
Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators



Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. ASE Materials and Extraction
2.3. Cell Culture
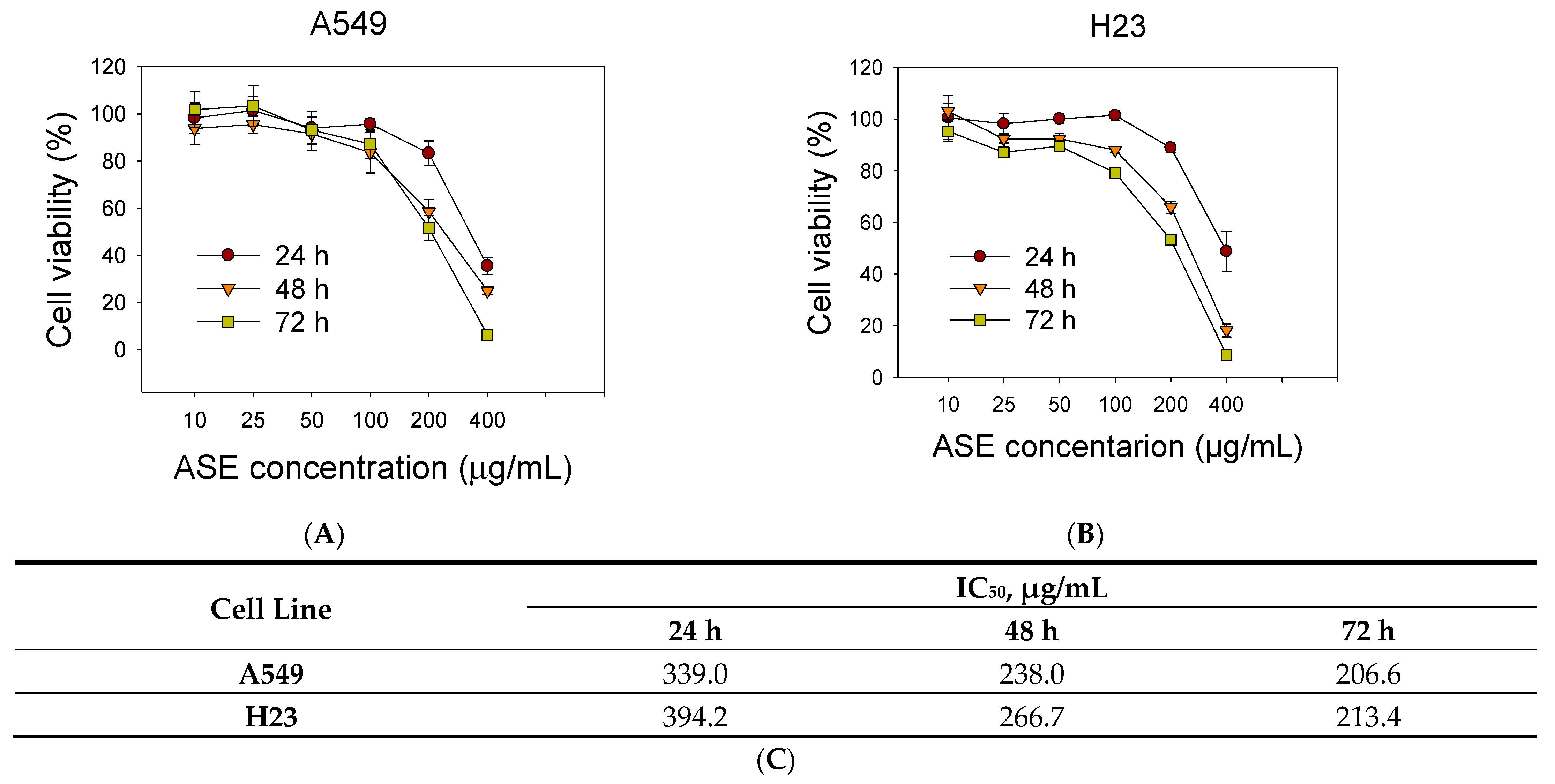
2.4. Cell Cytotoxicity Assay
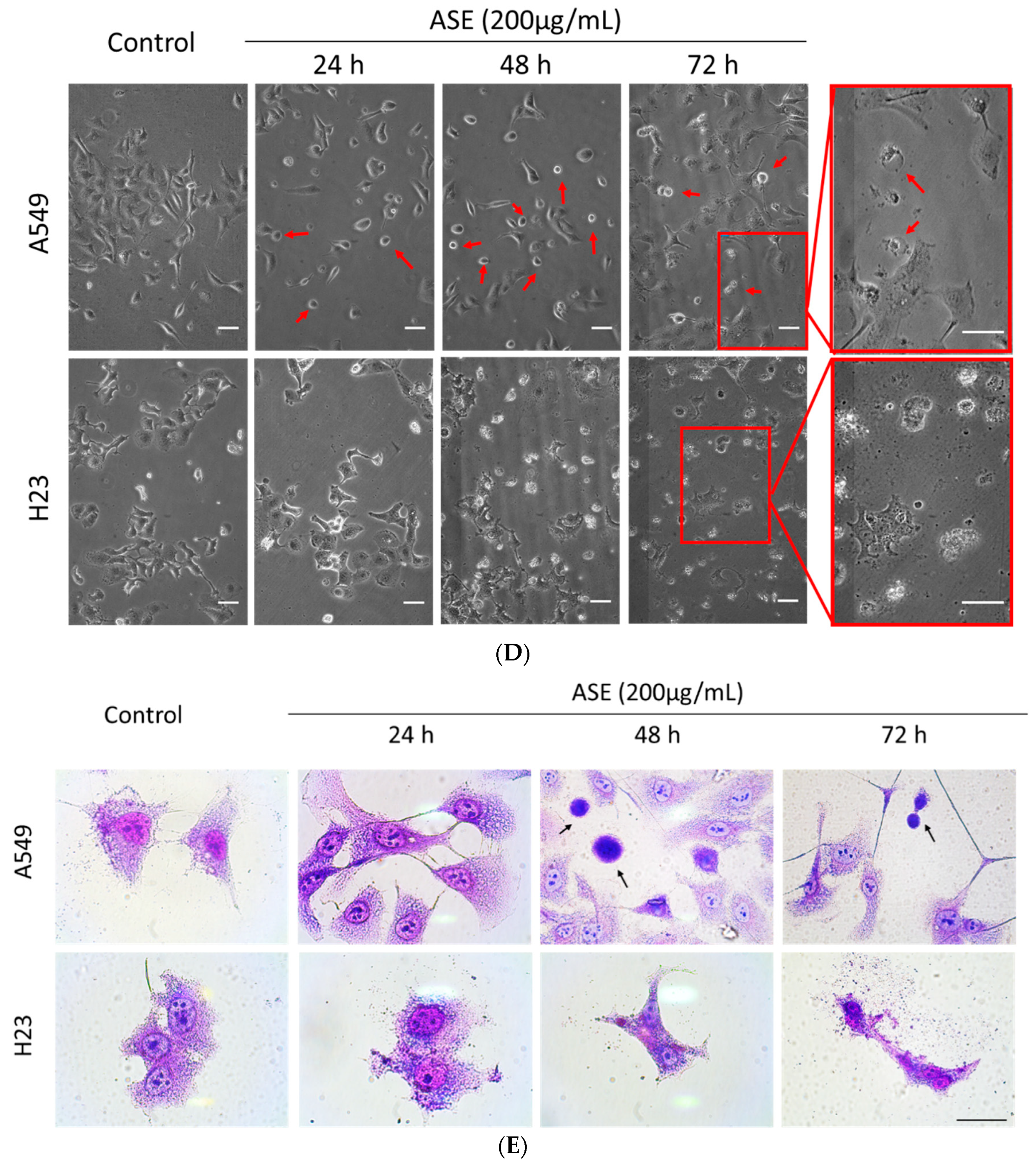
2.5. Cell Staining
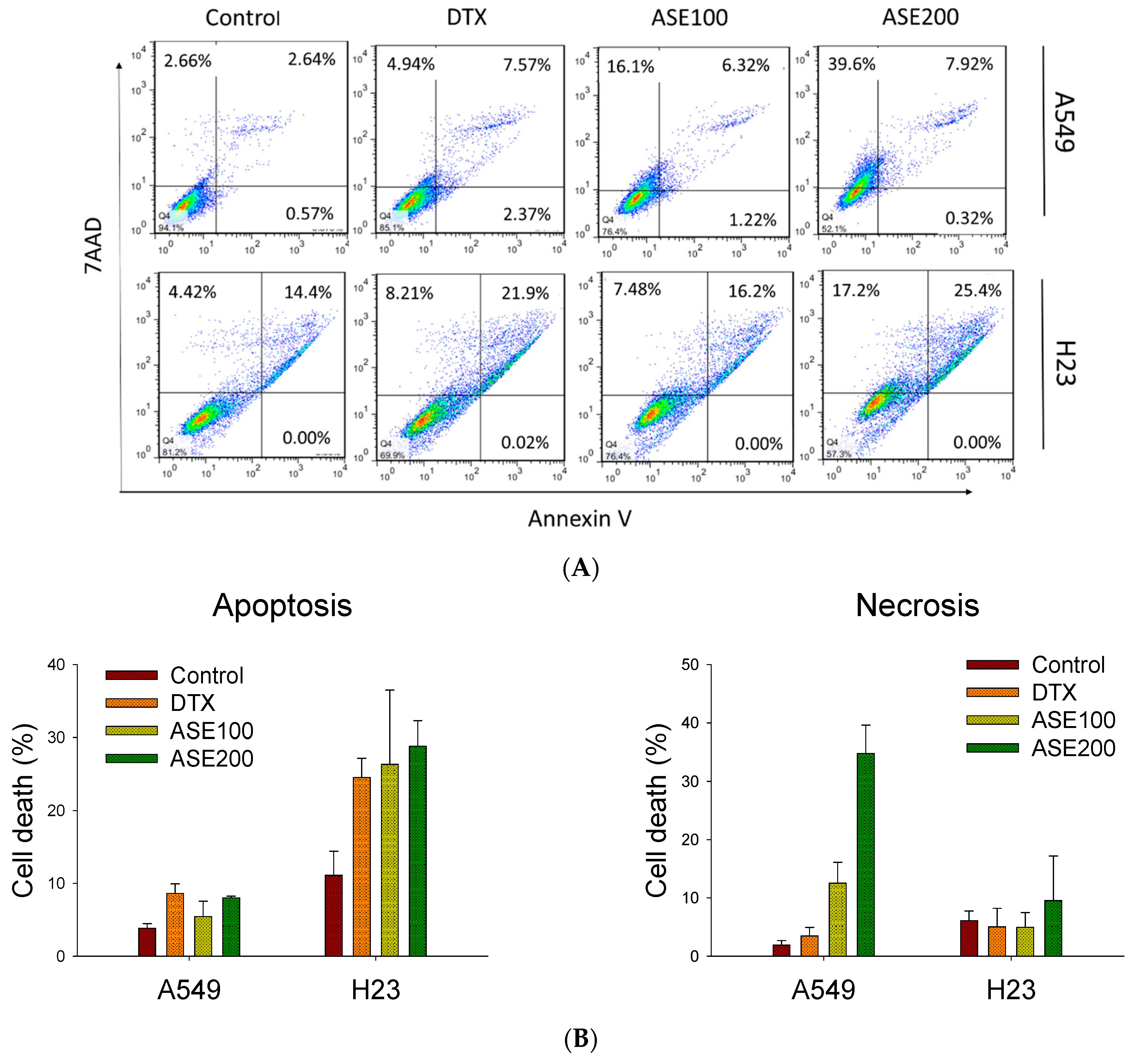
2.6. Apoptosis Analysis
2.7. Analysis of ROS
2.8. Analysis of Lipid ROS
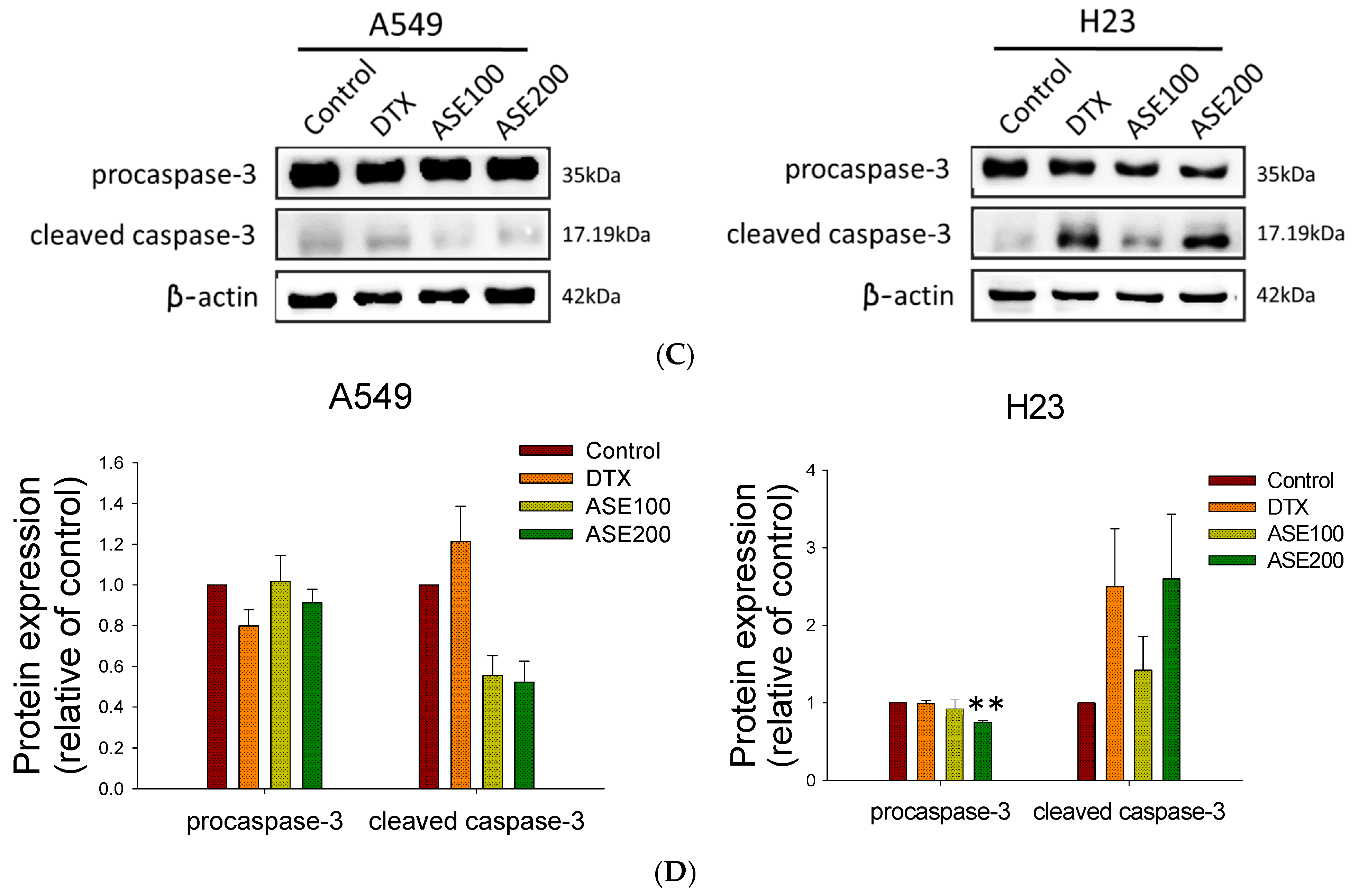
2.9. Western Blot Analysis
2.10. Liquid Chromatography (LC)-Tandem Mass Spectrometric (MS/MS) Analysis
2.11. Statistical Analysis
3. Results
3.1. ASE Inhibits A549 and H23 Cell Growth
3.2. ASE Triggers Different Cell Death Features
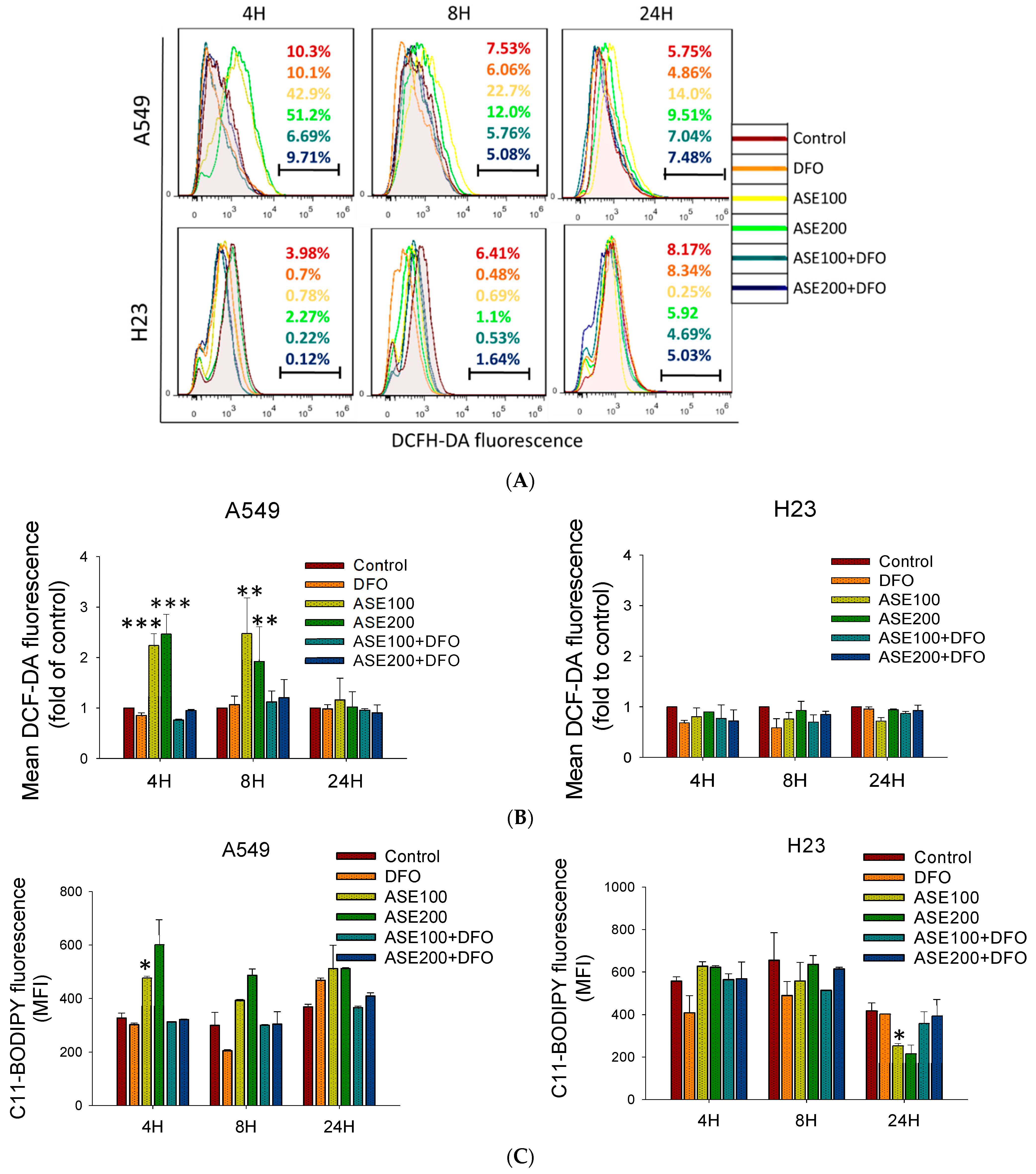
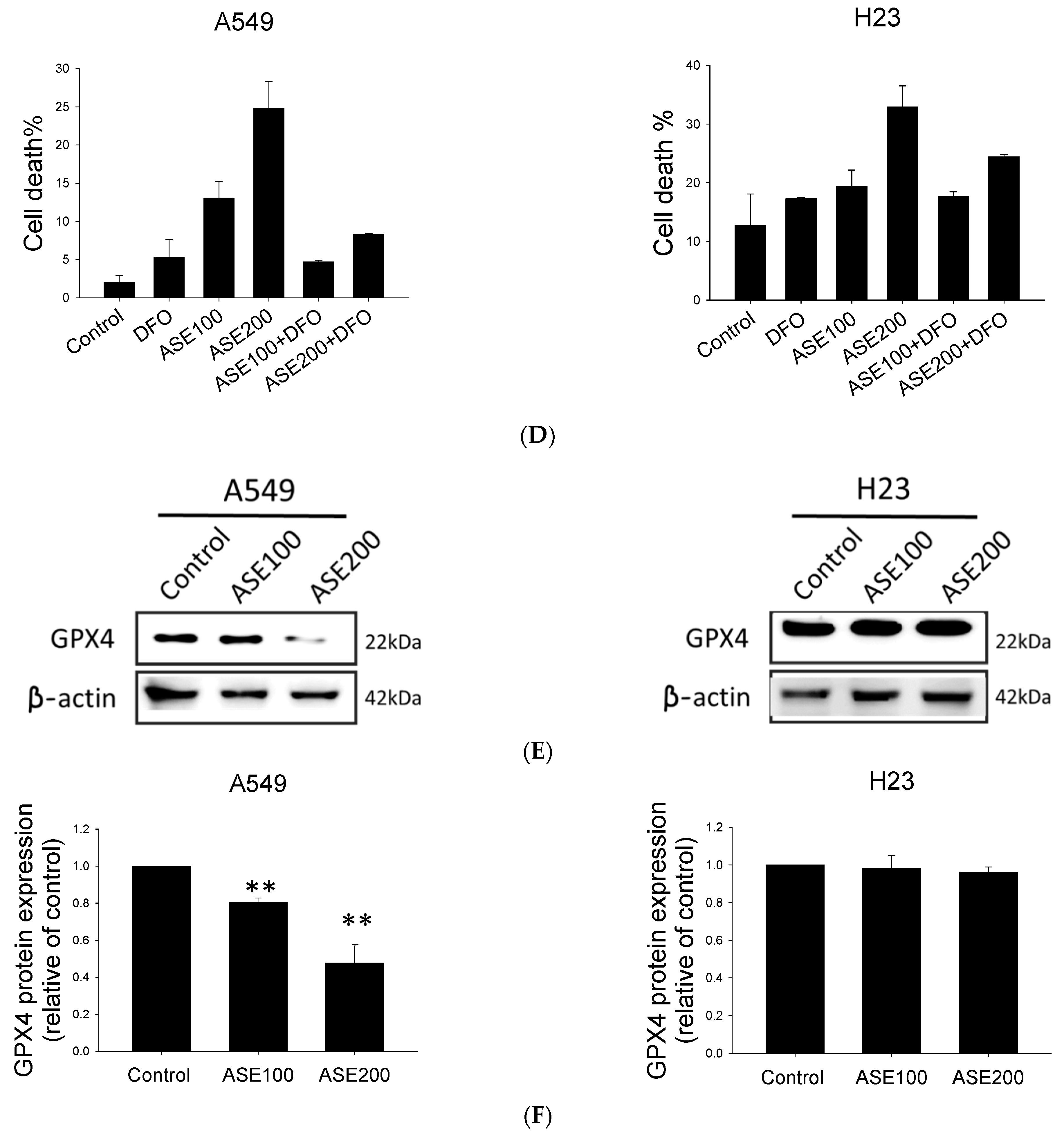
3.3. ASE Induced Ferroptosis Domineered in the A549 Cell Line
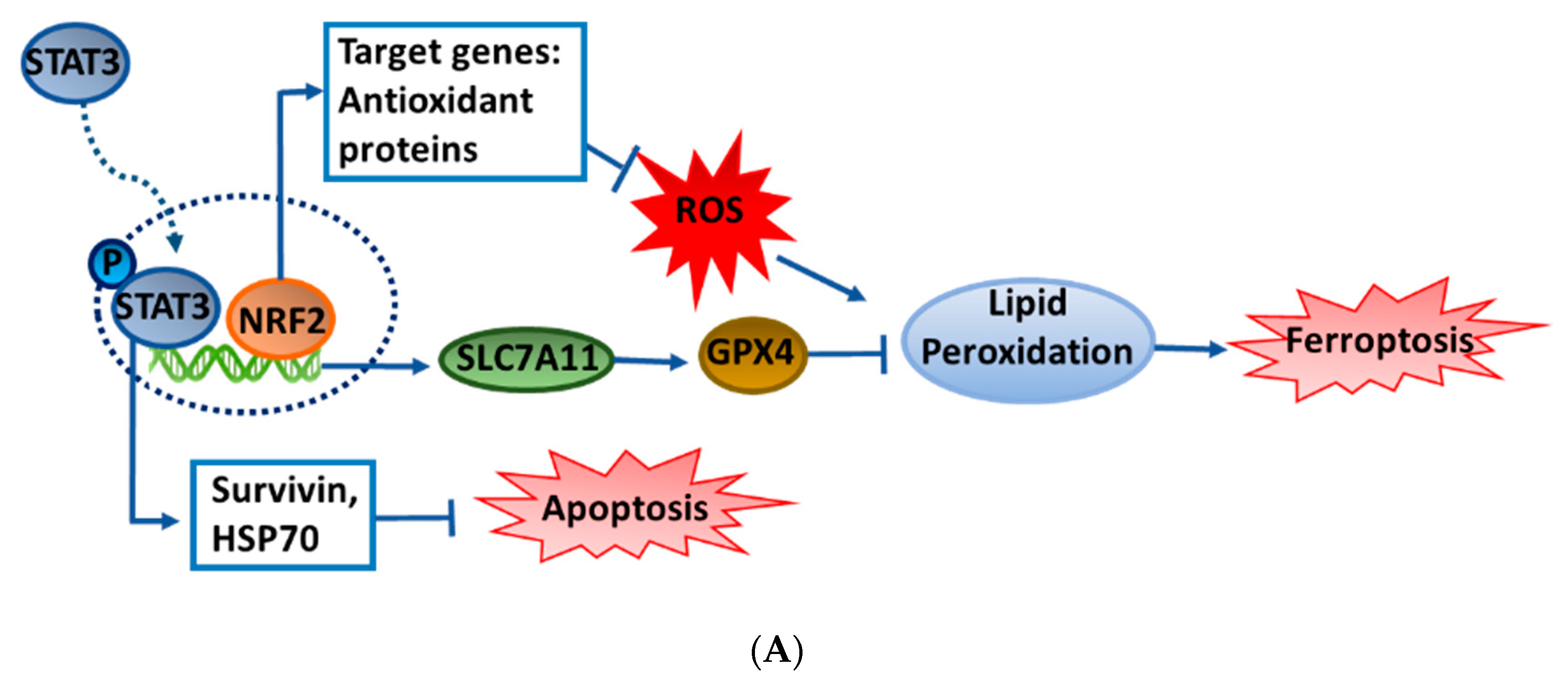
3.4. ASE Dual-Selectively Targets the NRF2/STAT3 Axis and Suppresses Survivin and HSP70 Expressions
3.5. Chemical Profile of ASE
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M.J. Cancer statistics, 2009. CA Cancer J. Clin. 2009, 59, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, D.; Cormier, J.N.; Xing, Y.; Liu, C.C.; Xia, R.; Du, X.L. Chemotherapy-associated toxicity in a large cohort of elderly patients with non-small cell lung cancer. J. Thorac. Oncol. 2010, 5, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Longley, D.B.; Johnston, P.G. Molecular mechanisms of drug resistance. J. Pathol. 2005, 205, 275–292. [Google Scholar] [CrossRef]
- Mukherjee, P.; Bahadur, S.; Harwansh, R.; Biswas, S.; Banerjee, S. Paradigm shift in natural product research: Traditional medicine inspired approaches. Phytochem. Rev. 2016, 16, 803–826. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Berman, B.M.; Moudgil, K.D. Herbal medicinal products target defined biochemical and molecular mediators of inflammatory autoimmune arthritis. Bioorg. Med. Chem. 2011, 19, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Zhao, C.; Deng, L.I.; Chen, J.; Yu, B.; Wu, X.; Pang, P.; Chen, X. Efficacy of traditional Chinese medicine in treating cancer. Biomed. Rep. 2016, 4, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Von Wehrden, H.; Wesche, K.; Miehe, G. Plant communities of the southern Mongolian Gobi. Phytocoenologia 2009, 39, 331–376. [Google Scholar] [CrossRef]
- Wesche, K.; Ronnenberg, K. Phytosociological affinities and habitat preferences of Juniperus sabina L. and Artemisia santolinifolia Turcz. ex Bess. in mountain sites of the south-eastern Gobi Altay, Mongolia. Feddes Repert. Z. Bot. Taxon. Und Geobot. 2004, 115, 585–600. [Google Scholar] [CrossRef]
- Tan, R.X.; Zheng, W.F.; Tang, H.Q. Biologically active substances from the genus Artemisia. Planta Med. 1998, 64, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, L.E.; Bates, P.L.; Evans, T.M.; Unwin, M.M.; Estes, J.R. Molecular phylogeny of Subtribe Artemisiinae (Asteraceae), including Artemisia and its allied and segregate genera. BMC Evol. Biol. 2002, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Aglarova, A.; Zilfikarov, I.; Severtseva, O. Biological characteristics and useful properties of tarragon (Artemisia dracunculus L.). Pharm. Chem. J. 2008, 42, 81–86. [Google Scholar] [CrossRef]
- Hussain, A.; Hayat, M.Q.; Sahreen, S.; Ain, Q.U.; Bokhari, S.A. Pharmacological promises of genus Artemisia (Asteraceae): A review. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2017, 54, 265–287. [Google Scholar]
- Ligaa, U. Medicinal Plants of Mongolia Used in Mongolian Traditional Medicine; KCA Press: Seoul, Korea, 1996; pp. 333–334. [Google Scholar]
- Choi, E.; Park, H.; Lee, J.; Kim, G. Anticancer, antiobesity, and anti-inflammatory activity of Artemisia species in vitro. J. Tradit. Chin. Med. 2013, 33, 92–97. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. The genus Artemisia: A comprehensive review. Pharm. Biol. 2011, 49, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Richard, A.J.; Burris, T.P.; Sanchez-Infantes, D.; Wang, Y.; Ribnicky, D.M.; Stephens, J.M. Artemisia extracts activate PPARgamma, promote adipogenesis, and enhance insulin sensitivity in adipose tissue of obese mice. Nutrition 2014, 30, S31–S36. [Google Scholar] [CrossRef] [Green Version]
- Ronnenberg, K. Reproductive ecology of two common woody species, Juniperus sabina and Artemisia santolinifolia, in mountain steppes of southern Mongolia. Erforsch. Biol. Ressour. Mong. 2005, 9, 207–223. [Google Scholar]
- Moulaie, S.; Mirzaie, A.; Aliasgari, E. Antibacterial and anticancer activities of silver nanoparticles fabricated by the Artemisia scoparia extract against lung cancer cell line (A549). Feyz J. Kashan Univ. Med. Sci. 2018, 22, 487–496. [Google Scholar]
- Lian, G.; Li, F.; Yin, Y.; Chen, L.; Yang, J. Herbal extract of Artemisia vulgaris (mugwort) induces antitumor effects in HCT-15 human colon cancer cells via autophagy induction, cell migration suppression and loss of mitochondrial membrane potential. J. BUON 2018, 23, 73–78. [Google Scholar]
- Ali, A.N.M.; Saeed, N.; Omear, H.A. The Anticancer Properties of Artemisia aucheri Boiss Extract on HT29 Colon Cancer Cells. J. Gastrointest. Cancer 2021, 52, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.J.; Schmiech, M.; Hafner, S.; Paetz, C.; Steinborn, C.; Huber, R.; Gaafary, M.E.; Werner, K.; Schmidt, C.Q.; Syrovets, T.; et al. Antitumor activity of an Artemisia annua herbal preparation and identification of active ingredients. Phytomedicine 2019, 62, 152962. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.; Fang, R.; Techatanawat, I.; Steventon, G.; Hylands, P.J.; Lee, C.C. The sulphorhodamine (SRB) assay and other approaches to testing plant extracts and derived compounds for activities related to reputed anticancer activity. Methods 2007, 42, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.K.; Chen, K.H.; Leu, Y.L.; Way, T.D.; Wang, L.W.; Chen, Y.J.; Liu, Y.M. Ethanol extracts of Cinnamomum kanehirai Hayata leaves induce apoptosis in human hepatoma cell through caspase-3 cascade. Onco Targets Ther. 2015, 8, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.M.; Tang, H.L. Cell recovery by reversal of ferroptosis. Biol. Open 2019, 8, bio043182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efferth, T. From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017, 46, 65–83. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Proneth, B.; Conrad, M. Ferroptosis and necroinflammation, a yet poorly explored link. Cell Death Differ. 2019, 26, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Thayyullathil, F.; Cheratta, A.R.; Alakkal, A.; Subburayan, K.; Pallichankandy, S.; Hannun, Y.A.; Galadari, S. Acid sphingomyelinase-dependent autophagic degradation of GPX4 is critical for the execution of ferroptosis. Cell Death Dis. 2021, 12, 26. [Google Scholar] [CrossRef]
- Wang, D.; Zhong, B.; Li, Y.; Liu, X. Dihydroartemisinin increases apoptosis of colon cancer cells through targeting Janus kinase 2/signal transducer and activator of transcription 3 signaling. Oncol. Lett. 2018, 15, 1949–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Wang, K. The induction of ferroptosis by impairing STAT3/Nrf2/GPx4 signaling enhances the sensitivity of osteosarcoma cells to cisplatin. Cell Biol. Int. 2019, 43, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Minker, C.; Duban, L.; Karas, D.; Jarvinen, P.; Lobstein, A.; Muller, C.D. Impact of Procyanidins from Different Berries on Caspase 8 Activation in Colon Cancer. Oxid. Med. Cell. Longev. 2015, 2015, 154164. [Google Scholar] [CrossRef] [PubMed]
- Im, E.; Martinez, J.D. Ursodeoxycholic acid (UDCA) can inhibit deoxycholic acid (DCA)-induced apoptosis via modulation of EGFR/Raf-1/ERK signaling in human colon cancer cells. J. Nutr. 2004, 134, 483–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Guo, G.F.; Zhang, B. Research of Brucea javanica against cancer. Chin. J. Integr. Med. 2017, 23, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Ning, N.; Li, Y.; Zhang, Q.F.; Xie, Y.C.; Irshad, M.; Feng, X.; Tao, X.J. Biatractylolide Modulates PI3K-Akt-GSK3beta-Dependent Pathways to Protect against Glutamate-Induced Cell Damage in PC12 and SH-SY5Y Cells. Evid. Based Complement. Altern. Med. 2017, 2017, 1291458. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Zhou, Y.; Jiang, Z.; Zhao, Y.; Zhang, D.; Cong, X.; Cao, R.; Li, H.; Tian, W. Cytotoxic and chemosensitization effects of Scutellarin from traditional Chinese herb Scutellaria altissima L. in human prostate cancer cells. Oncol. Rep. 2017, 38, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhang, S. Scutellarin-induced apoptosis in HepG2 hepatocellular carcinoma cells via a STAT3 pathway. Phytother. Res. 2013, 27, 1524–1528. [Google Scholar] [CrossRef]
- Kim, D.S.; Jeon, B.K.; Lee, Y.E.; Woo, W.H.; Mun, Y.J. Diosgenin Induces Apoptosis in HepG2 Cells through Generation of Reactive Oxygen Species and Mitochondrial Pathway. Evid. Based Complement. Altern. Med. 2012, 2012, 981675. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Tan, H.Y.; Chen, F.; Wang, N.; Feng, Y. Targeting Cancer Metabolism to Resensitize Chemotherapy: Potential Development of Cancer Chemosensitizers from Traditional Chinese Medicines. Cancers 2020, 12, 404. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, F.; Mojarrab, M.; Ghazi-Khansari, M.; Hosseinzadeh, L. A semipolar fraction of petroleum ether extract of Artemisia aucheri induces apoptosis and enhances the apoptotic response to doxorubicin in human neuroblastoma SKNMC cell line. Res. Pharm. Sci. 2015, 10, 335–344. [Google Scholar] [PubMed]
- Kim, E.J.; Kim, G.T.; Kim, B.M.; Lim, E.G.; Kim, S.Y.; Kim, Y.M. Apoptosis-induced effects of extract from Artemisia annua Linne by modulating PTEN/p53/PDK1/Akt/ signal pathways through PTEN/p53-independent manner in HCT116 colon cancer cells. BMC Complement. Altern. Med. 2017, 17, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooko, E.; Saeed, M.E.; Kadioglu, O.; Sarvi, S.; Colak, M.; Elmasaoudi, K.; Janah, R.; Greten, H.J.; Efferth, T. Artemisinin derivatives induce iron-dependent cell death (ferroptosis) in tumor cells. Phytomedicine 2015, 22, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yu, Q.; Huo, C.; Li, Y.; He, L.; Ran, B.; Chen, J.; Li, Y.; Liu, W. Ferroptosis: A Novel Mechanism of Artemisinin and its Derivatives in Cancer Therapy. Curr. Med. Chem. 2021, 28, 329–345. [Google Scholar] [CrossRef]
- Lei, P.; Bai, T.; Sun, Y. Mechanisms of Ferroptosis and Relations With Regulated Cell Death: A Review. Front. Physiol. 2019, 10, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, K.; Hayano, M.; Pagano, N.C.; Stockwell, B.R. Cell-Line Selectivity Improves the Predictive Power of Pharmacogenomic Analyses and Helps Identify NADPH as Biomarker for Ferroptosis Sensitivity. Cell Chem. Biol. 2016, 23, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodson, M.; Castro-Portuguez, R.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Y.; Wang, P.; Zhang, J.; Chen, H.; Zhang, L.; Du, X.; Zhao, C.; Wu, D.; Liu, F.; et al. A comprehensive review of integrative pharmacology-based investigation: A paradigm shift in traditional Chinese medicine. Acta Pharm. Sin. B 2021, 11, 1379–1399. [Google Scholar] [CrossRef]
- Silva, M.M.; Rocha, C.R.R.; Kinker, G.S.; Pelegrini, A.L.; Menck, C.F.M. The balance between NRF2/GSH antioxidant mediated pathway and DNA repair modulates cisplatin resistance in lung cancer cells. Sci. Rep. 2019, 9, 17639. [Google Scholar] [CrossRef]
- Barry, S.P.; Townsend, P.A.; McCormick, J.; Knight, R.A.; Scarabelli, T.M.; Latchman, D.S.; Stephanou, A. STAT3 deletion sensitizes cells to oxidative stress. Biochem. Biophys. Res. Commun. 2009, 385, 324–329. [Google Scholar] [CrossRef]
- Qiang, Z.; Dong, H.; Xia, Y.; Chai, D.; Hu, R.; Jiang, H. Nrf2 and STAT3 Alleviates Ferroptosis-Mediated IIR-ALI by Regulating SLC7A11. Oxid. Med. Cell. Longev. 2020, 2020, 5146982. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Gao, H.; Yuan, R.; Han, S.; Li, X.X.; Tang, M.; Dong, B.; Li, J.X.; Zhao, L.C.; Feng, J.; et al. Procyanidin A2, a polyphenolic compound, exerts anti-inflammatory and anti-oxidative activity in lipopolysaccharide-stimulated RAW264.7 cells. PLoS ONE 2020, 15, e0237017. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-Y.; Feng, X.-H.; Zhao, P.-F.; Damirin, A.; Ma, C.-M. Procyanidin A2 penetrates L-02 cells and protects against tert-butyl hydroperoxide-induced oxidative stress by activating Nrf2 through JNK and p38 phosphorylation. J. Funct. Foods 2019, 62, 103562. [Google Scholar] [CrossRef]
- Lv, W.L.; Liu, Q.; An, J.H.; Song, X.Y. Scutellarin inhibits hypoxia-induced epithelial-mesenchymal transition in bladder cancer cells. J. Cell. Physiol. 2019, 234, 23169–23175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Chen, W.Y.; Li, X.B.; Ke, H.; Zhou, X.L. Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-beta1/smad2/ROS/caspase-3 pathway. Open Life Sci. 2021, 16, 961–968. [Google Scholar] [CrossRef]
- Qian, Q.; Chen, W.; Cao, Y.; Cao, Q.; Cui, Y.; Li, Y.; Wu, J. Targeting Reactive Oxygen Species in Cancer via Chinese Herbal Medicine. Oxid. Med. Cell. Longev. 2019, 2019, 9240426. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Compound Formula | Observed RT (min) | Observed m/z | Mass Error (ppm) | Reference |
|---|---|---|---|---|---|---|
| 1. | Procyanidin A2 | C30H24O12 | 5.56 | 577.1344 | 0.6 | [34] |
| 2. | Nor-ψ-tropine | C8H15NO | 18.97 | 128.1062 | −6.1 | - |
| 3. | Hyodeoxycholic acid | C24H40O4 | 18.54 | 393.298 | −4.9 | [35] |
| 4. | Nigakilactone H | C22H32O8 | 6.69 | 425.2141 | −6.8 | [36] |
| 5. | Siraitic acid C | C29H44O5 | 14.72 | 441.3018 | 4.3 | - |
| 6. | 16-O-Acetylisoiridogermanal | C46H78O6 | 6.98 | 517.3895 | 1.5 | - |
| 7. | Blestrianol B | C37H32O7 | 19.03 | 589.2231 | 1.7 | - |
| 8. | Biatractylolide | C30H38O4 | 6.43 | 463.2882 | 8.3 | [37] |
| 9. | Scutellarin | C21H18O12 | 19.11 | 449.1113 | 7.7 | [38,39] |
| 10. | Protodiosgenin | C27H42O3 | 6.33 | 595.389 | 8.3 | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batbold, U.; Liu, J.-J. Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators. Appl. Sci. 2022, 12, 4868. https://doi.org/10.3390/app12104868
Batbold U, Liu J-J. Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators. Applied Sciences. 2022; 12(10):4868. https://doi.org/10.3390/app12104868
Chicago/Turabian StyleBatbold, Uyanga, and Jun-Jen Liu. 2022. "Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators" Applied Sciences 12, no. 10: 4868. https://doi.org/10.3390/app12104868
APA StyleBatbold, U., & Liu, J.-J. (2022). Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators. Applied Sciences, 12(10), 4868. https://doi.org/10.3390/app12104868