
Effects of Endurance Cycling on Mechanomyographic Median Power Frequency of the Vastus Lateralis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. VO2MAX Testing
2.3. Ultrasound Imaging
2.4. Isometric Strength Testing
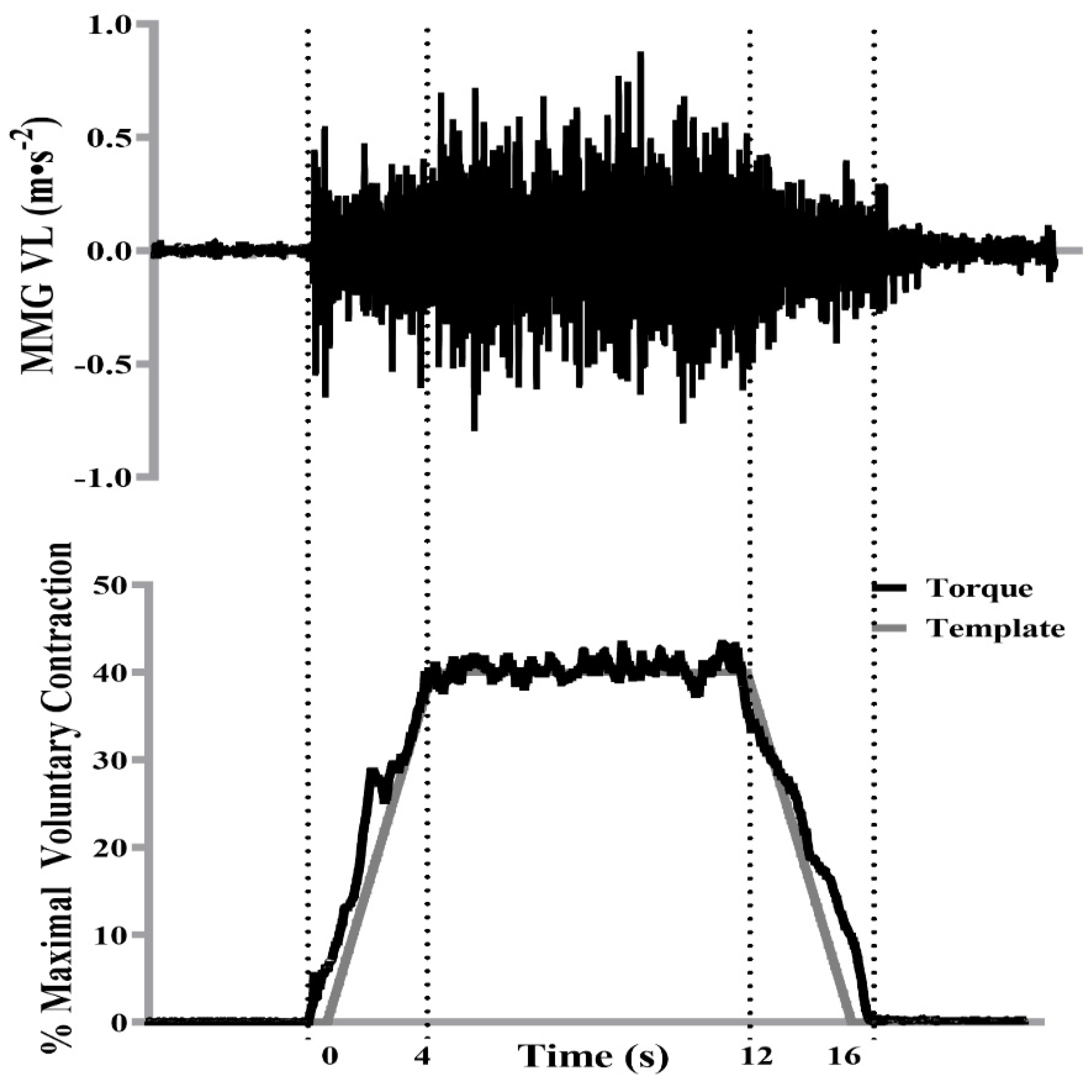
2.5. Mechanomyography and Signal Processing
2.6. Continuous Cycle Training
2.7. Statistical Analysis
3. Results
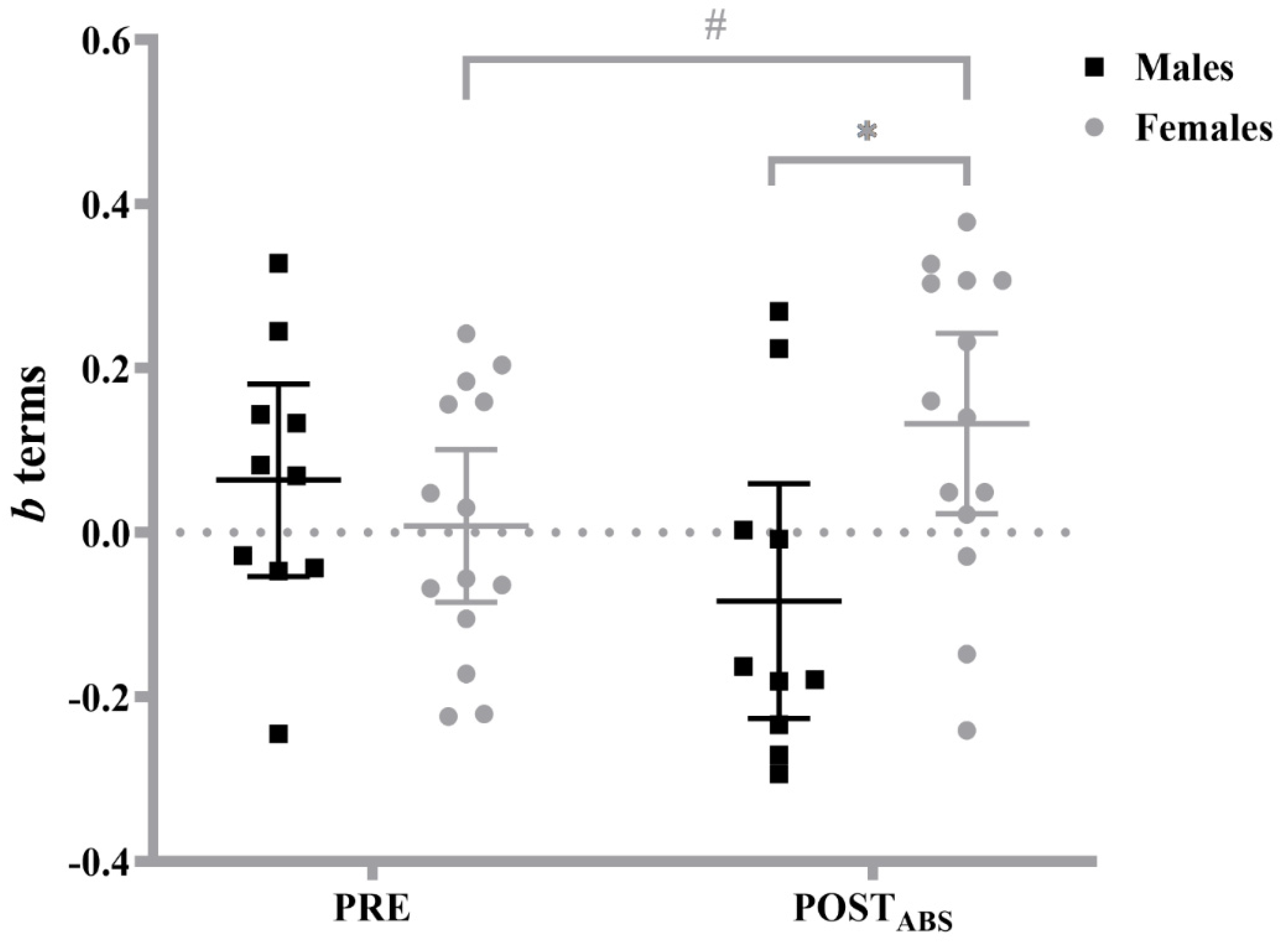
3.1. Log-Transformed MMGMDF–Torque Relationships
3.2. Steady Torque Segment
3.3. Correlations
3.4. sFAT Matched Subset Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orizio, C. Muscle sound: Bases for the introduction of a mechanomyographic signal in muscle studies. Crit. Rev. Biomed. Eng. 1993, 21, 201–243. [Google Scholar] [PubMed]
- Barry, D.; Cole, N. Muscle sounds are emitted at the resonant frequencies of skeletal muscle. IEEE Trans. Biomed. Eng. 1990, 37, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Coburn, J.W.; Housh, T.J.; Cramer, J.T.; Weir, J.P.; Miller, J.M.; Beck, T.W.; Malek, M.H.; Johnson, G.O. Mechanomyographic and electromyographic responses of the vastus medialis muscle during isometric and concentric muscle actions. J. Strength Cond. Res. 2005, 19, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.W.; Housh, T.J.; Johnson, G.O.; Cramer, J.T.; Weir, J.P.; Coburn, J.W.; Malek, M.H. Does the frequency content of the surface mechanomyographic signal reflect motor unit firing rates? A brief review. J. Electromyogr. Kinesiol. 2007, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Herda, T.J.; Ryan, E.D.; Beck, T.W.; Costa, P.B.; DeFreitas, J.M.; Stout, J.R.; Cramer, J.T. Reliability of mechanomyographic amplitude and mean power frequency during isometric step and ramp muscle actions. J. Neurosci. Methods 2008, 171, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.D.; Beck, T.W.; Herda, T.J.; Hartman, M.J.; Stout, J.R.; Housh, T.J.; Cramer, J.T. Mechanomyographic amplitude and mean power frequency responses during isometric ramp vs. step muscle actions. J. Neurosci. Methods 2008, 168, 293–305. [Google Scholar] [CrossRef]
- Trevino, M.A.; Herda, T.J. Mechanomyographic mean power frequency during an isometric trapezoid muscle action at multiple contraction intensities. Physiol. Meas. 2015, 36, 1383–1397. [Google Scholar] [CrossRef]
- Vila-Chã, C.; Falla, D.; Farina, D. Motor unit behavior during submaximal contractions following six weeks of either endurance or strength training. J. Appl. Physiol. 2010, 109, 1455–1466. [Google Scholar] [CrossRef] [Green Version]
- Trevino, M.A.; Dimmick, H.L.; Parra, M.E.; Sterczala, A.J.; Miller, J.D.; Deckert, J.A.; Gallagher, P.M.; Fry, A.C.; Weir, J.P.; Herda, T.J. Effects of continuous cycling training on motor unit firing rates, input excitation, and myosin heavy chain of the vastus lateralis in sedentary females. Exp. Brain Res. 2022, 240, 825–839. [Google Scholar] [CrossRef]
- Martinez-Valdes, E.; Falla, D.; Negro, F.; Mayer, F.; Farina, D. Differential Motor Unit Changes after Endurance or High-Intensity Interval Training. Med. Sci. Sports Exerc. 2017, 49, 1126–1136. [Google Scholar] [CrossRef] [Green Version]
- Sontag, S.A.; Trevino, M.A.; Herda, T.J.; Sterczala, A.J.; Miller, J.D.; Parra, M.E.; Dimmick, H.L.; Deckert, J. Endurance training alters motor unit activation strategies for the vastus lateralis, yet sex-related differences and relationships with muscle size remain. Eur. J. Appl. Physiol. 2021, 121, 1367–1377. [Google Scholar] [CrossRef]
- Beck, T.W.; Housh, T.J.; Cramer, J.T.; Weir, J.P.; Johnson, G.O.; Coburn, J.W.; Malek, M.H.; Mielke, M. Mechanomyographic amplitude and frequency responses during dynamic muscle actions: A comprehensive review. Biomed. Eng. Online 2005, 4, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orizio, C.; Perini, R.; Veicsteinas, A. Muscular sound and force relationship during isometric contraction in man. Eur. J. Appl. Physiol. Occup. Physiol. 1989, 58, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Zwarts, M.J.; Keidel, M. Relationship between electrical and vibratory output of muscle during voluntary contraction and fatigue. Muscle Nerve 1991, 14, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Farina, D.; Merletti, R.; Enoka, R.M. The extraction of neural strategies from the surface EMG: An update. J. Appl. Physiol. 2014, 117, 1215–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herda, T.J.; Housh, T.J.; Fry, A.C.; Weir, J.P.; Schilling, B.; Ryan, E.D.; Cramer, J.T. A noninvasive, log-transform method for fiber type discrimination using mechanomyography. J. Electromyogr. Kinesiol. 2010, 20, 787–794. [Google Scholar] [CrossRef]
- Beck, T.W.; Housh, T.J.; Fry, A.C.; Cramer, J.; Weir, J.P.; Schilling, B.; Falvo, M.J.; Moore, C.A. The influence of muscle fiber type composition on the patterns of responses for electromyographic and mechanomyographic amplitude and mean power frequency during a fatiguing submaximal isometric muscle action. Electromyogr. Clin. Neurophysiol. 2007, 47, 221–232. [Google Scholar]
- Howald, H.; Hoppeler, H.; Claassen, H.; Mathieu, O.; Straub, R. Influences of endurance training on the ultrastructural composition of the different muscle fiber types in humans. Pflüg. Archiv. 1985, 403, 369–376. [Google Scholar] [CrossRef]
- Garnett, R.A.; O’Donovan, M.; Stephens, J.A.; Taylor, A. Motor unit organization of human medial gastrocnemius. J. Physiol. 1979, 287, 33–43. [Google Scholar] [CrossRef]
- Whaley, M.H.; Brubaker, P.H.; Otto, R.M.; Armstrong, L.E. ACSM’s Guidelines for Exercise Testing and Prescription; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- Beck, T.W.; DeFreitas, J.M.; Stock, M.S.; Dillon, M.A. An examination of mechanomyographic signal stationarity during concentric isokinetic, eccentric isokinetic and isometric muscle actions. Physiol. Meas. 2010, 31, 339–361. [Google Scholar] [CrossRef]
- Karvonen, M.; Kentala, K.; Mustala, O. The effects of training heart rate: A longitudinal study. Ann. Med. Exp. Biol. Fenn. 1957, 35, 307–315. [Google Scholar] [PubMed]
- Lounana, J.; Campion, F.; Noakes, T.D.; Medelli, J. Relationship between %HRmax, %HR Reserve, %VO2max, and %VO2 Reserve in Elite Cyclists. Med. Sci. Sports Exerc. 2007, 39, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Swain, D.P. Energy Cost Calculations for Exercise Prescription. Sports Med. 2000, 30, 17–22. [Google Scholar] [CrossRef]
- Trevino, M.A.; Herda, T.J. The effects of training status and muscle action on muscle activation of the vastus lateralis. Acta Bioeng. Biomech. 2015, 17, 107–114. [Google Scholar]
- Trevino, M.A.; Herda, T.J. The effects of chronic exercise training status on motor unit activation and deactivation control strategies. J. Sports Sci. 2015, 34, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Herda, T.J.; Vardiman, J.P.; Gallagher, P.M.; Fry, A.C. Relationships between skinfold thickness and electromyographic and mechanomyographic amplitude recorded during voluntary and non-voluntary muscle actions. J. Electromyogr. Kinesiol. 2014, 24, 207–213. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.J.; Contessa, P. Biomechanical benefits of the Onion-Skin motor unit control scheme. J. Biomech. 2014, 48, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Farina, D.; Holobar, A.; Gazzoni, M.; Zazula, D.; Merletti, R.; Enoka, R.M. Adjustments differ among low-threshold motor units during intermittent, isometric contractions. J. Neurophysiol. 2009, 101, 350–359. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.J.; Contessa, P. Hierarchical control of motor units in voluntary contractions. J. Neurophysiol. 2012, 107, 178–195. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.J.; Kline, J.C.; Contessa, P. Transposed firing activation of motor units. J. Neurophysiol. 2014, 112, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Trevino, M.A.; Herda, T.J.; Fry, A.C.; Gallagher, P.M.; Vardiman, J.P.; Mosier, E.M.; Miller, J.D. The influence of myosin heavy chain isoform content on mechanical behavior of the vastus lateralis in vivo. J. Electromyogr. Kinesiol. 2016, 28, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Olmos, A.A.; Herda, T.J.; Sontag, S.A.; Trevino, M.A. The influence of chronic training status on the mechanical behavior of the vastus lateralis during repetitive trapezoidal contractions. J. Musculoskelet. Neuronal Interact. 2022. Available online: https://www.ismni.org/jmni/accepted/JMNI_21M-09-194.pdf (accessed on 10 April 2022).
- Abrahin, O.; Rodrigues, R.P.; Marçal, A.C.; Alves, E.A.C.; Figueiredo, R.C.; de Sousa, E.C. Swimming and cycling do not cause positive effects on bone mineral density: A systematic review. Rev. Bras. de Reum. (Engl. Ed.) 2016, 56, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedlander, A.L.; Genant, H.K.; Sadowsky, S.; Byl, N.N.; Glüer, C.-C. A two-year program of aerobics and weight training enhances bone mineral density of young women. J. Bone Miner. Res. 2009, 10, 574–585. [Google Scholar] [CrossRef]
- Campos, R.; de Mello, M.T.; Tock, L.; Silva, P.L.; Masquio, D.C.; de Piano, A.; Sanches, P.L.; Carnier, J.; Corgosinho, F.C.; Foschini, D.; et al. Aerobic Plus Resistance Training Improves Bone Metabolism and Inflammation in Adolescents who Are Obese. J. Strength Cond. Res. 2014, 28, 758–766. [Google Scholar] [CrossRef]
- Armamento-Villareal, R.; Aguirre, L.; Waters, D.L.; Napoli, N.; Qualls, C.; Villareal, D.T. Effect of Aerobic or Resistance Exercise, or Both, on Bone Mineral Density and Bone Metabolism in Obese Older Adults While Dieting: A Randomized Controlled Trial. J. Bone Miner. Res. 2019, 35, 430–439. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linearly Increasing Segment | Linearly Decreasing Segment | |||||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||
| PRE | b terms | mean | 0.163 (0.200) | 0.095 (0.188) | −0.035 (0.211) | −0.079 (0.176) |
| 95% CI | 0.039–0.287 | −0.003–0.193 | −0.166–0.096 | −0.171–0.014 | ||
| POSTABS | b terms | mean | −0.050 (0.255) | 0.124 (0.246) | −0.117 (0.168) | 0.141 (0.229) |
| 95% CI | −0.208–0.108 | −0.005–0.253 | −0.211–−0.012 | 0.021–0.260 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez, S.; Olmos, A.A.; Sontag, S.A.; Parra, M.E.; Dimmick, H.L.; Sterczala, A.J.; Miller, J.D.; Deckert, J.A.; Herda, T.J.; Trevino, M.A. Effects of Endurance Cycling on Mechanomyographic Median Power Frequency of the Vastus Lateralis. Appl. Sci. 2022, 12, 5213. https://doi.org/10.3390/app12105213
Perez S, Olmos AA, Sontag SA, Parra ME, Dimmick HL, Sterczala AJ, Miller JD, Deckert JA, Herda TJ, Trevino MA. Effects of Endurance Cycling on Mechanomyographic Median Power Frequency of the Vastus Lateralis. Applied Sciences. 2022; 12(10):5213. https://doi.org/10.3390/app12105213
Chicago/Turabian StylePerez, Sergio, Alex A. Olmos, Stephanie A. Sontag, Mandy E. Parra, Hannah L. Dimmick, Adam J. Sterczala, Jonathan D. Miller, Jake A. Deckert, Trent J. Herda, and Michael A. Trevino. 2022. "Effects of Endurance Cycling on Mechanomyographic Median Power Frequency of the Vastus Lateralis" Applied Sciences 12, no. 10: 5213. https://doi.org/10.3390/app12105213