
Microbial Depolymerization of Epoxy Resins: A Novel Approach to a Complex Challenge

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Epoxy Samples
2.2. Experimental Set-Up
2.2.1. Standard Soil (Microcosms)
2.2.2. Microbiological Water
2.2.3. Isolation of Microbes from Microbiological Water
2.2.4. Genetic Identification of Isolated Microbes
2.3. Analytical Techniques
2.3.1. Weight Loss and Drying Procedure
2.3.2. Hardness
2.3.3. Flexure Test
2.3.4. Colorimetry
2.3.5. Wettability (Contact Angle)
2.3.6. Fourier Transformed Infrared Spectroscopy (FTIR)
2.3.7. Environmental Scanning Electron Microscopy (ESEM)
2.3.8. Statistical Analysis
3. Results
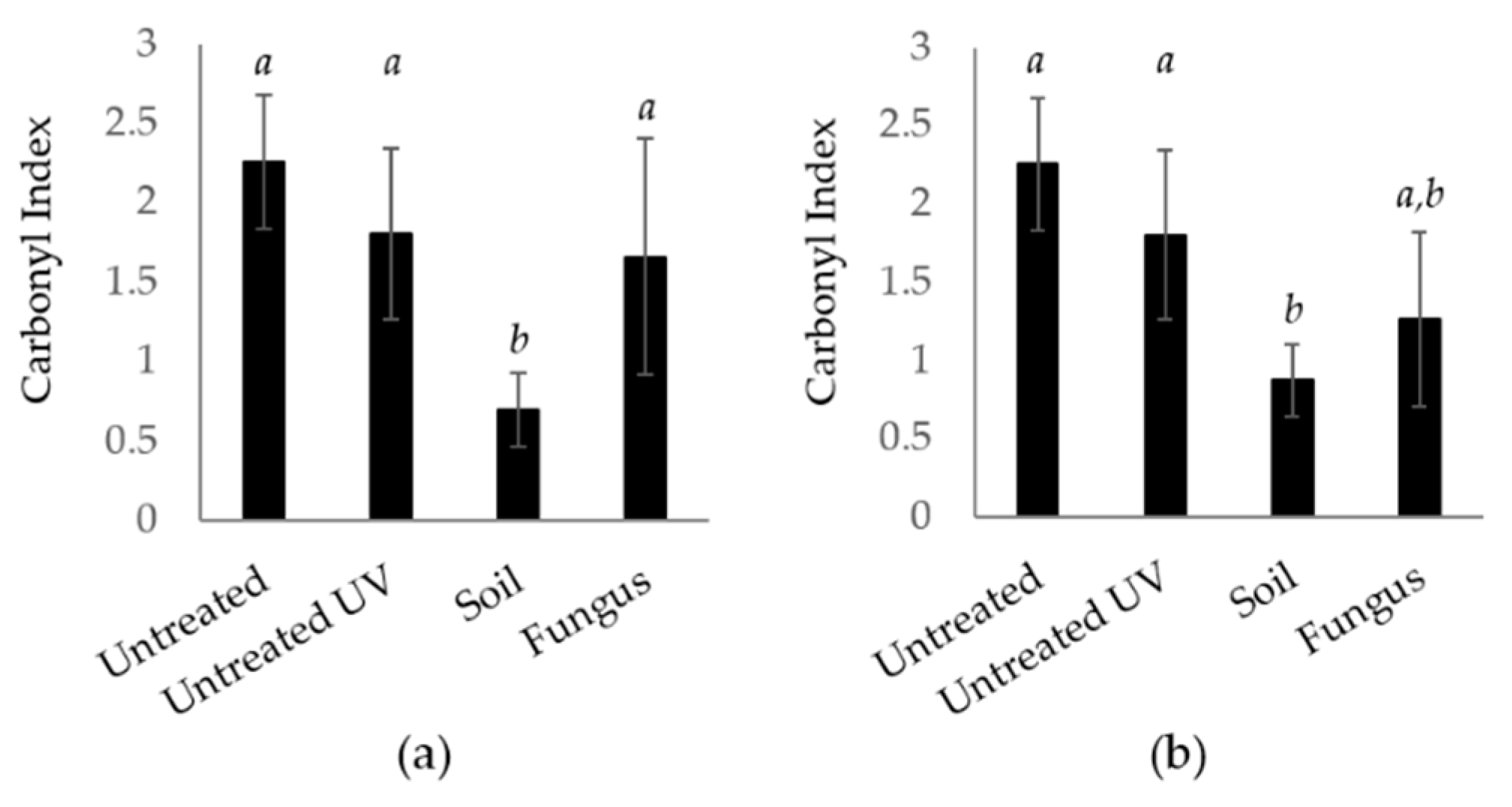
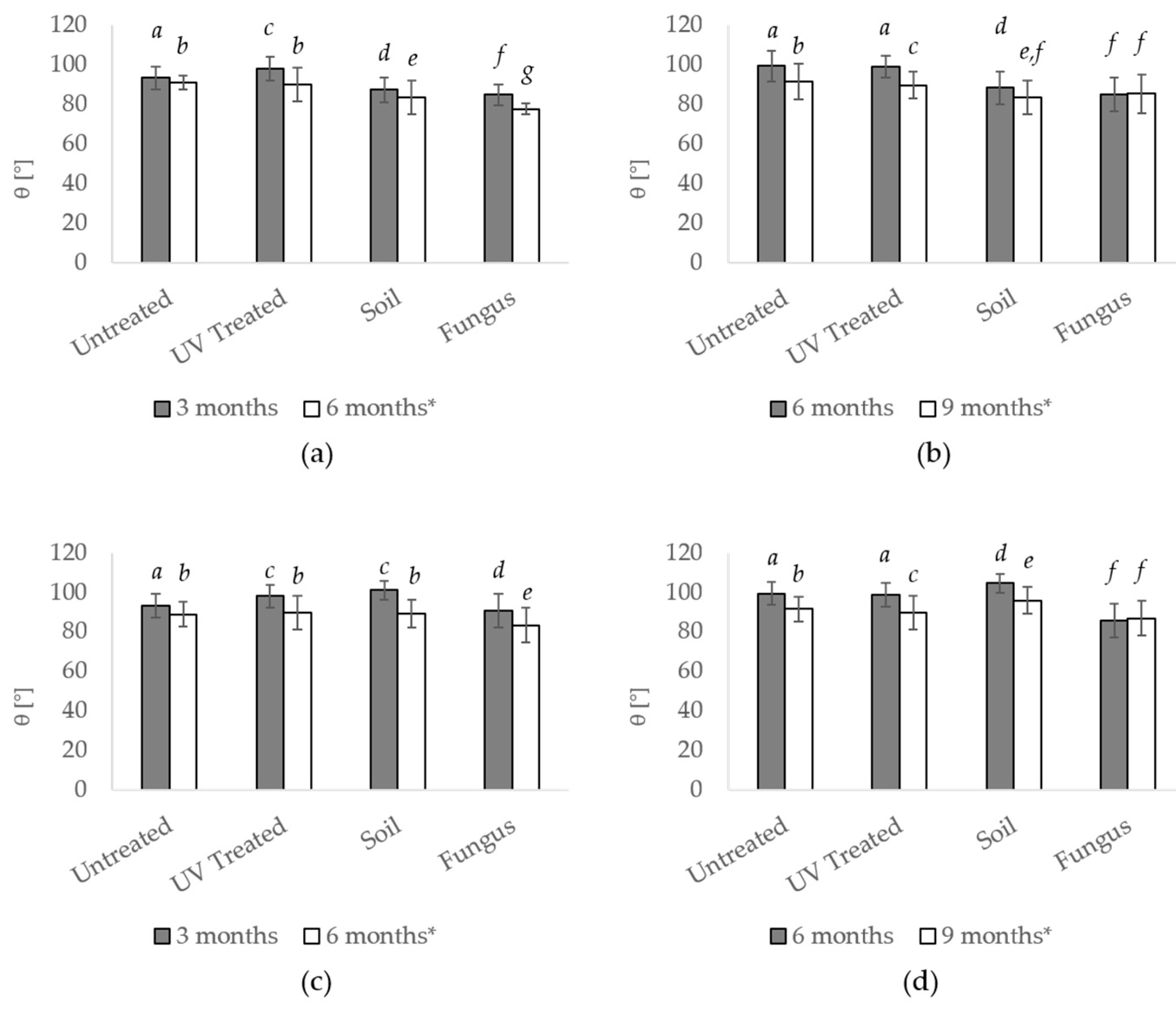
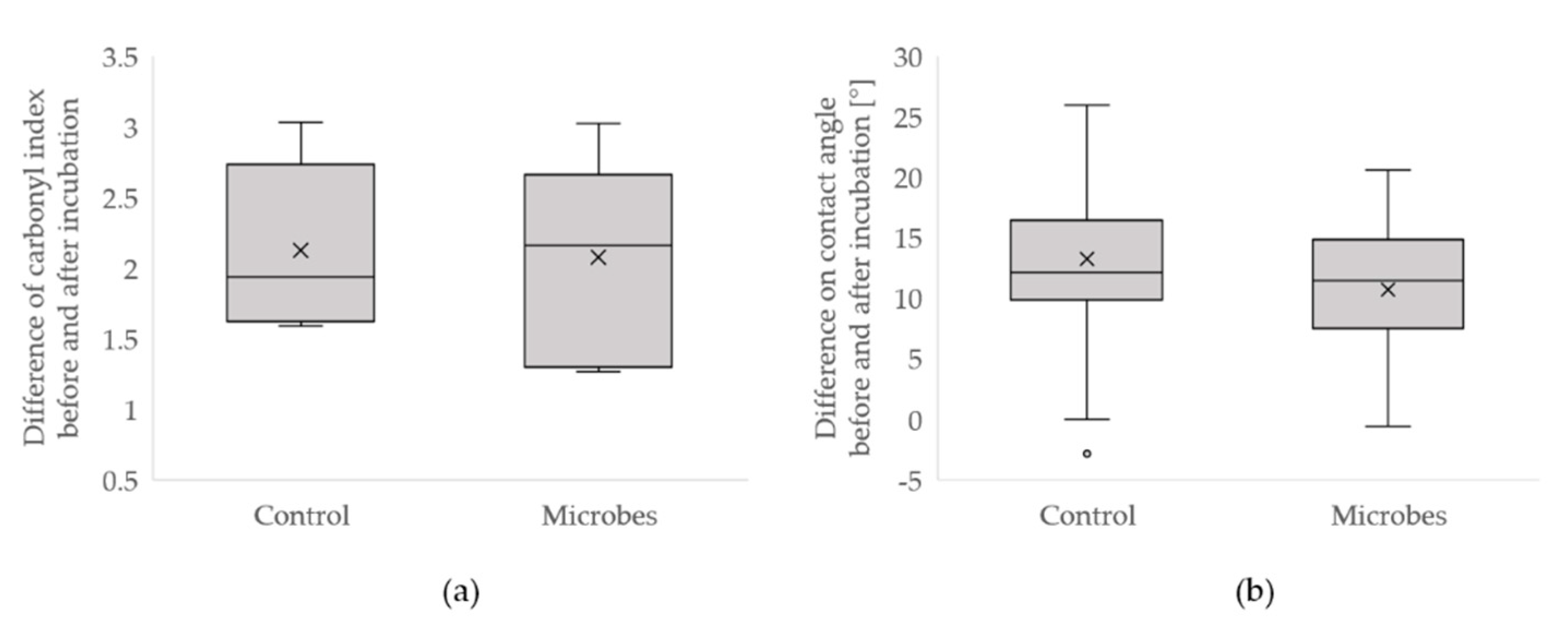
3.1. Standard Soil
3.2. Microbiological Water
3.2.1. Observation on Older Samples from Microbiological Water
3.2.2. Isolation and Identification of Microbes
3.2.3. New Samples Incubated for Three Months in the Microbiological Water
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Esposito, M.; Tse, T.; Soufani, K. Is the circular economy a new fast-expanding market? Thunderbird Int. Bus Rev. 2017, 59, 9–14. [Google Scholar] [CrossRef]
- Campbell, F.C. Structural Composite Materials; ASM International: Geauga County, OH, USA, 2010. [Google Scholar]
- Palucka, T.; Bensaude-Vincent, B. Composites Overview. 2002. Available online: http://authors.library.caltech.edu/5456/1/hrst.mit.edu/hrs/materials/public/composites/Composites_Overview.htm (accessed on 26 July 2017).
- Das, S.; Warren, J.; West, D.; Schexnayder, S.M. Global Carbon Fiber Composites Supply Chain Competitiveness Analysis; Oak Ridge National Laboratory: Oak Ridge, TN, USA; The University of Tennessee: Knoxville, TN, USA, 2016. [Google Scholar]
- Kunststoffe, A.V.K. Carbon Composites in Composites Market Report. 2016. Available online: https://www.carboncomposites.eu/media/2448/marktbericht_2016_ccev-avk (accessed on 26 July 2017).
- Teuten, E.L.; Saquing, J.M.; Knappe, D.R.; Barlaz, M.A.; Jonsson, S.; Björn, A. Transport and release of chemicals from plastics to the environment and to wildlife. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2027–2045. [Google Scholar] [CrossRef] [Green Version]
- Longana, M.L.; Yu, H.; Jalavand, M.; Wisnom, M.R.; Potter, K.D. Multiple closed loop recycling of carbon fibre composites with the HiPerDiF (High Performance Discontinuous Fibre) method. Compos. Struct. 2016, 153, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Longana, M.L.; Yu, H.; Jalavand, M.; Wisnom, M.R.; Potter, K.D. Aligned discontinuous intermingled reclaimed/virgin carbon fibre composites for high performance and pseudo-ductile behaviour in interlaminated carbon-glass hybrids. Compos. Sci. Technol. 2017, 143, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Feraboli, P.; Kawakami, H.; Wade, B.; Gasco, F.; DeOto, L.; Masini, A. Recyclability and reutilization of carbon fiber fabric/epoxy composites. J. Compos. Mater. 2012, 46, 1459–1473. [Google Scholar] [CrossRef]
- Pimenta, S.; Pinho, S.T. The effect of recycling on the mechanical response of carbon fibres and their composites. Compos. Struct. 2012, 94, 3669–3684. [Google Scholar] [CrossRef] [Green Version]
- Materials, K.K.T.N. Composite Recycling. 2010, pp. 1–26. Available online: https://compositesuk.co.uk/system/ (accessed on 26 July 2017).
- Tiso, T.; Narancic, T.; Wei, R.; Pollet, E.; Beagan, N.; Schröder, K.; Honak, A.; Jiang, M.; Shane, T.K.; Kenny, T.S.; et al. Towards bio-upcycling of polyethylene terephthalate. Metab. Eng. 2021, 66, 167–178. [Google Scholar] [CrossRef]
- Wagner, P.; Ray, R.; Hart, K.; Little, B.; Bradley, W. Microbial Degradation at Stressed Fiber Reinforced Polymeric Composites; Hitachi LDT Kanagawa (Japan) Production Engineering Research Lab: Tokyo, Japan, 1995. [Google Scholar]
- Cappitelli, F.; Principi, P.; Pedrazzani, R.; Toniolo, L.; Sorlini, C. Bacterial and fungal deterioration of the Milan Cathedral marble treated with protective synthetic resins. Sci. Total Environ. 2007, 385, 172–181. [Google Scholar] [CrossRef]
- Gu, J.-D. Microbiological deterioration and degradation of synthetic polymeric materials: Recent research advances. Int. Biodeterior. Biodegrad. 2003, 52, 69–91. [Google Scholar] [CrossRef]
- Wang, G.; Chai, K.; Wu, J.; Liu, F. Effect of Pseudomonas putida on the degradation of epoxy resin varnish coating in seawater. Int. Biodeterior. Biodegrad. 2016, 115, 156–163. [Google Scholar] [CrossRef]
- Gu, J.-D.; Ford, T.; Thorp, K.; Mitchell, R. Microbial growth on fiber reinforced composite materials. Int. Biodeter. 1996, 37, 197–204. [Google Scholar] [CrossRef]
- Pangallo, D.; Bučková, M.; Kraková, L.; Puškárová, A.; Šaková, N.; Grivalský, T.; Chovanová, K.; Zemánková, M. Biodeterioration of epoxy resin: A microbial survey through culture-independent and culture-dependent approaches. Environ. Microbiol. 2015, 17, 462–479. [Google Scholar] [CrossRef]
- Eliaz, N.; Ron, E.Z.; Gozin, M.; Younger, S.; Biran, D.; Tal, N. Microbial degradation of epoxy. Materials 2018, 11, 2123. [Google Scholar] [CrossRef] [Green Version]
- Cameron, M.D.; Timofeevski, S.; Aust, S.D. Enzymology of Phanerochaete chrysosporium with respect to the degradation of recalcitrant compounds and xenobiotics. Appl. Microbiol. Biotechnol. 2000, 54, 751–758. [Google Scholar] [CrossRef]
- Wong, D.W.S. Structure and Action Mechanism of Ligninolytic Enzymes. Appl. Biochem. Biotechnol. 2008, 157, 174–209. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Eldridge, J.D.; Hamonts, K.; Singh, B.K. Ant colonies promote the diversity of soil microbial communities. ISME J. 2019, 13, 1114–1118. [Google Scholar] [CrossRef]
- Ginzburg, O.; Whitford, W.G.; Steinberger, Y. Effects of harvester ant (Messor spp.) activity on soil properties and microbial communities in a Negev Desert ecosystem. Biol. Fertil. 2008, 45, 165–173. [Google Scholar] [CrossRef]
- Brosius, J.; Dull, T.J.; Sleeter, D.D.; Noller, H.F. Gene organization and primary structure of a ribosomal RNA operon from Escherichia coli. J. Mol. Biol. 1981, 148, 107–127. [Google Scholar] [CrossRef]
- Alijani Mamaghani, N.; Javan-Nikkhah, M.S.H.; De Respinis, S.; Pianta, E.; Tonolla, M. Endophytic Cephalotrichum spp. from Solanum tuberosum (potato) in Iran—A polyphasic analysis. Sydowia. Submitted.
- Ausubel, F.M.; Kingston, R.E.B.R.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology; Greene Publishing Associated & Wiley Interscience: New York, NY, USA, 1989. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar]
- Hall, B.G. Building Phylogenetic Trees from Molecular Data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- ASTM D1557-12e1; Standard Test Methods for Laboratory Compaction Characteristics of Soil Using Modified Effort (56,000 ft-lbf/ft3 (2700 kN-m/m3)). ASTM International: West Conshohocken, PA, USA, 2012. Available online: https://www.astm.org/ (accessed on 23 April 2021).
- Domagała, I.; Gil, L.; Firlej, M.; Pieniak, D.; Selech, J.; Romek, D.; Biedziak, B. Statistical Comparison of the Hardness and Scratch-Resistance of the PMMA Polymers Used in Orthodontic Appliances. Adv. Sci. Technol. Eng. Syst. J. 2020, 14, 250–261. [Google Scholar] [CrossRef]
- ISO Standard DIN EN ISO 14125 A 2013 (Fibre-Reinforced Plastic Composites—Determination of Flexural Properties). Available online: https://www.iso.org/standard/23637.html (accessed on 21 July 2021).
- Reddy, M.S. Biomineralization of calcium carbonates and their engineered applications: A review. Front Microbiol. 2013, 4, 314. [Google Scholar]
- Songulashvili, G.; Elisashvili, V.; Wasser, S.; Nevo, E.; Hadar, Y. Laccase and manganese peroxidase activities of Phellinus robustus and Ganoderma adspersum grown on food industry wastes in submerged fermentation. Biotechnol. Lett. 2006, 28, 1425–1429. [Google Scholar] [CrossRef]
- Sitarz, A.K.; Mikkelsen, J.D.; Højrup, P.; Meyer, A.S. Identification of a laccase from Ganoderma lucidum CBS 229.93 having potential for enhancing cellulase catalyzed lignocellulose degradation. Enzyme Microb. Technol. 2013, 53, 378–385. [Google Scholar] [CrossRef]
- Monger, H.C.; Curtis, H.; Daugherty Lindemann, L.A.; Liddell, C.W.; Craig, M. Microbial precipitation of pedogenic calcite. Geology 1991, 19, 997–1000. [Google Scholar] [CrossRef]
- Banks, E.D.; Taylor, M.N.; Gulley, J.; Lubbers, B.R.; Giarrizzo, J.G.; Bullen, H.A.; Hoehler, T.M.; Barton, A.H. Bacterial calcium carbonate precipitation in cave environments: A function of calcium homeostasis. Geomicrobiol. J. 2010, 27, 444–454. [Google Scholar] [CrossRef]
- Al-Thawadi, S.M. Ureolytic bacteria and calcium carbonate formation as a mechanism of strength enhancement of sand. J. Adv. Sci. Eng. Res. 2011, 1, 98–114. [Google Scholar]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Biomineralization of calcium carbonate polymorphs by the bacterial strains isolated from calcareous sites. J. Microbiol. Biotechnol. 2013, 23, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Guggiari, M.; Bloque, R.; Aragno, M.; Verrecchia, E.; Job, D.; Junier, P. Experimental calcium-oxalate crystal production and dissolution by selected wood-rot fungi. Int. Biodeter. 2011, 65, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Gadd, G.M.; Bahri-Esfahani, J.; Li, Q.; Rhee, J.G.; Wei, Z.; Fomina, M.; Liang, X. Oxalate production by fungi: Significance in geomycology, biodeterioration and bioremediation. Fungal Biol. Rev. 2014, 28, 36–55. [Google Scholar] [CrossRef]
- Simon, A. Highways and Subways: A Story of Fungi and Bacteria in Soils. Ph.D. Thesis, University of Neuchatel, Neuchâtel, Switzerland, 2016. [Google Scholar]
- Hervé, V.; Junier, T.; Bindschedler, S.; Verrecchia, E.; Junier, P. Diversity and ecology of oxalotrophic bacteria. World J. Microbiol. Biotechnol. 2016, 32, 28. [Google Scholar] [CrossRef]
- Mäkelä, M.; Galkin, S.; Hatakka, A.; Lundell, T. Production of organic acids and oxalate decarboxylase in lignin-degrading white rot fungi. Enzyme Microb. Technol. 2002, 30, 542–549. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Kidd, P.; Kuffner, M.; Prieto-Fernández, Á.; Hann, S.; Monterroso, C.; Sessitsch, A.; Wenzel, W.; Puschenreiter, M. Bacterially Induced Weathering of Ultramafic Rock and Its Implications for Phytoextraction. Appl. Environ. Microbiol. 2013, 79, 5094–5103. [Google Scholar] [CrossRef] [Green Version]
- Arvieu, J.-C.; Leprince, F.; Plassard, C. Release of oxalate and protons by ectomycorrhizal fungi in response to P-deficiency and calcium carbonate in nutrient solution. Ann. For. Sci. 2003, 60, 815–821. [Google Scholar] [CrossRef]
- Hamel, R.; Levasseur, R.; Appanna, V.D. Oxalic acid production and aluminum tolerance in Pseudomonas fluorescens. J. Inorg. Biochem. 1999, 76, 99–104. [Google Scholar] [CrossRef]
- Fomina, M.; Hillier, S.; Charnock, J.M.; Melville, K.; Alexander, I.J.; Gadd, G.M. Role of Oxalic Acid Overexcretion in Transformations of Toxic Metal Minerals by Beauveria caledonica. Appl. Environ. Microbiol. 2005, 71, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Nakata, P.A. The oxalic acid biosynthetic activity of Burkholderia mallei is encoded by a single locus. Microbiol. Res. 2011, 166, 531–538. [Google Scholar] [CrossRef]
- Van Kan, J.A.; Shaw, M.W.; Grant-Downton, R.T. Botrytis species: Relentless necrotrophic thugs or endophytes gone rogue? Mol. Plant. Pathol. 2014, 15, 957–961. [Google Scholar] [CrossRef]
- Bolaños, B.T.; Hernández, H.G.; Mejía, E.Z.; García, P.S.; Aguilera, G.M.; Díaz, C.N.; Laborde, J.I.D.R.; Cortes, R.R. Colonización de Trichoderma y Bacillus en Plántulas de Agave tequilana Weber, var. Azul y el Efecto Sobre la Fisiología de la Planta y Densidad de Fusarium. Rev. Mex. Fitopatol. 2014, 32, 62–74. [Google Scholar]
- Marzano, M.; Gallo, A.; Altomare, C. Improvement of biocontrol efficacy of Trichoderma harzianum vs. Fusarium oxysporum f. sp. lycopersici through UV-induced tolerance to fusaric acid. Biol. Control. 2013, 67, 397–408. [Google Scholar]
- Saravanakumar, K.; Yu, C.; Dou, K.; Wang, M.; Li, Y.; Chen, J. Biodiversity of Trichoderma community in the tidal flats and wetland of southeastern China. PLoS ONE 2016, 11, e0168020. [Google Scholar] [CrossRef]
- Greco, G.; Cecchi, G.; Di Piazza, S.; Cutroneo, L.; Capello, M.; Zotti, M. Fungal characterisation of a contaminated marine environment: The case of the Port of Genoa (North-Western Italy). Webbia 2018, 73, 97–106. [Google Scholar] [CrossRef]
- Vacondio, B.; Birolli, W.G.; Ferreira, I.M.; Seleghim, M.H.R.; Gonçalves, S.; Vasconcellos, S.P.; Porto, A.L.M. Biodegradation of pentachlorophenol by marine-derived fungus Trichoderma harzianum CBMAI 1677 isolated from ascidian Didemnun ligulum. Biocatal. Agric. Biotechnol. 2015, 4, 266–275. [Google Scholar] [CrossRef]
- Ezzi, M.I.; Lynch, J.M. Biodegradation of cyanide by Trichoderma spp. and Fusarium spp. Enzym. Microb. Technol. 2005, 36, 849–854. [Google Scholar] [CrossRef]
- Baarschers, W.H.; Heitland, H.S. Biodegradation of fenitrothion and fenitrooxon by the fungus Trichoderma viride. J. Agric. Food Chem. 1986, 34, 707–709. [Google Scholar] [CrossRef]
- Zafra, G.; Cortés-Espinosa, D.V. Biodegradation of polycyclic aromatic hydrocarbons by Trichoderma species: A mini review. Environ. Sci. Pollut. Res. 2015, 22, 19426–19433. [Google Scholar] [CrossRef]
- Lipsa, R.; Tudorachi, N.; Darie-Nita, R.N.; Oprică, L.; Vasile, C.; Chiriac, A. Biodegradation of poly (lactic acid) and some of its based systems with Trichoderma viride. Int. J. Biol. Macromol. 2016, 88, 515–526. [Google Scholar] [CrossRef]
- Wang, L. Four new records of Aspergillus sect. Usti from Shandong Province, China. Mycotaxon 2012, 120, 373–384. [Google Scholar] [CrossRef]
- De Carvalho, C.R.; Vieira, M.d.L.A.; Cantrell, C.L.; Wedge, D.E.; Alves, T.M.A.; Zani, C.L.; Pimenta, R.S.; Junior, P.A.S.; Murta, S.M.F.; Romanha, A.J.; et al. Biological activities of ophiobolin K and 6-epi-ophiobolin K produced by the endophytic fungus Aspergillus calidoustus. Nat. Prod. Res. 2016, 30, 478–481. [Google Scholar] [CrossRef]
- Samson, R.A. Ecology and general characteristics of indoor fungi. In Fundamentals of Mold Growth in Indoor Environments and Strategies for Healthy Living; Springer: Berlin/Heidelberg, Germany, 2011; pp. 101–116. [Google Scholar]
- Slack, G.J.; Puniani, E.; Frisvad, J.C.; Samson, R.A.; Miller, J.D. Secondary metabolites from Eurotium species, Aspergillus calidoustus and A. insuetus common in Canadian homes with a review of their chemistry and biological activities. Mycol. Res. 2009, 113, 480–490. [Google Scholar] [CrossRef]
- Antipova, T.V.; Zhelifonova, V.P.; Zaitsev, K.V.; Nedorezova, P.M.; Aladyshev, A.M.; Klyamkina, A.N.; Kostyuk, S.V.; Danilogorskaya, A.A.; Kozlovsky, A.G. Biodegradation of Poly-ε-caprolactones and Poly-l-lactides by Fungi. J. Polym. Environ. 2018, 26, 4350–4359. [Google Scholar] [CrossRef]
- Ghorbannezhad, H.; Moghimi, H.; Dastgheib, S.M.M. Biodegradation of high-molecular-weight aliphatic and aromatic hydrocarbons by Aspergillus calidoustus. J. Microb. World 2018, 4, 346–359. [Google Scholar]
- Sanguin, H.; Wilson, N.L.; Kertesz, M.A. Assessment of functional diversity and structure of phytate-hydrolysing bacterial community in Lolium perenne rhizosphere. Plant Soil 2016, 401, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.-K.; Kim, K.-D.; Ahn, K.-M.; Shin, D.-H.; Hwang, J.-H.; Seong, C.N.; Ka, J.O. Genetic and Phenotypic Diversity of Parathion-Degrading Bacteria Isolated from Rice Paddy Soils. J. Microbiol. Biotechnol. 2009, 19, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.-C.; E, G.; Wang, J.; Wang, N.; Chen, X.; Mu, Y.; Li, H.; Yang, Y.; Liu, Y.; Wang, Y. Functions and Unique Diversity of Genes and Microorganisms Involved in Arsenite Oxidation from the Tailings of a Realgar Mine. Appl. Environ. Microbiol. 2016, 82, 7019–7029. [Google Scholar] [CrossRef] [Green Version]
- González, A.J.; Fortunato, M.S.; Papalia, M.; Radice, M.; Gutkind, G.; Magdaleno, A.; Gallego, A.; Korol, S.E. Selection and identification of a bacterial community able to degrade and detoxify m-nitrophenol in continuous biofilm reactors. Ecotoxicol. Environ. Saf. 2015, 122, 245–251. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.; De Marco, P.; Bowerman, S.; Pacheco, C.; Lara, J.C.; Lidstrom, M.E.; Chistoserdova, L. Methyloversatilis universalis gen. nov., sp. nov., a novel taxon within the Betaproteobacteria represented by three methylotrophic isolates. Int. J. Syst. Evol. Microbiol. 2006, 56, 2517–2522. [Google Scholar] [CrossRef] [Green Version]
- Smalley, N.E.; Taipale, S.J.; De Marco, P.; Doronina, N.V.; Kyrpides, N.; Shapiro, N.; Woyke, T.; Kalyuzhnaya, M. Functional and genomic diversity of methylotrophic Rhodocyclaceae: Description of Methyloversatilis discipulorum sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 2227–2233. [Google Scholar] [CrossRef]
- Doronina, N.V.; Kaparullina, E.; Trotsenko, Y.A. Methyloversatilis thermotolerans sp. nov., a novel thermotolerant facultative methylotroph isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2014, 64, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zheng, S.; Zhang, H.; Duan, S. Autotrophic and heterotrophic nitrification-anoxic denitrification dominated the anoxic/oxic sewage treatment process during optimization for higher loading rate and energy savings. Bioresour. Technol. 2018, 263, 84–93. [Google Scholar] [CrossRef]
- Cai, T.; Qian, L.; Cai, S.; Chen, L. Biodegradation of Benazolin-Ethyl by Strain Methyloversatilis sp. cd-1 Isolated from Activated Sludge. Curr. Microbiol. 2010, 62, 570–577. [Google Scholar] [CrossRef]
 —addition of 10 mL of fungal suspension.
—addition of 10 mL of fungal suspension.
—addition of 10 mL of fungal suspension.
—addition of 10 mL of fungal suspension.







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardi-Comensoli, L.; Tonolla, M.; Colpo, A.; Palczewska, Z.; Revikrishnan, S.; Heeb, M.; Brunner, I.; Barbezat, M. Microbial Depolymerization of Epoxy Resins: A Novel Approach to a Complex Challenge. Appl. Sci. 2022, 12, 466. https://doi.org/10.3390/app12010466
Pardi-Comensoli L, Tonolla M, Colpo A, Palczewska Z, Revikrishnan S, Heeb M, Brunner I, Barbezat M. Microbial Depolymerization of Epoxy Resins: A Novel Approach to a Complex Challenge. Applied Sciences. 2022; 12(1):466. https://doi.org/10.3390/app12010466
Chicago/Turabian StylePardi-Comensoli, Lucrezia, Mauro Tonolla, Andrea Colpo, Zuzanna Palczewska, Sharanya Revikrishnan, Markus Heeb, Ivano Brunner, and Michel Barbezat. 2022. "Microbial Depolymerization of Epoxy Resins: A Novel Approach to a Complex Challenge" Applied Sciences 12, no. 1: 466. https://doi.org/10.3390/app12010466