheat quality. Crop. Sci. 2004, 44, 425. [Google Scholar] [CrossRef]
- Pauk, J.; Lantos, C.; Mihály, R.; Flamm, C.; Livaja, M.; Cseuz, L.; Schmolke, M.; Ruthner, S. Greenhouse Test for Drought Tolerance of the CORNET Wheat Variety Set. In Von Markergestutzter Selektion zu Genomischer Selektion in der Pflanzenzüchtung, 62. Jahrestagung der Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs, Irdning, Austria, 22–23 November 2011; Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs, Ed.; Raumberg-Gumpenstein: Irdning, Austria, 2012; pp. 43–48. [Google Scholar]
- Liu, Y.; Liang, H.; Lv, X.; Liu, D.; Wen, X.; Liao, Y. Effect of polyamines on the grain filling of wheat under drought stress. Plant Physiol. Biochem. 2016, 100, 113–129. [Google Scholar] [CrossRef]
- Hernández-Espinosa, N.; Mondal, S.; Autrique, E.; Gonzalez-Santoyo, H.; Crossa, J.; Huerta-Espino, J.; Singh, R.P.; Guzmán, C. Milling, processing and end-use quality traits of CIMMYT spring bread wheat germplasm under drought and heat stress. Field Crop. Res. 2018, 215, 104–112. [Google Scholar] [CrossRef]
- Blumenthal, C.S.; Barlow, E.W.R.; Wrigley, C.W. Growth environment and wheat quality: The effect of heat stress on dough properties and gluten proteins. J. Cereal Sci. 1993, 18, 3–21. [Google Scholar] [CrossRef]
- Blumenthal, C.S.; Bekes, F.; Gras, P.W.; Barlow, E.W.R.; Wrigley, C.W. Identification of wheat genotypes tolerant to the effects of heat stress on grain quality. Cereal Chem. 1995, 72, 539–544. [Google Scholar]
- Daniel, C.; Triboï, E. Changes in wheat protein aggregation during grain development: Effects of temperatures and water stress. Eur. J. Agron. 2002, 16, 1–12. [Google Scholar] [CrossRef]
- Shamsi, K.; Petrosyan, M.; Noor-Mohammadi, G.; Haghparast, A.; Kobraee, S.; Rasekhi, B. Differential agronomic responses of bread wheat cultivars to drought stress in the west of Iran. Afr. J. Biotechnol. 2011, 10, 2708–2715. [Google Scholar] [CrossRef] [Green Version]
- Reeves, T.G.; Rajaram, S.; Ginkel, M.V.; Trethowan, R.; Braun, H.J.; Cassaday, R. New Wheats for a Secure, Sustainable Future; CIMMYT: Mexico City, Mexico, 1999; ISBN 970-648-040-4. [Google Scholar]
- Zörb, C.; Becker, E.; Merkt, N.; Kafka, S.; Schmidt, S.; Schmidhalter, U. Shift of grain protein composition in bread wheat under summer drought events. J. Plant Nutr. Soil Sci. 2017, 180, 49–55. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Osborne, T.B. The Vegetable Proteins. In Monographs on Biochemistry, 2nd ed.; Longmans Green and Company: London, UK, 1924. [Google Scholar]
- Wieser, H.; Seilmeier, W. The influence of nitrogen fertilization on quantities and proportions of different protein types in wheat flour. J. Sci. Food Agric. 1998, 76, 49–55. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G.; Belton, P.S.; Tatham, A.S. The structure and properties of gluten. An elastic protein from wheat grain. Philos. Trans. R. Soc. B 2002, 357, 133–142. [Google Scholar] [CrossRef]
- Rekowski, A.; Langenkämper, G.; Dier, M.; Wimmer, M.A.; Scherf, K.A.; Zörb, C. Determination of soluble wheat protein fractions using the Bradford assay. Cereal Chem. 2021, 98, 1059–1065. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Kieffer, R.; Belitz, H.D.; Zweier, M.; Ipfelkofer, R.; Fischbeck, G. Der Rapid-Mix-Test als 10-g-Mikrobackversuch. Eur. Food Res. Technol. 1993, 197, 134–136. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Stark, J.C.; O’Brien, K.; Souza, E. Relative Sensitivity of Spring Wheat Grain Yield and Quality Parameters to Moisture Deficit. Crop. Sci. 2001, 41, 327–335. [Google Scholar] [CrossRef]
- Zörb, C.; Langenkämper, G.; Betsche, T.; Niehaus, K.; Barsch, A. Metabolite Profiling of Wheat Grains (Triticum aestivum L.) from Organic and Conventional Agriculture. J. Agric. Food Chem. 2006, 54, 8301–8306. [Google Scholar] [CrossRef]
- Xie, Z.; Jiang, D.; Cao, W.; Dai, T.; Jing, Q. Relationships of endogenous plant hormones to accumulation of grain protein and starch in winter wheat under different post-anthesis soil water statuses. Plant Growth Regul. 2003, 41, 117–127. [Google Scholar] [CrossRef]
- Guóth, A.; Tari, I.; Gallé, Á.; Csiszár, J.; Pécsváradi, A.; Cseuz, L.; Erdei, L. Comparison of the Drought Stress Responses of Tolerant and Sensitive Wheat Cultivars During Grain Filling: Changes in Flag Leaf Photosynthetic Activity, ABA Levels, and Grain Yield. J. Plant Growth Regul. 2009, 28, 167–176. [Google Scholar] [CrossRef]
- Field, J.M.; Shewry, P.R.; Miflin, B.J. Solubilisation and characterisation of wheat gluten proteins: Correlations between the amount of aggregated proteins and baking quality. J. Sci. Food Agric. 1983, 34, 370–377. [Google Scholar] [CrossRef]
- Fido, R.J.; Békés, F.; Gras, P.W.; Tatham, A.S. Effects of α-, β-, γ- and ω-Gliadins on the Dough Mixing Properties of Wheat Flour. J. Cereal Sci. 1997, 26, 271–277. [Google Scholar] [CrossRef]
- Ebermann, R.; Elmadfa, I. Lehrbuch Lebensmittelchemie und Ernährung, 2nd ed.; Springer: Vienna, Austria, 2011; Available online: http://site.ebrary.com/lib/alltitles/docDetail.action?docID=10494376 (accessed on 13 October 2021).
- Shewry, P.R.; Tatham, A.S.; Forde, J.; Kreis, M.; Miflin, B.J. The classification and nomenclature of wheat gluten proteins: A reassessment. J. Cereal Sci. 1986, 4, 97–106. [Google Scholar] [CrossRef]
- Triboï, E.; Martre, P.; Triboï-Blondel, A. Environmentally-induced changes in protein composition in developing grains of wheat are related to changes in total protein content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef]





| Genotype | Quality Level | Company | Protein Concentr. | Drought Resistance |
|---|---|---|---|---|
| Iranian genotype | ||||
| Alvand | class 1 | National Seed and Plant Improvement Institute (SPII) | high | sensitive |
| Mihan | class 2 | low | tolerant | |
| German genotype | ||||
| Impression | A | I.G. Pflanzenzucht | high | sensitive |
| Discus | A | DSV | high | sensitive |
| Rumor | B | Saaten-Union | low | tolerant |
| Hybery | B | Saaten-Union | low | tolerant |
| Genotype | Alvand | Mihan | Impression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| ω-Gliadin | 14.9 b | 15.5 b | 17.0 a | 12.4 c | 14.6 b | 15.5 a | 16.4 b | 17.2 ab | 17.4 a |
| γ-Gliadin | 36.1 | 37.0 | 48.1 | 34.6 Ab | 31.9 B | 35.5 A | 23.2 | 23.7 | 24.8 |
| α/β-Gliadin | 45.7 | 48.1 | 48.4 | 53.0 a | 53.5 a | 48.7 b | 60.4 | 59.1 | 57.8 |
| Σ | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Genotype | Discus | Rumor | Hybery | ||||||
| Treatments | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| ω-Gliadin | 11.3 c | 12.8 b | 14.4 a | 10.7 c | 12.0 b | 13.1 a | 13.3 c | 14.4 b | 16.5 a |
| γ-Gliadin | 28.2 B | 30.9 Ab | 31.5 A | 29.2 B | 30.9 B | 34.1 A | 24.9 | 25.1 | 23.9 |
| α/β-Gliadin | 60.5 a | 56.2 b | 54.1 b | 60.1 a | 57.1 a | 52.7 b | 61.8 | 60.6 | 59.7 |
| Σ | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Genotype | Alvand | Mihan | Impression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| HMW | 27.3 | 27.2 | 28.7 | 33.3 | 35.2 | 35.2 | 41.3 b | 41.4 b | 48.0 a |
| LMW-D | 4.3 | 4.6 | 5.0 | - | - | - | 1.6 | 1.5 | 1.0 |
| LMW-B | 31.2 | 30.2 | 31.0 | 39.2 | 35.6 | 36.0 | 26.7 | 28.0 | 28.1 |
| LMW-C | 37.2 ab | 37.9 a | 35.3 b | 27.5 | 29.2 | 28.8 | 30.6 a | 31.9 a | 22.8 b |
| Σ | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| HMW/LMW | 0.37 | 0.37 | 0.40 | 0.50 | 0.54 | 0.54 | 0.70 b | 0.67 b | 0.92 a |
| Genotype | Discus | Rumor | Hybery | ||||||
| Treatments | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| HMW | 27.3 b | 29.2 b | 32.6 a | 32.0 b | 32.4 b | 36.0 a | 31.2 b | 30.9 b | 41.2 a |
| LMW-D | 5.1 B | 5.3 B | 6.8 A | 6.4 B | 6.5 B | 10.6 A | 5.2 | 4.8 | 3.9 |
| LMW-B | 36.3 | 34.5 | 35.2 | 32.8 ab | 33.7 a | 30.8 b | 32.9 | 33.4 | 31.7 |
| LMW-C | 31.4 a | 31.0 a | 25.4 b | 28.5 a | 27.8 a | 22.7 b | 30.7 a | 30.8 a | 23.2 b |
| Σ | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| HMW/LMW | 0.37 b | 0.41 b | 0.48 a | 0.47 b | 0.48 b | 0.56 a | 0.45 b | 0.45 b | 0.70 a |
| Genotype | Alvand | Mihan | Impression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| Specific volume (mL/g) | 3.3 b | 3.4 b | 3.7 a | 2.9 b | 3.3 a | 3.4 a | 3.5 b | 3.6 b | 3.8 a |
| Freshness retention (N) | 13.1 | 16.7 | 15.6 | 14.8 | 12.7 | 12.4 | 17.1 | 15.3 | 15.4 |
| Hardness (N) | 4.6 | 5.1 | 4.6 | 5.1 | 4.4 | 4.0 | 5.0 | 4.9 | 4.6 |
| Genotype | Discus | Rumor | Hybery | ||||||
| Treatments | WW | MD | SD | WW | MD | SD | WW | MD | SD |
| Specific volume (mL/g) | 2.8 c | 3.0 b | 3.3 a | 2.8 b | 2.9 b | 3.0 a | 3.2 b | 3.2 b | 3.4 a |
| Freshness retention (N) | 17.0 | 16.2 | 13.8 | 18.6 A | 13.8 BB | 13.3 B | 15.4 | 14.4 | 14.0 |
| Hardness (N) | 6.0 a | 5.0 ab | 4.2 b | 6.1 | 5.6 | 5.1 | 5.2 | 4.4 | 4.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rekowski, A.; Wimmer, M.A.; Tahmasebi, S.; Dier, M.; Kalmbach, S.; Hitzmann, B.; Zörb, C. Drought Stress during Anthesis Alters Grain Protein Composition and Improves Bread Quality in Field-Grown Iranian and German Wheat Genotypes. Appl. Sci. 2021, 11, 9782. https://doi.org/10.3390/app11219782
Rekowski A, Wimmer MA, Tahmasebi S, Dier M, Kalmbach S, Hitzmann B, Zörb C. Drought Stress during Anthesis Alters Grain Protein Composition and Improves Bread Quality in Field-Grown Iranian and German Wheat Genotypes. Applied Sciences. 2021; 11(21):9782. https://doi.org/10.3390/app11219782
Chicago/Turabian StyleRekowski, Azin, Monika A. Wimmer, Sirous Tahmasebi, Markus Dier, Sarah Kalmbach, Bernd Hitzmann, and Christian Zörb. 2021. "Drought Stress during Anthesis Alters Grain Protein Composition and Improves Bread Quality in Field-Grown Iranian and German Wheat Genotypes" Applied Sciences 11, no. 21: 9782. https://doi.org/10.3390/app11219782
APA StyleRekowski, A., Wimmer, M. A., Tahmasebi, S., Dier, M., Kalmbach, S., Hitzmann, B., & Zörb, C. (2021). Drought Stress during Anthesis Alters Grain Protein Composition and Improves Bread Quality in Field-Grown Iranian and German Wheat Genotypes. Applied Sciences, 11(21), 9782. https://doi.org/10.3390/app11219782