
Plasticizer Di-(2-ethylhexyl) Phthalate and Its Metabolite Mono(2-ethylhexyl) Phthalate Inhibit Myogenesis in Differentiating Mouse and Human Skeletal Muscle Cell Models
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Primary HSMPC Culture
2.4. Cell Viability Measurement
2.5. Creatine Kinase Activity Assay
2.6. Myogenic Differentiation (Myotube Formation)
2.7. Immunoblotting
2.8. Analysis of Statistics
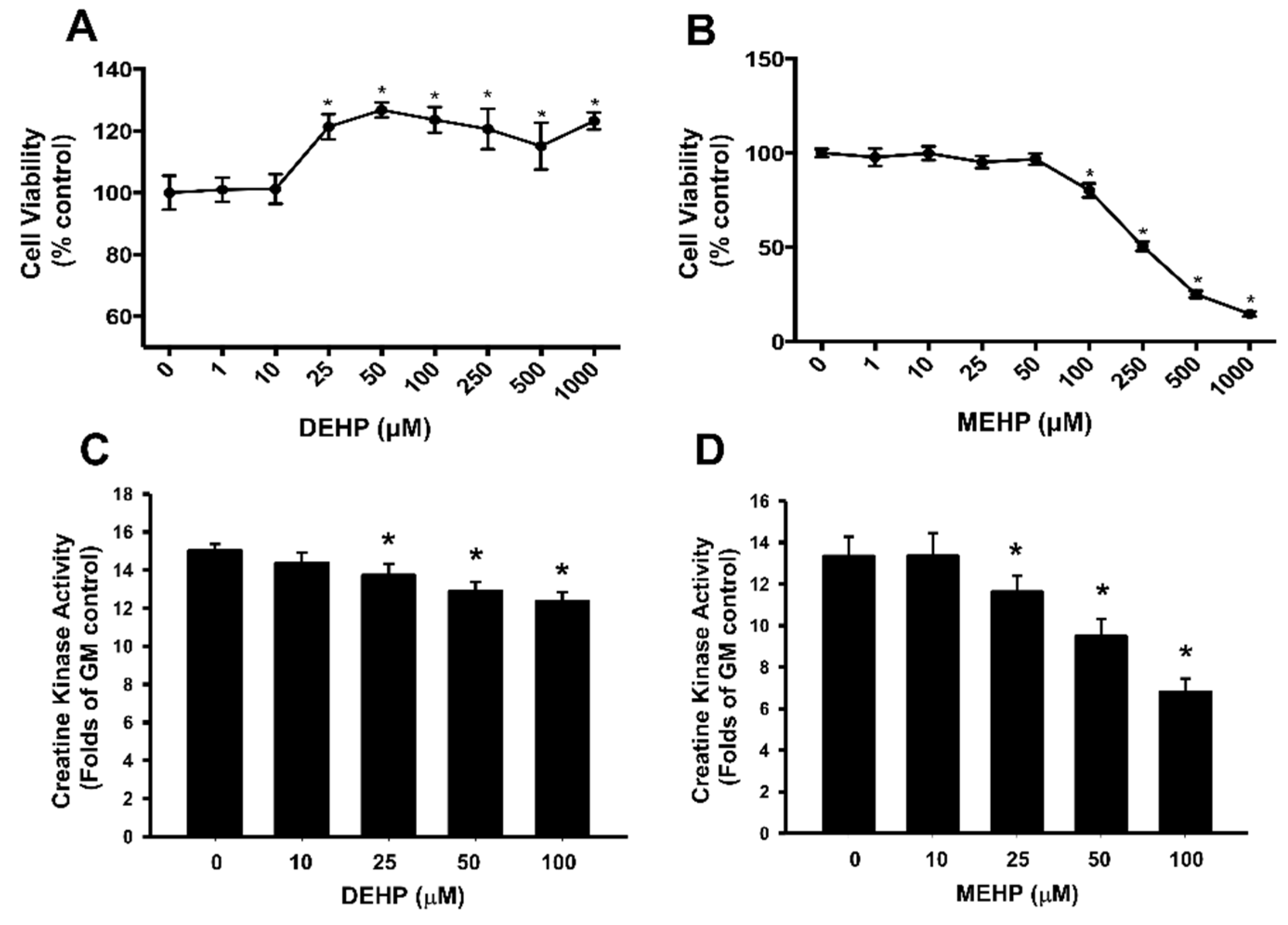
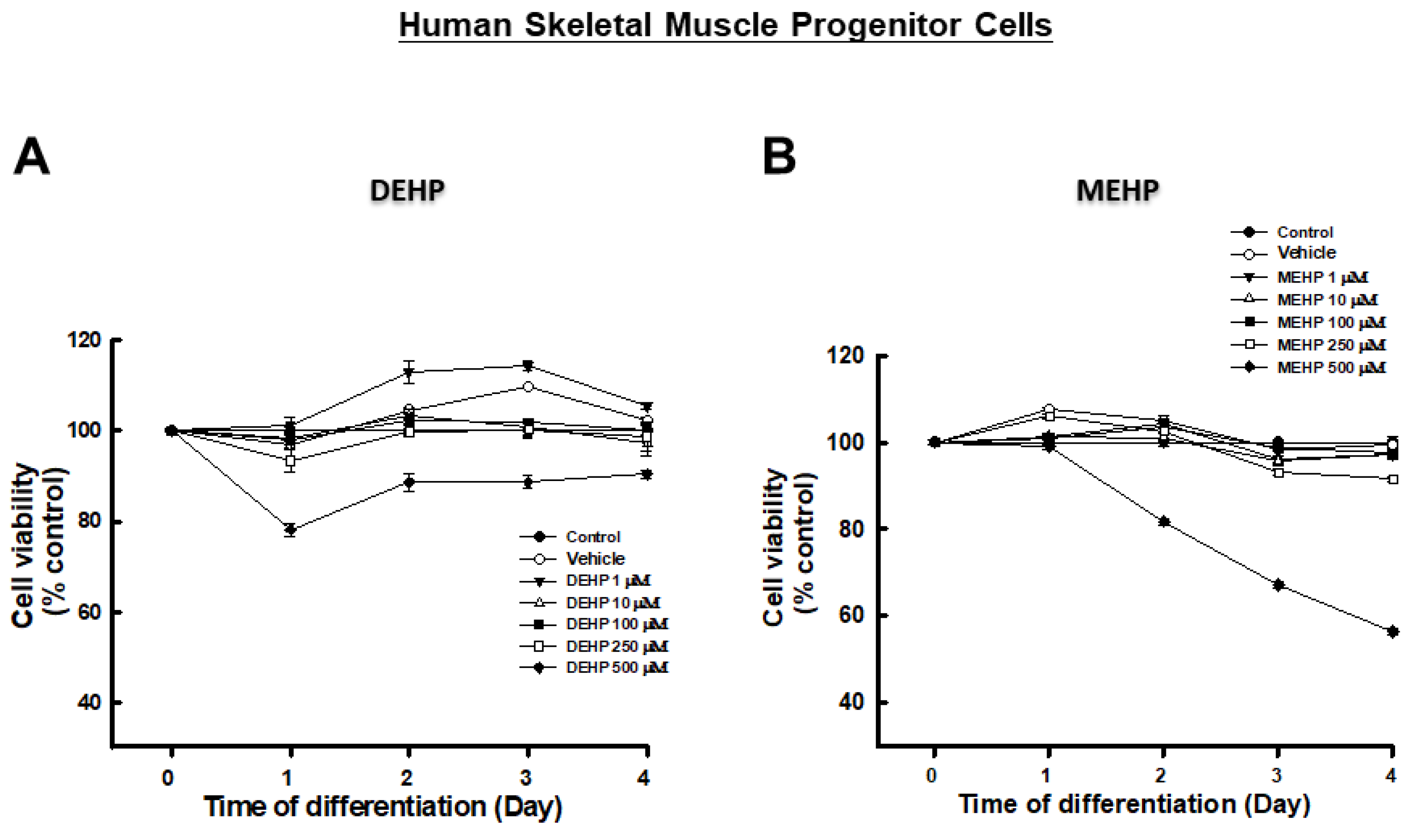
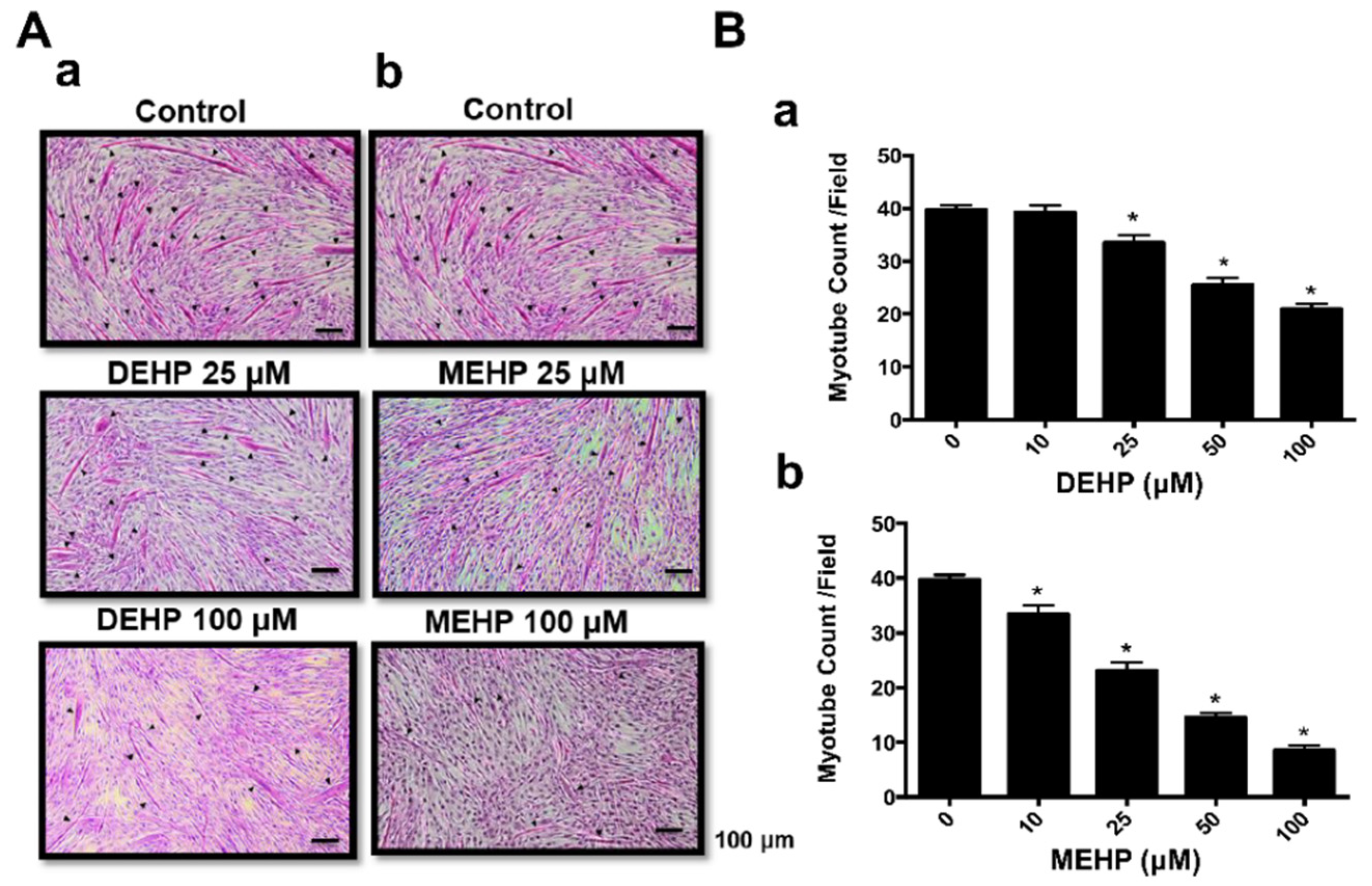
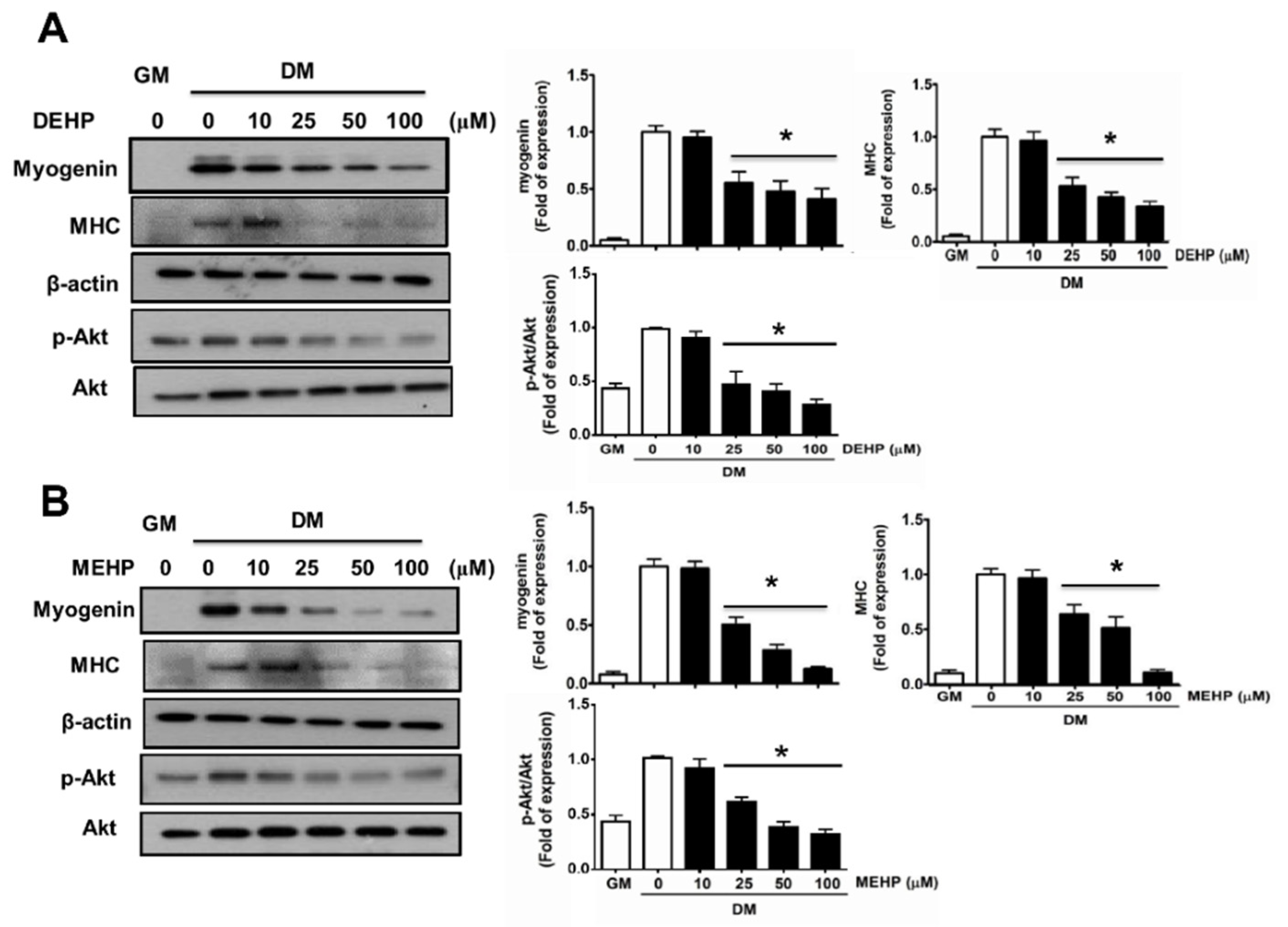
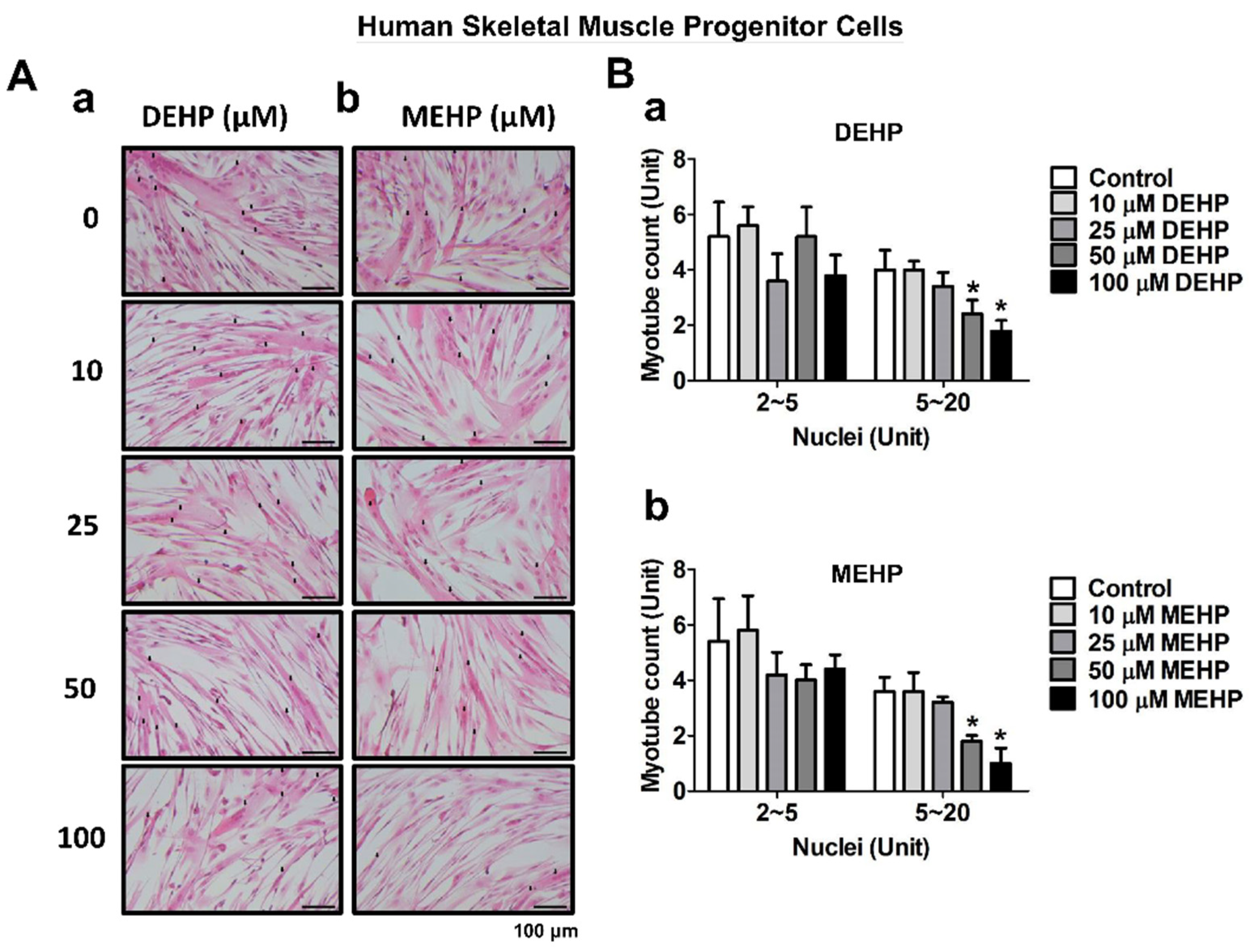
3. Results
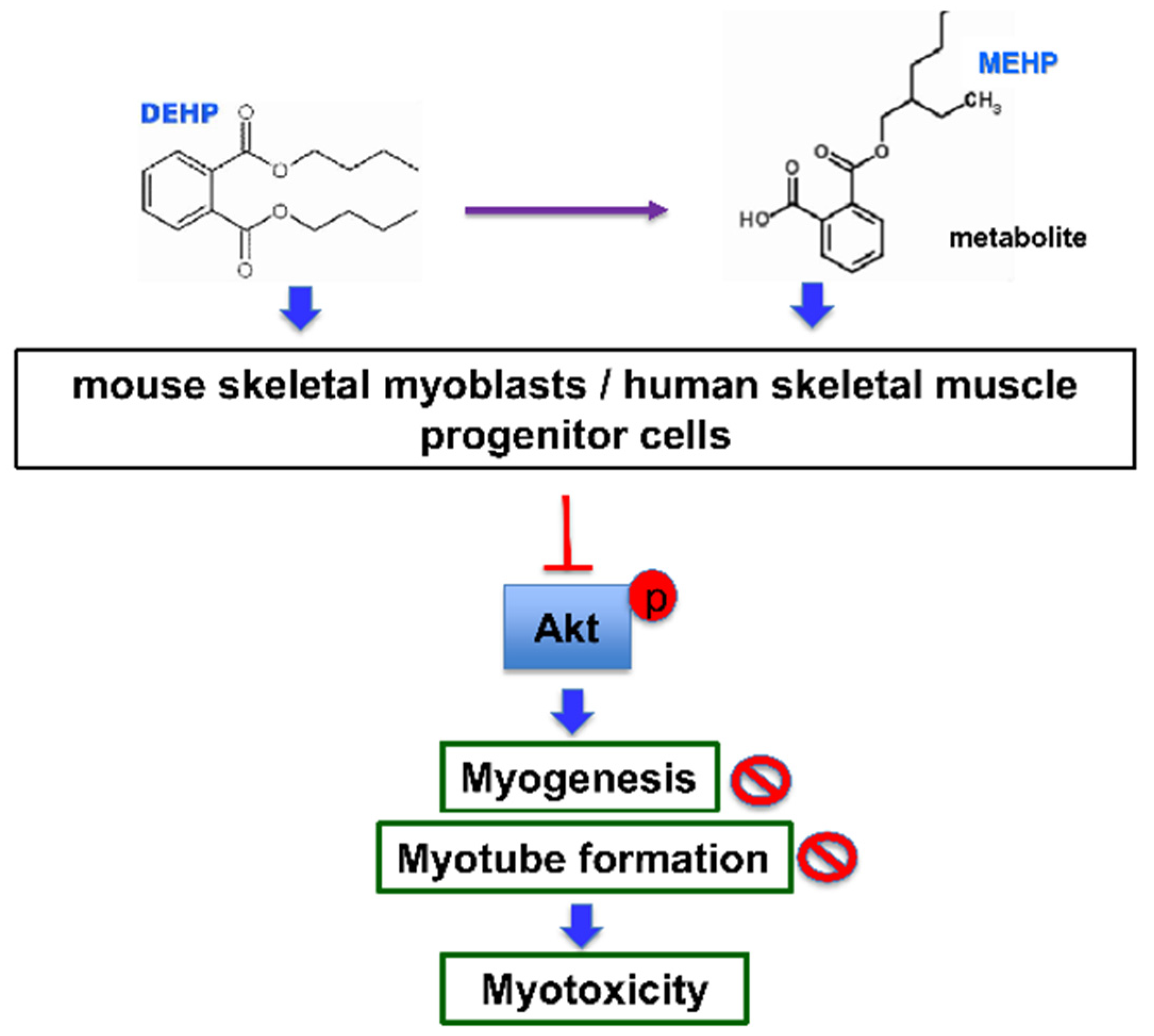
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bizzari, S.; Oppenberg, B.; Iskikawa, Y. Plasticizers Chemical Economics Handbook; SRI International: Palo Alto, CA, USA, 2000. [Google Scholar]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Needham, L.L.; Silva, M.J.; Lambert, G. Exposure to di-(2-ethylhexyl) phthalate among premature neonates in a neonatal intensive care unit. Pediatrics 2004, 113, e429–e434. [Google Scholar] [CrossRef] [PubMed]
- Hanawa, T.; Muramatsu, E.; Asakawa, K.; Suzuki, M.; Tanaka, M.; Kawano, K.; Seki, T.; Juni, K.; Nakajima, S. Investigation of the release behavior of diethylhexyl phthalate from the polyvinyl-chloride tubing for intravenous administration. Int. J. Pharm. 2000, 210, 109–115. [Google Scholar] [CrossRef]
- Ito, Y.; Yokota, H.; Wang, R.; Yamanoshita, O.; Ichihara, G.; Wang, H.; Kurata, Y.; Takagi, K.; Nakajima, T. Species differences in the metabolism of di(2-ethylhexyl) phthalate (DEHP) in several organs of mice, rats, and marmosets. Arch. Toxicol. 2005, 79, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Takehisa, H.; Naoko, E.; Masahiko, S.; Katsuhide, T.; Moriyuki, O.; Keizoh, S.; Mutsuko, T.; Kenji, K.; Shin’ichiro, N.; Toshio, O. Release behavior of diethylhexyl phthalate from the polyvinyl-chloride tubing used for intravenous administration and the plasticized PVC membrane. Int. J. Pharm. 2005, 297, 30–37. [Google Scholar] [CrossRef]
- Koch, H.M.; Bolt, H.M.; Preuss, R.; Angerer, J. New metabolites of di(2-ethylhexyl)phthalate (DEHP) in human urine and serum after single oral doses of deuterium-labelled DEHP. Arch. Toxicol. 2005, 79, 367–376. [Google Scholar] [CrossRef]
- Kavlock, R.; Boekelheide, K.; Chapin, R.; Cunningham, M.; Faustman, E.; Foster, P.; Golub, M.; Henderson, R.; Hinberg, I.; Little, R.; et al. NTP Center for the evaluation of risks to human reproduction; Phthalates expert panel report on the reproductive and developmental toxicity of di(2-ethylhexyl)phthalate. Reprod. Toxicol. 2002, 16, 529–653. [Google Scholar] [CrossRef]
- Latini, G.; De Felice, C.; Presta, G.; Del Vecchio, A.; Paris, I.; Ruggieri, F.; Mazzeo, P. In utero exposure to di-(2-ethylhexyl) phthalate and duration of human pregnancy. Environ. Health Perspect. 2003, 111, 1783–1785. [Google Scholar] [CrossRef]
- Bekö, G.; Weschler, C.J.; Langer, S.; Callesen, M.; Toftum, J.; Clausen, G. Children’s phthalate intakes and resultant cumulative exposures estimated from urine compared with estimates from dust ingestion, inhalation and dermal absorption in their homes and daycare centers. PLoS ONE 2013, 8, e62442. [Google Scholar] [CrossRef]
- Kensara, O.A.; Wootton, S.A.; Phillips, D.I.; Patel, M.; Jackson, A.A.; Elia, M.; Hertfordshire Study Group. Fetal programming of body composition: Relation between birth weight and body composition measured with dual-energy x-ray absorptiometry and anthropometric methods in older Englishmen. Am. J. Clin. Nutr. 2005, 82, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lin, L.; Cao, Y.; Chen, B.; Zheng, L.; Ge, R.S. Phthalate levels and low birth weight: A nested case-control study of Chinese newborns. J. Pediatr. 2009, 155, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, J.; Garcia, J.M.; Lin, H.; Wang, Y.; Yan, P.; Wang, L.; Tan, Y.; Luo, J.; Qiu, Z.; et al. Phthalate levels in cord blood are associated with preterm delivery and fetal growth parameters in Chinese women. PLoS ONE 2014, 9, e87430. [Google Scholar] [CrossRef]
- Strømmen, K.; Lyche, J.L.; Blakstad, E.W.; Moltu, S.J.; Veierød, M.B.; Almaas, A.N.; Sakhi, A.K.; Thomsen, C.; Nakstad, B.; Brække, K.; et al. Increased levels of phthalates in very low birth weight infants with septicemia and bronchopulmonary dysplasia. Environ. Int. 2016, 89–90, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Rekiel, A.; Więcek, J.; Batorska, M.; Kulisiewicz, J. Effect of piglet birth weight on carcass muscle and fat content and pork quality—A Review. Ann. Anim. Sci. 2015, 15, 271–287. [Google Scholar] [CrossRef]
- Chang, E.I.; Rozance, P.J.; Wesolowski, S.R.; Nguyen, L.M.; Shaw, S.C.; Sclafani, R.A.; Bjorkman, K.K.; Peter, A.K.; Hay, W., Jr.; Brown, L.D. Rates of myogenesis and myofiber numbers are reduced in late gestation IUGR fetal sheep. J. Endocrinol. 2019, 244, 339–352. [Google Scholar] [CrossRef]
- Hansen, N.S.; Hjort, L.; Broholm, C.; Gillberg, L.; Schrölkamp, M.; Schultz, H.S.; Mortensen, B.; Jørgensen, S.W.; Friedrichsen, M.; Wojtaszewski, J.F.; et al. Metabolic and transcriptional changes in cultured muscle stem cells from low birth weight subjects. J. Clin. Endocrinol. Metab. 2016, 101, 2254–2264. [Google Scholar] [CrossRef]
- Chen, S.S.; Hung, H.T.; Chen, T.J.; Hung, H.S.; Wang, D.C. Di-(2-ethylhexyl)-phthalate reduces MyoD and myogenin expression and inhibits myogenic differentiation in C2C12 cells. J. Toxicol. Sci. 2013, 38, 783–791. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Yen, Y.P.; Tsai, K.S.; Yang, R.S.; Liu, S.H. Low-dose benzo(a)pyrene and its epoxide metabolite inhibit myogenic differentiation in human skeletal muscle-derived progenitor cells. Toxicol. Sci. 2014, 138, 344–353. [Google Scholar] [CrossRef]
- Metterlein, T.; Hoffmann, P.; Späth, R.; Gruber, M.; Graf, B.M.; Zink, W. In vitro myotoxic effects of bupivacaine on rhabdomyosarcoma cells, immortalized and primary muscle cells. Cancer Cell Int. 2015, 15, 75. [Google Scholar] [CrossRef]
- Nouette-Gaulain, K.; Bellance, N.; Prévost, B.; Passerieux, E.; Pertuiset, C.; Galbes, O.; Smolkova, K.; Masson, F.; Miraux, S.; Delage, J.P.; et al. Erythropoietin protects against local anesthetic myotoxicity during continuous regional analgesia. Anesthesiology 2009, 110, 648–659. [Google Scholar] [CrossRef] [Green Version]
- Yen, Y.P.; Tsai, K.S.; Chen, Y.W.; Huang, C.F.; Yangm, R.S.; Liu, S.H. Arsenic inhibits myogenic differentiation and muscle regeneration. Environ. Health Perspect. 2010, 118, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.P.; Tsai, K.S.; Chen, Y.W.; Huang, C.F.; Yang, R.S.; Liu, S.H. Arsenic induces apoptosis in myoblasts through a reactive oxygen species-induced endoplasmic reticulum stress and mitochondrial dysfunction pathway. Arch. Toxicol. 2012, 86, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Asakura, A.; Seale, P.; Girgis-Gabardo, A.; Rudnicki, M.A. Myogenic specification of side population cells in skeletal muscle. J. Cell. Biol. 2002, 159, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Shainberg, A.; Yagil, G.; Yaffe, D. Alterations of enzymatic activities during muscle differentiation in vitro. Dev. Biol. 1971, 25, 1–29. [Google Scholar] [CrossRef]
- Chen, H.J.; Wang, C.C.; Chan, D.C.; Chiu, C.Y.; Yang, R.S.; Liu, S.H. Adverse effects of acrolein, a ubiquitous environmental toxicant, on muscle regeneration and mass. J. Cachexia Sarcopenia Muscle 2019, 10, 165–176. [Google Scholar] [CrossRef]
- Kaliman, P.; Viñals, F.; Testar, X.; Palacín, M.; Zorzano, A. Phosphatidylinositol 3-kinase inhibitors block differentiation of skeletal muscle cells. J. Biol. Chem. 1996, 271, 19146–19151. [Google Scholar] [CrossRef]
- Otake, T.; Yoshinaga, J.; Yanagisawa, Y. Exposure to phthalate esters from indoor environment. J. Expo. Anal. Environ. Epidemiol. 2004, 14, 524–528. [Google Scholar] [CrossRef]
- Venkataramanan, R.; Burckart, G.J.; Ptachcinski, R.J.; Blaha, R.; Logue, L.W.; Bahnson, A.; Giam, C.S.; Brady, J.E. Leaching of diethylhexyl phthalate from polyvinyl chloride bags into intravenous cyclosporine solution. Am. J. Hosp. Pharm. 1986, 43, 2800–2802. [Google Scholar] [CrossRef]
- Karle, V.A.; Short, B.L.; Martin, G.R.; Bulas, D.I.; Getson, P.R.; Luban, N.L.; O’Brien, A.M.; Rubin, R.J. Extracorporeal membrane oxygenation exposes infants to the plasticizer, di(2-ethylhexyl) phthalate. Crit. Care Med. 1997, 25, 696–703. [Google Scholar] [CrossRef]
- Sjöberg, P.O.; Bondesson, U.G.; Sedin, E.G.; Gustafsson, J.P. Exposure of newborn infants to plasticizers. Plasma levels of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate during exchange transfusion. Transfusion 1985, 25, 424–428. [Google Scholar] [CrossRef]
- Weintraub, H. The MyoD family and myogenesis: Redundancy, networks, and thresholds. Cell 1993, 75, 1241–1244. [Google Scholar] [CrossRef]
- Weintraub, H.; Davis, R.; Tapscott, S.; Thayer, M.; Krause, M.; Benezra, R.; Blackwell, T.K.; Turner, D.; Rupp, R.; Hollenberg, S. The myoD gene family: Nodal point during specification of the muscle cell lineage. Science 1991, 251, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Lassar, A.B.; Skapek, S.X.; Novitch, B. Regulatory mechanisms that coordinate skeletal muscle differentiation and cell cycle withdrawal. Curr. Opin. Cell Biol. 1994, 6, 788–794. [Google Scholar] [CrossRef]
- Olson, E.N.; Perry, M.; Schulz, R.A. Regulation of muscle differentiation by the MEF2 family of MADS box transcription factors. Dev. Biol. 1995, 172, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Tamir, Y.; Bengal, E. Phosphoinositide 3-kinase induces the transcriptional activity of MEF2 proteins during muscle differentiation. J. Biol. Chem. 2000, 275, 34424–34432. [Google Scholar] [CrossRef]
- Jensen, C.B.; Martin-Gronert, M.S.; Storgaard, H.; Madsbad, S.; Vaag, A.; Ozanne, S.E. Altered PI3-kinase/Akt signalling in skeletal muscle of young men with low birth weight. PLoS ONE 2008, 3, e3738. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, K.-C.; Weng, T.-I.; Chiang, W.-C.; Chiu, C.-Y.; Chan, D.-C.; Yang, R.-S.; Liu, S.-H. Plasticizer Di-(2-ethylhexyl) Phthalate and Its Metabolite Mono(2-ethylhexyl) Phthalate Inhibit Myogenesis in Differentiating Mouse and Human Skeletal Muscle Cell Models. Appl. Sci. 2022, 12, 9195. https://doi.org/10.3390/app12189195
Lan K-C, Weng T-I, Chiang W-C, Chiu C-Y, Chan D-C, Yang R-S, Liu S-H. Plasticizer Di-(2-ethylhexyl) Phthalate and Its Metabolite Mono(2-ethylhexyl) Phthalate Inhibit Myogenesis in Differentiating Mouse and Human Skeletal Muscle Cell Models. Applied Sciences. 2022; 12(18):9195. https://doi.org/10.3390/app12189195
Chicago/Turabian StyleLan, Kuo-Cheng, Te-I Weng, Wei-Che Chiang, Chen-Yuan Chiu, Ding-Cheng Chan, Rong-Sen Yang, and Shing-Hwa Liu. 2022. "Plasticizer Di-(2-ethylhexyl) Phthalate and Its Metabolite Mono(2-ethylhexyl) Phthalate Inhibit Myogenesis in Differentiating Mouse and Human Skeletal Muscle Cell Models" Applied Sciences 12, no. 18: 9195. https://doi.org/10.3390/app12189195