
The Triad Hsp60-miRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Histopathology
2.3. Tissue Processing and Immunomorphological Analysis
2.4. EV Isolation and Characterization
2.5. Western Blot
2.6. MicroRNAs Extraction and Real-Time PCR
2.7. Statistics
3. Results
3.1. Patients Follow-Up
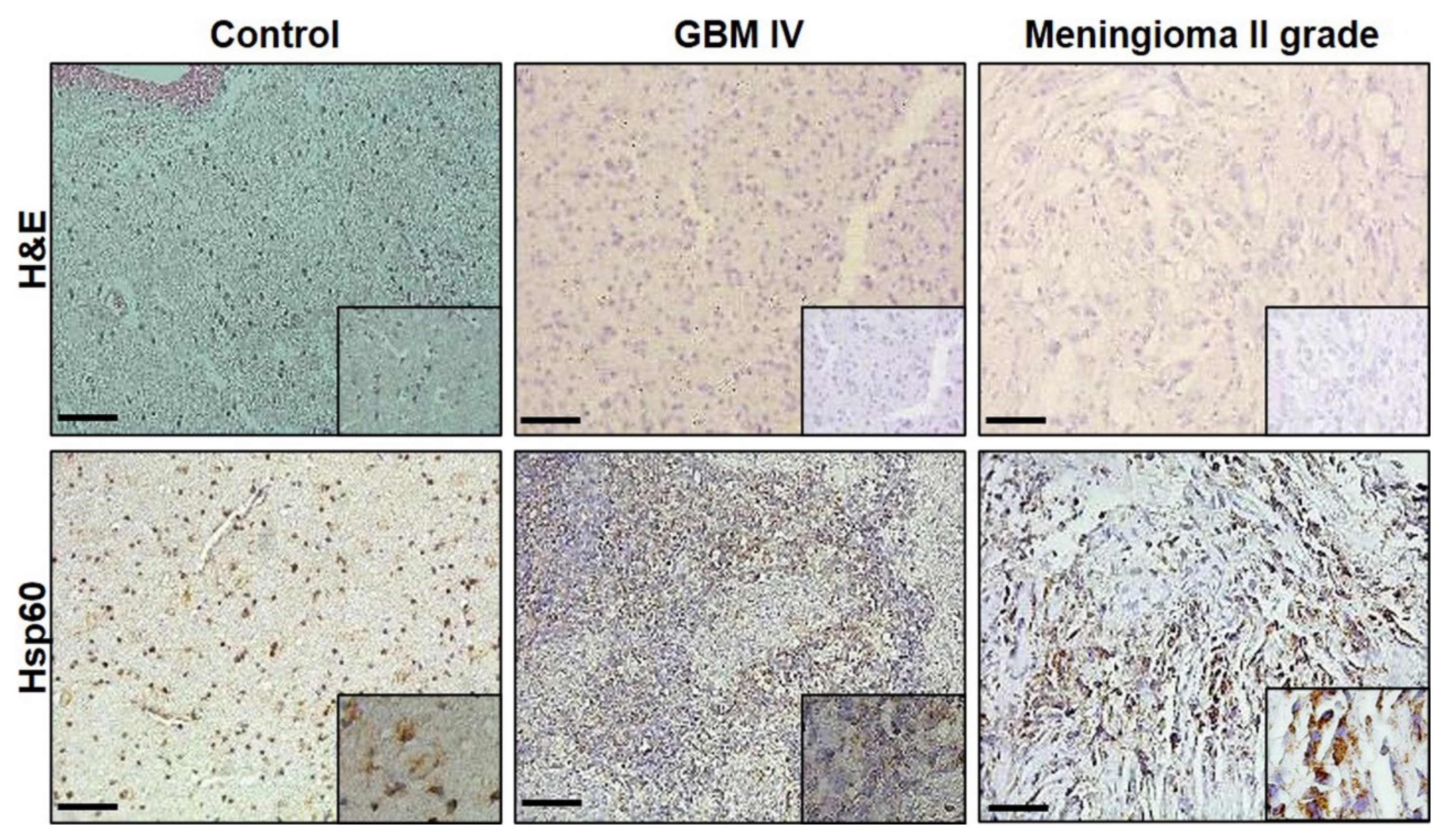
3.2. Immunomorphological Analysis
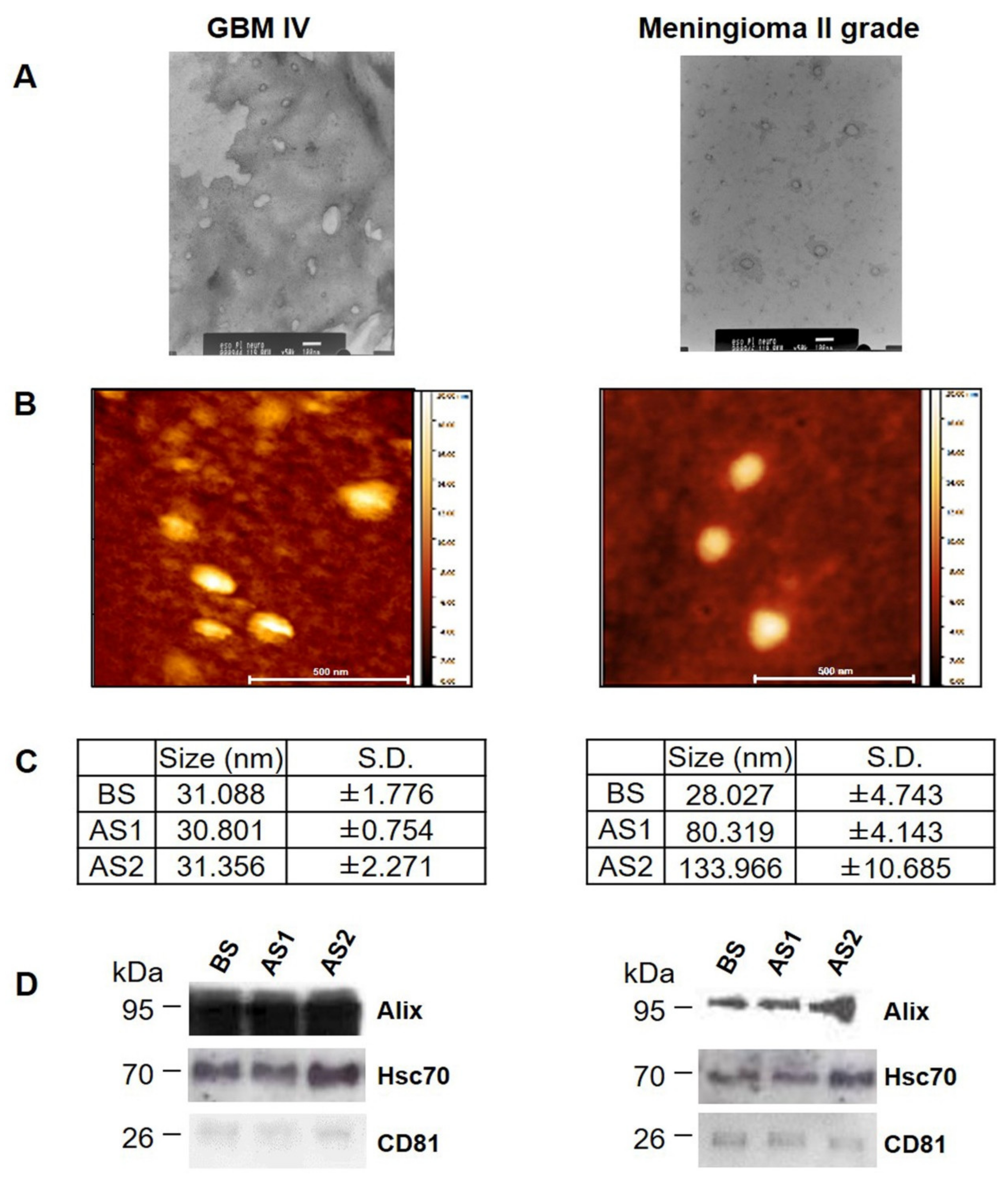
3.3. Characterization of EVs from GBM and Meningioma II Grade
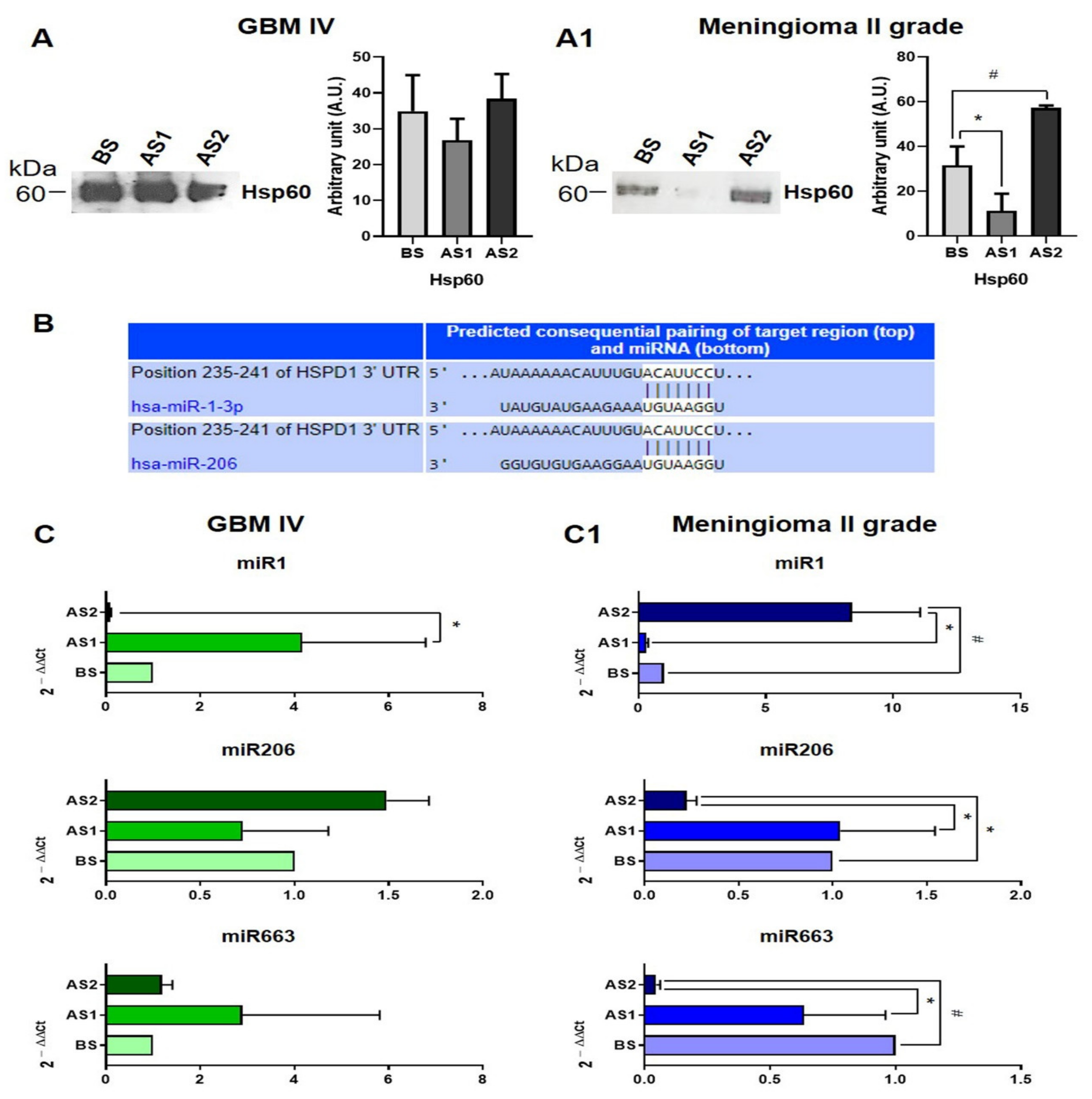
3.4. Hsp60 and Related miRNAs Levels in EVs
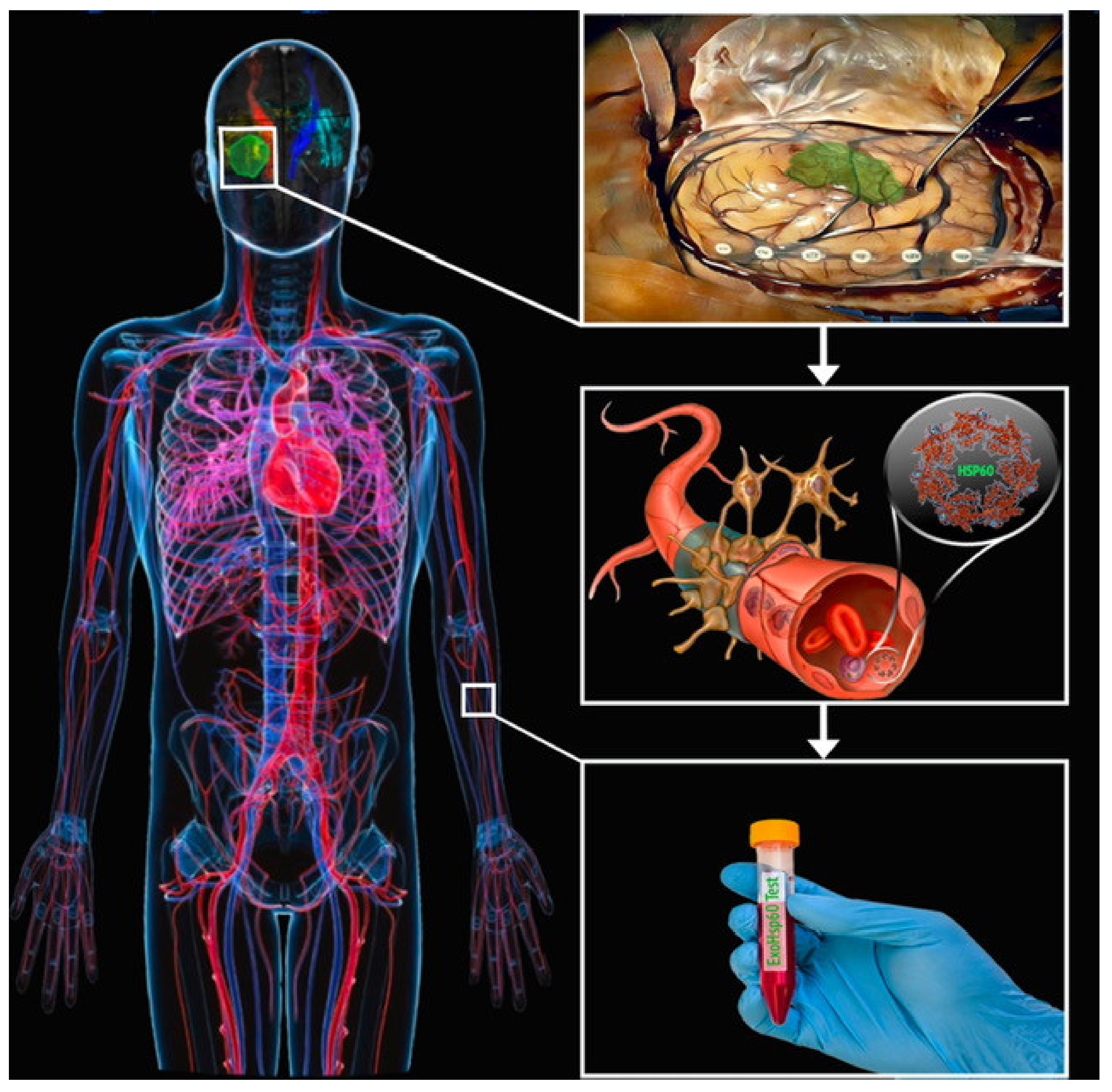
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Cioffi, G.; Gittleman, H.; Patil, N.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the united states in 2012–2016. Neurooncology 2019, 21, v1–v100. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA A Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Alifieris, C.; Trafalis, D.T. Glioblastoma multiforme: Pathogenesis and treatment. Pharmacol. Ther. 2015, 152, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Ashby, L.S.; Ryken, T.C. Management of malignant glioma: Steady progress with multimodal approaches. Neurosurg. Focus 2006, 20, E3. Available online: https://pubmed.ncbi.nlm.nih.gov/16709034/ (accessed on 4 January 2021). [CrossRef] [PubMed]
- Ryken, T.C.; Frankel, B.; Julien, T.; Olson, J.J. Surgical management of newly diagnosed glioblastoma in adults: Role of cytoreductive surgery. J. Neuro-Oncology 2008, 89, 271–286. [Google Scholar] [CrossRef]
- Brancato, V.; Nuzzo, S.; Tramontano, L.; Condorelli, G.; Salvatore, M.; Cavaliere, C. Predicting survival in glioblastoma patients using diffusion MR imaging metrics—A systematic review. Cancers 2020, 12, 2858. [Google Scholar] [CrossRef]
- Montemurro, N.; Anania, Y.; Cagnazzo, F.; Perrini, P. Survival outcomes in patients with recurrent glioblastoma treated with Laser Interstitial Thermal Therapy (LITT): A systematic review. Clin. Neurol. Neurosurg. 2020, 195, 105942. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Kondengaden, S.M.; Che, F.; Wang, L.; Heng, X. Potential epigenetic-based therapeutic targets for glioma. Front. Mol. Neurosci. 2018, 11, 408. [Google Scholar] [CrossRef]
- Liu, L.; Wang, G.; Wang, L.; Yu, C.; Li, M.; Song, S.; Hao, L.; Ma, L.; Zhang, Z. Computational identification and characterization of glioma candidate biomarkers through multi-omics integrative profiling. Biol. Direct 2020, 15, 1–14. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Chaperone proteins and chaperonopathies. In Handbook of Stress; Stress Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Elsevier/Academic Press: Cambridge, MA, USA, 2019; Volume 3, Chapter 12; pp. 135–152. [Google Scholar] [CrossRef]
- Kocaturk, N.M.; Gozuacik, D. Crosstalk between mammalian autophagy and the ubiquitin-proteasome system. Front. Cell Dev. Biol. 2018, 6, 128. [Google Scholar] [CrossRef]
- Tekirdag, K.; Cuervo, A.M. Chaperone-mediated autophagy and endosomal microautophagy: Jointed by a chaperone. J. Biol. Chem. 2018, 293, 5414–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macario, A.J.L.; Conway de Macario, E. Molecular mechanisms in chaperonopathies: Clues to understanding the histopathological abnormalities and developing novel therapies. J. Pathol. 2020, 250, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Rappa, F.; Unti, E.; Baiamonte, P.; Cappello, F.; Scibetta, N. Different immunohistochemical levels of Hsp60 and Hsp70 in a subset of brain tumors and putative role of Hsp60 in neuroepithelial tumorigenesis. Eur. J. of Histochem. 2013, 57, 20. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, G.A.; Vartholomatos, G.; Stefanaki, K.; Patereli, A.; Dova, L.; Karamoutsios, A.; Lallas, G.; Sfakianos, G.; Moschovi, M.; Prodromou, N. Expression of heat shock proteins in medulloblastoma. J. Neurosurgery Pediatr. 2013, 12, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Lo Cascio, F.; Mocciaro, E.; Vitale, A.M.; Vergilio, G.; Pace, A.; Cappello, F.; Campanella, C.; Palumbo Piccionello, A. Curcumin affects HSP60 folding activity and levels in neuroblastoma cells. Int. J. Mol. Sci. 2020, 21, 661. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, G.A.; Karamoutsios, A.; Lallas, G.; Ragos, V.; Goussia, A.; Kyritsis, A.P.; Voulgaris, S.; Vartholomatos, G. Expression of heat shock proteins in brain tumors. Turk. Neurosurg. 2014, 24, 745–749. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Li, J.; Liu, X.; Wang, G.; Luo, M.; Deng, H. Down-regulation of HSP60 suppresses the proliferation of glioblastoma cells via the ROS/AMPK/mTOR pathway. Sci. Rep. 2016, 6, 28388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Z.-X.; Lin, Q.-X.; Deng, C.-Y.; Zhu, J.-N.; Mai, L.-P.; Liu, J.-L.; Fu, Y.-H.; Liu, X.-Y.; Li, Y.-X.; Zhang, Y.-Y.; et al. miR-1/miR-206 regulate Hsp60 expression contributing to glucose-mediated apoptosis in cardiomyocytes. FEBS Lett. 2010, 584, 3592–3600. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Sun, X.; Ren, J.; Li, X.; Gao, X.; Lu, C.; Zhang, Y.; Sun, H.; Wang, Y.; Wang, H.; et al. miR-1 exacerbates cardiac ischemia-reperfusion injury in mouse models. PLoS ONE 2012, 7, e50515. [Google Scholar] [CrossRef]
- Neumann, E.; Brandenburger, T.; Santana-Varela, S.; Deenen, R.; Köhrer, K.; Bauer, I.; Hermanns, H.; Wood, J.N.; Zhao, J.; Werdehausen, R. MicroRNA-1-associated effects of neuron-specific brain-derived neurotrophic factor gene deletion in dorsal root ganglia. Mol. Cell. Neurosci. 2016, 75, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Choghaei, E.; Khamisipour, G.; Falahati, M.; Naeimi, B.; Mossahebi-Mohammadi, M.; Tahmasebi, R.; Hasanpour, M.; Shamsian, S.; Hashemi, Z.S. Knockdown of microRNA-29a changes the expression of heat shock proteins in breast carcinoma MCF-7 cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 23, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Chen, X.; Li, X.; Li, Z.; Diao, H.; Liu, L.; Zhang, J.; Ju, J.; Wen, L.; Liu, X.; et al. MicroRNA-1 downregulation induced by carvedilol protects cardiomyocytes against apoptosis by targeting heat shock protein 60. Mol. Med. Rep. 2019, 19, 3527–3536. [Google Scholar] [CrossRef]
- Wang, L.L.; Dong, J.J.; An, B.Z.; Liang, J.; Cai, K.R.; Jin, Z.S.; Jin, H.S.; Hu, J.P. HAS-MIR-17 increases the malignancy of gastric lymphoma by HSP60/TNFR2 pathway. J. Biol. Regul. Homeost. Agents 2020, 34, 1317–1324. [Google Scholar] [CrossRef]
- Cesarini, V.; Silvestris, D.A.; Tassinari, V.; Tomaselli, S.; Alon, S.; Eisenberg, E.; Locatelli, F.; Gallo, A. ADAR2/miR-589-3p axis controls glioblastoma cell migration/invasion. Nucleic Acids Res. 2018, 46, 2045–2059. [Google Scholar] [CrossRef]
- Banelli, B.; Forlani, A.; Allemanni, G.; Morabito, A.; Pistillo, M.P.; Romani, M. MicroRNA in glioblastoma: An overview. Int. J. Genom. 2017, 2017, 7639084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, N.; Kim, Y.-J.; Mueller, S.C.; Backes, C.; Werner, T.V.; Galata, V.; Sartorius, E.; Bohle, R.M.; Keller, A.; Meese, E. Posttranscriptional deregulation of signaling pathways in meningioma subtypes by differential expression of miRNAs. Neurooncology 2015, 17, 1250–1260. [Google Scholar] [CrossRef] [Green Version]
- Kol, A.; Lichtman, A.H.; Finberg, R.W.; Libby, P.; Kurt-Jones, E.A. Cutting Edge: Heat Shock Protein (HSP) 60 activates the innate immune response: CD14 is an essential receptor for HSP60 activation of mononuclear cells. J. Immunol. 2000, 164, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flohé, S.B.; Brüggemann, J.; Lendemans, S.; Nikulina, M.; Meierhoff, G.; Flohé, S.; Kolb, H. Human heat shock protein 60 induces maturation of dendritic cells versus a Th1-promoting phenotype. J. Immunol. 2003, 170, 2340–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Knowlton, A.A. HSP60 trafficking in adult cardiac myocytes: Role of the exosomal pathway. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H3052–H3056. [Google Scholar] [CrossRef] [Green Version]
- Hayoun, D.; Kapp, T.; Edri-Brami, M.; Ventura, T.; Cohen, M.; Avidan, A.; Lichtenstein, R.G. HSP60 is transported through the secretory pathway of 3-MCA-induced fibrosarcoma tumour cells and undergoes N-glycosylation. FEBS J. 2012, 279, 2083–2095. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Bucchieri, F.; Merendino, A.M.; Fucarino, A.; Burgio, G.; Corona, D.F.V.; Barbieri, G.; David, S.; Farina, F.; Zummo, G.; et al. The odyssey of Hsp60 from tumor cells to other destinations includes plasma membrane-associated stages and golgi and exosomal protein-trafficking modalities. PLoS ONE 2012, 7, e42008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz Junho, C.V.; Trentin-Sonoda, M.; Alvim, J.M.; Gaisler-Silva, F.; Carneiro-Ramos, M.S. Ca2+/Calmodulin-dependent kinase II delta B is essential for cardiomyocyte hypertrophy and complement gene expression after LPS and HSP60 stimulation in vitro. Braz. J. Med Biol. Res. 2019, 52, e8732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Liu, Y.; Wang, S.; Zhao, N.; Qin, J.; Li, Y.; Fan, C.; Shan, Z.; Teng, W. Circulating exosomes activate dendritic cells and induce unbalanced CD4+ t cell differentiation in Hashimoto thyroiditis. J. Clin. Endocrinol. Metab. 2019, 104, 4607–4618. [Google Scholar] [CrossRef] [PubMed]
- Zabegina, L.; Nazarova, I.; Knyazeva, M.; Nikiforova, N.; Slyusarenko, M.; Titov, S.; Vasilyev, D.; Sleptzov, I.; Malek, A. miRNA let-7 from TPO(+) extracellular vesicles is a potential marker for a differential diagnosis of follicular thyroid nodules. Cells 2020, 9, 1917. [Google Scholar] [CrossRef]
- Graziano, F.; Caruso Bavisotto, C.; Marino Gammazza, A.; Rappa, F.; Conway de Macario, E.; Macario, A.J.L.; Cappello, F.; Campanella, C.; Maugeri, R.; Iacopino, D.G. Chaperonology: The third eye on brain gliomas. Brain Sci. 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso Bavisotto, C.; Graziano, F.; Rappa, F.; Marino Gammazza, A.; Logozzi, M.; Fais, S.; Maugeri, R.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; et al. Exosomal chaperones and miRNAs in gliomagenesis: State-of-art and theranostics perspectives. Int. J. Mol. Sci. 2018, 19, 2626. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Caruso Bavisotto, C.; Marino Gammazza, A.; Nikolic, D.; Rappa, F.; David, S.; Cappello, F.; Bucchieri, F.; Fais, S. Exosomal heat shock proteins as new players in tumour cell-to-cell communication. J. Circ. Biomarkers 2014, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Cappello, F.; Macario, A.J.L.; Conway de Macario, E.; Logozzi, M.; Fais, S.; Campanella, C. Exosomal HSP60: A potentially useful biomarker for diagnosis, assessing prognosis, and monitoring response to treatment. Expert Rev. Mol. Diagn. 2017, 17, 815–822. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Rappa, F.; Fucarino, A.; Pitruzzella, A.; David, S.; Campanella, C. Exosomes: Can doctors still ignore their existence? EuroMediterr. Biomed. J. 2013, 8. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; Bent, M.J.V.D.; Taphoorn, M.J.B.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Marino Gammazza, A.; Pitruzzella, A.; et al. Immunomorphological pattern of molecular chaperones in normal and pathological thyroid tissues and circulating exosomes: Potential use in clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Rappa, F.; Sciumè, C.; Marino Gammazza, A.; Barone, R.; Bucchieri, F.; David, S.; Curcurù, G.; Caruso Bavisotto, C.; Pitruzzella, A.; et al. Heat shock protein 60 levels in tissue and circulating exosomes in human large bowel cancer before and after ablative surgery. Cancer 2015, 121, 3230–3239. [Google Scholar] [CrossRef] [PubMed]
- TargetScan. TargetScanHuman 7.2 Predicted Targeting of Human HSPD1. Available online: http://www.targetscan.org/cgi-bin/targetscan/vert_72/view_gene.cgi?rs=ENST00000388968.3&taxid=9606&members=&showcnc=0&shownc=0&showncf1=&showncf2=&subset=1#miR-1-3p/206 (accessed on 6 August 2020).
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, C.; Yu, S.-Z.; Liu, Q.; Rao, J.; Zhang, H.-R.; Xiao, H.-L.; Fu, T.-W.; Long, H.; He, Z.-C.; et al. miR-663 suppresses oncogenic function of CXCR4 in glioblastoma. Clin. Cancer Res. 2015, 21, 4004–4013. [Google Scholar] [CrossRef] [Green Version]
- Lammers, T.; Aime, S.; Hennink, W.E.; Storm, G.; Kiessling, F. Theranostic Nanomedicine. Acc. Chem. Res. 2011, 44, 1029–1038. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Guo, K.; Adkins, G.B.; Jiang, Q.; Liu, Y.; Sedano, S.; Duan, Y.; Yan, W.; Wang, S.E.; Bergersen, K.; et al. A Single Extracellular Vesicle (EV) flow cytometry approach to reveal EV heterogeneity. Angew. Chem. Int. Ed. 2018, 57, 15675–15680. [Google Scholar] [CrossRef]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular vesicle heterogeneity: Subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso Bavisotto, C.; Scalia, F.; Marino Gammazza, A.; Carlisi, D.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; Cappello, F.; Campanella, C. Extracellular vesicle-mediated cell–cell communication in the nervous system: Focus on neurological diseases. Int. J. Mol. Sci. 2019, 20, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Moita, C.; Van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; Manel, N.; Moita, L.F.; Théry, C.; Raposo, G. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 2013, 126, 5553–5565. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||||
| Pt a | Sex/Age (years) | Brain Site | Histopathology b | Follow-Up | |
| Molecular BS-AS1-AS2 | Clinical/Months | ||||
| 1 | M/65 | Left temporal-peritrigonal | HGG GBM IV WHO NOS: GFAP+, IDH+, Synaptofisin-; Ki67+ <10% | C | 14 |
| 2 | M/57 | Right frontal | HGG GBM IV WHO NOS: p53+, GFAP+, CAM 5.2-; Ki67 > 60% | C | 10 |
| 3 | M/60 | Left temporo-parietal | LGG II WHO: GFAP+, Synaptofisin+, Ki67+ < 3% | C | 12 |
| 4 | M/55 | Left fronto-temporal | HGG GBM IV WHO NOS: GFAP++, PanCK+, Ki67+ > 30% | C | 8 |
| 5 | F/65 | Right fronto-temporo-parietal | HGG GBM IV WHO IDH mutant: Framments HGG: GFAP+, IDH+, p53+ > 3%; Ki67+ > 30%: Framments Anaplastic Astrocitoma: Ki67+ < 5% | C | 9 |
| 6 | F/70 | Left fronto-temporal-peritrigonal | HGG III WHO NOS: High differentiated elements: GFAP +, S-100+, synaptofisin +, rounded cells. CD57+, vimentin+, GFAP-, NSE-, synaptofisin- with Ki67 15%, CAM 5.2 -, PanCK -, p53 -, factor XIII -, desmin -, CD68 -, CD45 - | I | 5;D |
| 7 | F/58 | Right frontal | HGG GBM IV NOS: GFAP+; S-100+; Synaptofisin-; NSE-; CD34-; Ki-67+: 30% | C | 4; D |
| 8 | M/60 | Left temporo-parietal | HGG GBM IV NOS: GFAP (+/-), S100 (+), p53 (+/-), synaptofisin -, CD57 -, CD99 -, Cam 5.2-, bcatenin -, CD45-, Ki67 30% | C | 12 |
| 9 | M/67 | Left frontal | HGG GBM IV NOS: GFAP+, Synaptofisin-, NSE+/-, Vimentin +/-; Ki-67+: 30% S-100+ | C | 10 |
| 10 | F/56 | Right temporo-parietal | HGG GBM IV IDH mutant: GFAP+, S100+, synaptofisin-, PanCK+, p53+, IDH+; Ki67 > 35% | C | 14 |
| 11 | M/68 | Left frontal | HGG GBM IV NOS: GFAP+, synaptofisin+, PanCK+, Ki67 30% | I | 5; R |
| 12 | F/58 | Left frontal | HGG GBM IV NOS: GFAP+++; synaptofisin -; Ki67 + 25% | I | 5 |
| 13 | M/63 | Right temporo-parietal | HGG GBM IV NOS: LARGE CELLS GFAP+, S100+, PanCK+, p53+/-, synaptofisin-, c-myc-, CD45-, CD20-, CD57-, NSE-, CD99+/-, neurofilaments-. SMALL CELLS GFAP-, S100 -/+, PanCK-, p53+/-, synaptofisin-, c-myc-, CD45-, CD20-, CD57-, NSE-/+, CD99+/-, neurofilaments- | C | 18 |
| 14 | M/72 | Right parietal | HGG GBM IV IDH wild type: GFAP+, synaptofisin-, Ki-67+ 20%. | C | 6; R |
| 15 | M/59 | Left frontal | HGG GBM IV IDH MUTANT: GFAP+, synaptofisin-, IDH +, CD34+, Ki67 30% | C | 10 |
| 16 | M/64 | Left fronto-parietal | HGG GBM IV NOS: GFAP+, CD34-, synaptofisin-, Ki67 30% | C | 8; R |
| (B) | |||||
| Pt a | Sex/Age (years) | Anatomical Site | Histopathological Classification b | Follow-Up | |
| Molecular BS-AS1-AS2 | Clinical /Months | ||||
| 1 | M/60 | Right temporal convexity | Angiomatous meningioma G I | C | 14 |
| 2 | F/55 | Right convexity frontal | Fibrous meningioma G I | C | 10 |
| 3 | M/60 | Left parietal convexity | Atypical meningioma G II | C | 12 |
| 4 | F/54 | Fronto-basal/Olfactory groove | Angiomatous meningioma G I | C | 8 |
| 5 | F/60 | Parafalcine parietal convexity | Transitional meningioma G I | C | 9 |
| 6 | F/70 | Temporal CPA | Meningothelial meningioma G I | C | 10 |
| 7 | M/60 | Right convexity frontal | Atypical meningioma G II | C | 12 |
| 8 | M/68 | Occipital/Tentorial | Transitional meningioma G I | C | 12 |
| 9 | F/62 | Left temporal convexity | Meningothelial meningioma G I | C | 10 |
| 10 | F/50 | Occipital-Foramen Magnum | Fibrous meningioma G I | C | 14 |
| 11 | M/72 | Left frontal convexity | Atypical meningioma G II | C | |
| 12 | F/65 | Right temporal-sphenoid wing | Atypical meningioma G II | C | 5 |
| 13 | M/68 | Fronto-basal/Planum | Transitional meningioma G I | C | 18 |
| 14 | M/71 | Parafalcine frontoparietal convexity | Atypical meningiomas G II | C | 12; D |
| 15 | F/59 | Skull base/olfactory groove | Atypical meningioma G II | C | 10 |
| 16 | M/67 | Parafalicine left parietal convexity | Atypical meningioma G II | C | 8 |
| 17 | F/48 | Temporal-CPA | Fibrous meningioma G I | C | 12 |
| 18 | M/69 | Right frontal convexity | Meningothelial meningioma G I | C | 10 |
| Name | Sequence |
|---|---|
| Hs_miR-1_2 | 5′-UGGAAUGUAAAGAAGUAUGUAU |
| Hs_miR-206_1 | 5′-UGGAAUGUAAGGAAGUGUGUGG |
| Hs_miR-663b_2 | 5′-GGUGGCCCGGCCGUGCCUGAGG |
| Hs_miR-16 | 5′-AGCAGCACGUAAAUAUUGGCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graziano, F.; Iacopino, D.G.; Cammarata, G.; Scalia, G.; Campanella, C.; Giannone, A.G.; Porcasi, R.; Florena, A.M.; Conway de Macario, E.; Macario, A.J.L.; et al. The Triad Hsp60-miRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients. Appl. Sci. 2021, 11, 2867. https://doi.org/10.3390/app11062867
Graziano F, Iacopino DG, Cammarata G, Scalia G, Campanella C, Giannone AG, Porcasi R, Florena AM, Conway de Macario E, Macario AJL, et al. The Triad Hsp60-miRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients. Applied Sciences. 2021; 11(6):2867. https://doi.org/10.3390/app11062867
Chicago/Turabian StyleGraziano, Francesca, Domenico Gerardo Iacopino, Giacomo Cammarata, Gianluca Scalia, Claudia Campanella, Antonino Giulio Giannone, Rossana Porcasi, Ada Maria Florena, Everly Conway de Macario, Alberto J.L. Macario, and et al. 2021. "The Triad Hsp60-miRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients" Applied Sciences 11, no. 6: 2867. https://doi.org/10.3390/app11062867
APA StyleGraziano, F., Iacopino, D. G., Cammarata, G., Scalia, G., Campanella, C., Giannone, A. G., Porcasi, R., Florena, A. M., Conway de Macario, E., Macario, A. J. L., Nicoletti, G. F., & Caruso Bavisotto, C. (2021). The Triad Hsp60-miRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients. Applied Sciences, 11(6), 2867. https://doi.org/10.3390/app11062867