
Novel Lipid-Oligonucleotide Conjugates Containing Long-Chain Sulfonyl Phosphoramidate Groups: Synthesis and Biological Properties

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oligonucleotide Synthesis
2.2. Dynamic Light Scattering (DLS) Characterization
2.3. Cell Lines
2.4. MTT Assay of Cell Viability
2.5. Flow Cytometry
2.6. Confocal Microscopy
2.7. Gene Silencing
3. Results
3.1. Synthesis of Lipid-Oligonucleotide Conjugates (LONs) Incorporating Long-Chain (Sulfonyl)Phosphoramidate Groups
3.2. Spontaneous Nanoparticle Formation by LONs in Aqueous Media
3.3. LONs were Nontoxic to Murine or Human Macrophages at Therapeutic Concentrations
3.4. Conjugates with Longer Lipidic Chains Accumulated in Cells with Higher Efficiency
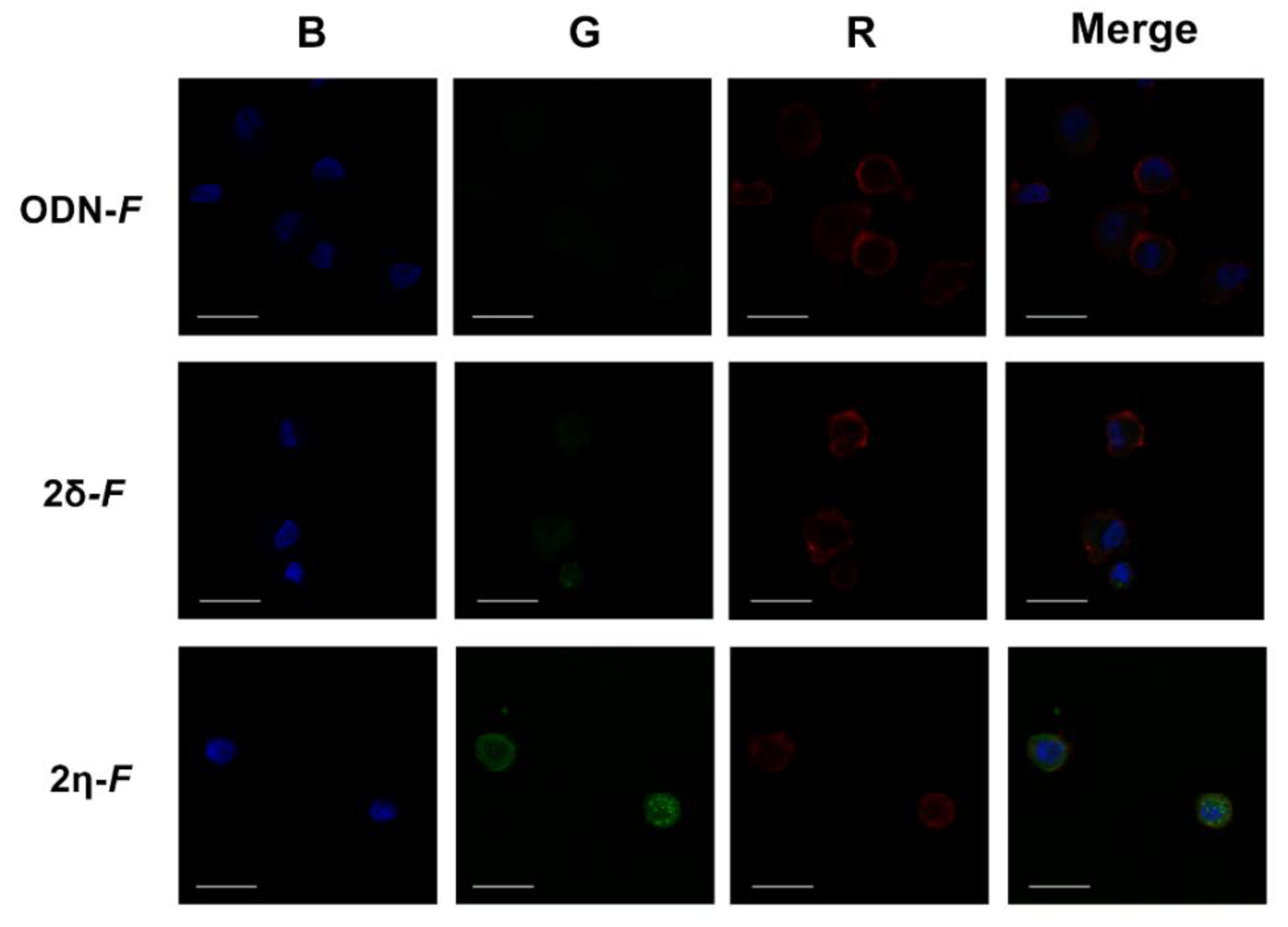
3.5. Lipid-Oligonucleotide Conjugates are Efficiently Taken up by Cells in the Absence of Transfection Agents and Localize Mostly in the Cytoplasm within Endosomes
3.6. Lipid-Conjugated RGMa siRNAs Demonstrated Gene-Silencing Ability Similar to Unmodified RGMa siRNA in PK-59 Human Pancreatic Carcinoma Cell Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Crooke, S.T. Molecular mechanisms of antisense oligonucleotides. Nucleic Acid Ther. 2017, 27, 70–77. [Google Scholar] [CrossRef]
- Shen, X.; Corey, D.R. Chemistry, mechanism and clinical status of antisense oligonucleotides and duplex RNAs. Nucleic Acids Res. 2018, 46, 1584–1600. [Google Scholar] [CrossRef]
- Smith, C.I.E.; Zain, R. Therapeutic Oligonucleotides: State of the Art. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 605–630. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.M.; Toonen, L.J.A.; van Roon-Mom, W.M.C. Antisense oligonucleotides in therapy for neurodegenerative disorders. Adv. Drug Deliv. Rev. 2015, 87, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung-Htut, M.T.; McIntosh, C.S.; Ham, K.A.; Pitout, I.L.; Flynn, L.L.; Greer, K.; Fletcher, S.; Wilton, S.D. Systematic Approach to Developing Splice Modulating Antisense Oligonucleotides. Int. J. Mol. Sci. 2019, 20, 5030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.F.; Baker, B.F.; Pham, N.; Swayze, E.; Geary, R.S. Pharmacology of Antisense Drugs. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 81–105. [Google Scholar] [CrossRef]
- Castanotto, D.; Stein, C.A. Antisense oligonucleotides in cancer. Curr. Opin. Oncol. 2014, 26, 584–589. [Google Scholar] [CrossRef]
- Wagner, A.; Bock, C.T.; Fechner, H.; Kurreck, J. Application of modified antisense oligonucleotides and siRNAs as antiviral drugs. Future Med. Chem. 2015, 7, 1637–1642. [Google Scholar] [CrossRef]
- Hegarty, J.P.; Stewart, D.B. Advances in therapeutic bacterial antisense biotechnology. Appl. Microbiol. Biotechnol. 2018, 102, 1055–1065. [Google Scholar] [CrossRef]
- Shimo, T.; Maruyama, R.; Yokota, T. Designing Effective Antisense Oligonucleotides for Exon Skipping. In Duchenne Muscular Dystrophy. Methods and Protocols; Bernardini, C., Ed.; Humana Press: New York, NY, USA, 2018. [Google Scholar]
- Ma, C.-C.; Wang, Z.-L.; Xu, T.; He, Z.-Y.; Wei, Y.-Q. The approved gene therapy drugs worldwide: From 1998 to 2019. Biotechnol. Adv. 2020, 40, 107502. [Google Scholar] [CrossRef]
- Juliano, R.L. The delivery of therapeutic oligonucleotides. Nucleic Acids Res. 2016, 44, 6518–6548. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug. Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, S.; Khatra, H.; Saha, S.; Sinha, S. A cationic morpholino antisense oligomer conjugate: Synthesis, cellular uptake and inhibition of Gli1 in the hedgehog signalling pathway. RSC Adv. 2014, 4, 1951–1954. [Google Scholar] [CrossRef]
- Hnedzko, D.; McGee, D.W.; Karamitas, Y.A.; Rozners, E. Sequence-selective recognition of double-stranded RNA and enhanced cellular uptake of cationic nucleobase and backbone-modified peptide nucleic acids. RNA 2017, 23, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wu, B.; Shah, S.N.; Lu, P.; Lu, Q. Polyquaternium-mediated delivery of morpholino oligonucleotides for exon-skipping in vitro and in mdx mice. Drug Deliv. 2017, 24, 952–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morcos, P.A.; Li, Y.; Jiang, S. Vivo-Morpholinos: A non-peptide transporter delivers Morpholinos into a wide array of mouse tissues. Biotechniques 2018, 45, 613–623. [Google Scholar] [CrossRef]
- Craig, K.; Abrams, M.; Amiji, M. Recent preclinical and clinical advances in oligonucleotide conjugates. Expert Opin. Drug Deliv. 2018, 15, 629–640. [Google Scholar] [CrossRef]
- Ren, T.; Song, Y.K.; Zhang, G.; Liu, D. Structural basis of DOTMA for its high intravenous transfection activity in mouse. Gene Ther. 2000, 7, 764–768. [Google Scholar] [CrossRef] [Green Version]
- Patwa, A.; Gissot, A.; Bestel, I.; Barthélémy, P. Hybrid lipid oligonucleotide conjugates: Synthesis, self-assemblies and biomedical applications. Chem. Soc. Rev. 2011, 40, 5844–5854. [Google Scholar] [CrossRef]
- Zhao, B.; Tian, Q.; Bagheri, Y.; You, M. Lipid–oligonucleotide conjugates for simple and efficient cell membrane engineering and bioanalysis. Curr. Opin. Biomed. Eng. 2020, 13, 76–83. [Google Scholar] [CrossRef]
- Li, X.; Feng, K.; Li, L.; Yang, L.; Pan, X.; Yazd, H.S.; Cui, C.; Li, J.; Moroz, L.; Sun, Y.; et al. Lipid-oligonucleotide conjugates for bioapplications. Nat. Sci. Rev. 2020, 7, 1933–1953. [Google Scholar] [CrossRef]
- Osborn, M.F.; Khvorova, A. Improving siRNA Delivery In Vivo Through Lipid Conjugation. Nucleic Acid Ther. 2018, 28, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Østergaard, M.E.; Jackson, M.; Low, A.; Chappell, A.E.; Lee, R.G.; Peralta, R.Q.; Yu, J.; Kinberger, G.A.; Dan, A.; Carty, R.; et al. Conjugation of hydrophobic moieties enhances potency of antisense oligonucleotides in the muscle of rodents and non-human primates. Nucleic Acids Res. 2019, 47, 6045–6058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernikov, I.V.; Meschaninova, M.I.; Chernolovskaya, E.L. Preparation, Determination of Activity, and Biodistribution of Cholesterol-Containing Nuclease-Resistant siRNAs In Vivo. In RNA Interference and CRISPR Technologies. Technical Advances and New Therapeutic Opportunities; Sioud, M., Ed.; Humana Press: New York, NY, USA, 2020. [Google Scholar]
- Prokhorova, D.V.; Chelobanov, B.P.; Burakova, E.A.; Fokina, A.A.; Stetsenko, D.A. New oligodeoxyribonucleotide derivatives bearing internucleotide N-tosyl phosphoramidate groups: Synthesis and complementary binding to DNA and RNA. Russ. J. Bioorg. Chem. 2017, 43, 38–42. [Google Scholar] [CrossRef]
- Chelobanov, B.P.; Burakova, E.A.; Prokhorova, D.V.; Fokina, A.A.; Stetsenko, D.A. New oligodeoxynucleotide derivatives containing N-(methanesulfonyl)-phosphoramidate (mesyl phosphoramidate) internucleotide group. Russ. J. Bioorg. Chem. 2017, 43, 664–668. [Google Scholar] [CrossRef]
- Burakova, E.A.; Derzhalova, A.S.; Chelobanov, B.P.; Fokina, A.A.; Stetsenko, D.A. New Oligodeoxynucleotide Derivatives Containing N-(Sulfonyl)-Phosphoramide Groups. Russ. J. Bioorg. Chem. 2017, 45, 662–668. [Google Scholar] [CrossRef]
- Kitayama, M.; Ueno, M.; Itakura, T.; Yamashita, T. Activated Microglia Inhibit Axonal Growth through RGMa. PLoS ONE 2011, 6, e25234. [Google Scholar] [CrossRef]
- Farzan, V.M.; Ulashchik, E.A.; Martynenko-Makaev, Y.V.; Kvach, M.V.; Aparin, I.O.; Brylev, V.A.; Prikazchikova, T.A.; Maklakova, S.Y.; Majouga, A.G.; Ustinov, A.V.; et al. Automated Solid-Phase Click Synthesis of Oligonucleotide Conjugates: From Small Molecules to Diverse N-Acetylgalactosamine Clusters. Bioconjug. Chem. 2017, 28, 2599–2607. [Google Scholar] [CrossRef]
- Matano, Y.; Ohkubo, H.; Honsho, Y.; Saito, A.; Seki, S.; Imahori, H. Synthesis and Charge-Carrier Transport Properties of Poly(phosphole P-alkanesulfonylimide)s. Org. Lett. 2013, 15, 932–935. [Google Scholar] [CrossRef]
- Kamerkar, S.; LeBleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef]
- Song, E.; Lee, S.K.; Wang, J.; Ince, N.; Ouyang, N.; Min, J.; Chen, J.; Lieberman, S.P. RNA interference targeting Fas protects mice from fulminant hepatitis. J. Nat. Med. 2003, 9, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Markov, O.V.; Filatov, A.V.; Kupryushkin, M.S.; Chernikov, I.V.; Patutina, O.A.; Strunov, A.A.; Chernolovskaya, E.L.; Vlassov, V.V.; Pyshnyi, D.V.; Zenkova, M.A. Transport Oligonucleotides—A Novel System for Intracellular Delivery of Antisense Therapeutics. Molecules 2020, 25, 3663. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T.; Tokita, S.; Yamashita, T. Repulsive guidance molecule-a and demyelination: Implications for multiple sclerosis. J. Neuroimmune Pharmacol. 2012, 7, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Mothe, A.J.; Tassew, N.G.; Shabanzadeh, A.P.; Penheiro, R.; Vigouroux, R.J.; Huang, L.; Grinnell, C.; Cui, Y.F.; Fung, E.; Monnier, P.P.; et al. RGMa inhibition with human monoclonal antibodies promotes regeneration, plasticity and repair, and attenuates neuropathic pain after spinal cord injury. Sci. Rep. 2017, 7, 10529. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, S.K.; Patutina, O.A.; Burakova, E.A.; Chelobanov, B.P.; Fokina, A.A.; Vlassov, V.V.; Altman, S.; Zenkova, M.A.; Stetsenko, D.A. Mesyl phosphoramidate antisense oligonucleotides as an alternative to phosphorothioates: Improved biochemical and biological properties. Proc. Natl. Acad. Sci. USA 2019, 116, 1229–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patutina, O.A.; Gaponova (Miroshnichenko), S.K.; Sen’kova, A.V.; Savin, I.A.; Gladkikh, D.V.; Burakova, E.A.; Fokina, A.A.; Maslov, M.A.; Shmendel’, E.V.; Wood, M.J.A.; et al. Mesyl phosphoramidate backbone modified antisense oligonucleotides targeting miR-21 with enhanced in vivo therapeutic potency. Proc. Natl. Acad. Sci. USA 2020, 117. in press. [Google Scholar] [CrossRef]
- Kupryushkin, M.S.; Pyshnyi, D.V.; Stetsenko, D.A. Phosphoryl Guanidines: A New Type of Nucleic Acid Analogues. Acta Nat. 2014, 6, 116–118. [Google Scholar] [CrossRef]
- Ezzat, K.; Aoki, Y.; Koo, T.; McClorey, G.; Benner, L.; Coenen-Stass, A.; O’Donovan, L.; Lehto, T.; Garcia-Guerra, A.; Nordin, J.; et al. Self-Assembly into Nanoparticles Is Essential for Receptor Mediated Uptake of Therapeutic Antisense Oligonucleotides. Nano Lett. 2015, 15, 4364–4373. [Google Scholar] [CrossRef] [Green Version]
- Deo, S.; Patel, T.R.; Dzananovic, E.; Booy, E.P.; Zeid, K.; McEleney, K.; Harding, S.E.; McKenna, S.A. Activation of 2′5′-Oligoadenylate Synthetase by Stem Loops at the 5′-End of the West Nile Virus Genome. PLoS ONE 2014, 9, e92545. [Google Scholar] [CrossRef] [Green Version]
- Awino, J.K.; Gudipati, S.; Hartmann, A.K.; Santiana, J.J.; Cairns-Gibson, D.F.; Gomez, N.; Rouge, J.L. Nucleic Acid Nanocapsules for Enzyme-Triggered Drug Release. J. Am. Chem. Soc. 2017, 139, 6278–6281. [Google Scholar] [CrossRef]
- Soutschek, J.; Akinc, A.; Bramlage, B.; Charisse, K.; Constien, R.; Donoghue, M.; Elbashir, S.; Geick, A.; Hadwiger, P.; Harborth, J.; et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature 2004, 432, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Benizri, S.; Mejías, R.; Baylot, V.; Branger, N.; Nguyen, T.; Vialet, B.; Oumzil, K.; Barthélémy, P.; Rocchi, P. Lipid-oligonucleotide conjugates improve cellular uptake and efficiency of TCTP-antisense in castration-resistant prostate cancer. J. Control. Release 2017, 258, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, F.; Liu, Y.; Zheng, M.; Wang, Y.; Zhang, D.; Anraku, Y.; Zou, Y.; Li, J.; Wu, H.; et al. Blood-brain barrier-penetrating siRNA nanomedicine for Alzheimer’s disease therapy. Sci. Adv. 2020, 6, eabc7031. [Google Scholar] [CrossRef] [PubMed]
- Sathyaprakash, C.; Manzano, R.; Varela, M.A.; Hashimoto, Y.; Wood, M.J.A.; Talbot, K.; Aoki, Y. Development of LNA Gapmer Oligonucleotide-Based Therapy for ALS/FTD Caused by the C9orf72 Repeat Expansion. In Gapmers: Methods and Protocols; Yokota, T., Maruyama, R., Eds.; Springer: New York, NY, USA, 2020; pp. 185–208. [Google Scholar]
- Scoles, D.R.; Meera, P.; Schneider, M.D.; Paul, S.; Dansithong, W.; Figueroa, K.P.; Hung, G.; Rigo, F.; Bennett, C.F.; Otis, T.S.; et al. Antisense oligonucleotide therapy for spinocerebellar ataxia type 2. Nature 2017, 544, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Givosiran: First Approval. Drugs 2020, 80, 335–339. [Google Scholar] [CrossRef]
- Heo, Y.-A. Golodirsen: First Approval. Drugs 2020, 80, 329–333. [Google Scholar] [CrossRef]
- Paik, J.; Duggan, S. Volanesorsen: First Global Approval. Drugs 2019, 79, 1349–1354. [Google Scholar] [CrossRef]
- Keam, S.J. Inotersen: First Global Approval. Drugs 2018, 78, 1371–1376. [Google Scholar] [CrossRef]
- Remaut, K.; Lucas, B.; Braeckmans, K.; Demeester, J.; De Smedt, S.C. Pegylation of liposomes favours the endosomal degradation of the delivered phosphodiester oligonucleotides. J. Control. Release 2007, 117, 256–266. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Z.; Chen, H.; Gao, J.; Liang, W. Transfection efficiency and intracellular fate of polycation liposomes combined with protamine. Biomaterials 2011, 32, 1412–1418. [Google Scholar] [CrossRef] [PubMed]
- Ara, M.N.; Matsuda, T.; Hyodo, M.; Sakurai, Y.; Hatakeyama, H.; Ohga, N.; Hida, K.; Harashima, H. An aptamer ligand based liposomal nanocarrier system that targets tumor endothelial cells. Biomaterials 2014, 35, 7110–7120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal escape pathways for delivery of biologicals. J. Control. Release 2011, 151, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.S.; Chernikov, I.V.; Meschaninova, M.I.; Dovydenko, I.S.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Carrier-free cellular uptake and the gene-silencing activity of the lipophilic siRNAs is strongly affected by the length of the linker between siRNA and lipophilic group. Nucleic Acids Res. 2012, 40, 2330–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Designation | Oligonucleotide Sequence, 5′–3′ 1 | Molecular Mass, Da2 | |
|---|---|---|---|---|
| Calc. [M] | Exp. [M] | |||
| Model antisense oligonucleotide | ODN | agtctcgacttgctacc 3 | 5121.25 | 5120.98 |
| ODN-F 4 | agtctcgacttgctacc-Flu | 5688.74 | 5687.93 | |
| 1δ | aδgtctcgacttgctacc | 5248.86 | 5426.08 | |
| 2δ | aδgδtctcgacttgctacc | 5736.36 | 5733.28 | |
| 1δ-F | aδgtctcgacttgctacc-Flu | 5996.35 | 5993.21 | |
| 2δ-F | aδgδtctcgacttgctacc-Flu | 6303.85 | 6300.41 | |
| 1η | aηgtctcgacttgctacc | 5408.87 | 5406.11 | |
| 2η | aηgηtctcgacttgctacc | 5696.38 | 5693.34 | |
| 1η-F | aηgtctcgacttgctacc-Flu | 5976.36 | 5973.24 | |
| 2η-F | aηgηtctcgacttgctacc-Flu | 6263.87 | 6260.47 | |
| siRNA | EGFP | Sense CCUACGGCAAGCUGACCCUGA | 6698.10 | 6695.84 |
| Antisense AGGGUCAGCUUGCCGUAGGUG | 6812.10 | 6811.57 | ||
| RGMa | Sense CCAUCAUCUUCAAGAACUUCC | 6563.99 | 6565.38 | |
| Antisense AAGUUCUUGAAGAUGAUGGUG | 6806.20 | 6811.30 | ||
| 2Δ-RGMa | Sense CCAUCAUCUUCAAGAACUUCδCδ-p6 | 7219.40 | 7220.40 | |
| Antisense AAGUUCUUGAAGAUGAUGGUG | 6806.20 | 6811.30 | ||
| 4Δ-RGMa | Sense CCAUCAUCUUCAAGAACUUCδCδ-p | 7219.40 | 7220.40 | |
| Antisense AAGUUCUUGAAGAUGAUGGCδCδ-p | 7460.60 | 7466.90 | ||
| 2H-RGMa | Sense CCAUCAUCUUCAAGAACUUCηCη-p | 7260.00 | 7262.90 | |
| Antisense AAGUUCUUGAAGAUGAUGGUG | 6806.20 | 6811.30 | ||
| 4H-RGMa | Sense CCAUCAUCUUCAAGAACUUCηCη-p | 7260.00 | 7262.90 | |
| Antisense AAGUUCUUGAAGAUGAUGGUηGη-p | 7501.20 | 7502.00 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derzhalova, A.; Markov, O.; Fokina, A.; Shiohama, Y.; Zatsepin, T.; Fujii, M.; Zenkova, M.; Stetsenko, D. Novel Lipid-Oligonucleotide Conjugates Containing Long-Chain Sulfonyl Phosphoramidate Groups: Synthesis and Biological Properties. Appl. Sci. 2021, 11, 1174. https://doi.org/10.3390/app11031174
Derzhalova A, Markov O, Fokina A, Shiohama Y, Zatsepin T, Fujii M, Zenkova M, Stetsenko D. Novel Lipid-Oligonucleotide Conjugates Containing Long-Chain Sulfonyl Phosphoramidate Groups: Synthesis and Biological Properties. Applied Sciences. 2021; 11(3):1174. https://doi.org/10.3390/app11031174
Chicago/Turabian StyleDerzhalova, Alina, Oleg Markov, Alesya Fokina, Yasuo Shiohama, Timofei Zatsepin, Masayuki Fujii, Marina Zenkova, and Dmitry Stetsenko. 2021. "Novel Lipid-Oligonucleotide Conjugates Containing Long-Chain Sulfonyl Phosphoramidate Groups: Synthesis and Biological Properties" Applied Sciences 11, no. 3: 1174. https://doi.org/10.3390/app11031174