Abstract
Human lysozyme is a natural non-specific immune protein that participates in the immune response of infants against bacterial and viral infections. Lysozyme is a well-known hydrolase that cleaves peptidoglycan in bacterial cell walls. Several crystal structures of human lysozyme have been reported, but little is known regarding how it recognizes sugar molecules. In this study, the crystal structures of human lysozyme in its native and two N-acetyl-α-d-glucosamine (α-D-NAG)-bound forms were determined at 1.3 Å and 1.55/1.60 Å resolution, respectively. Human lysozyme formed a typical c-type lysozyme fold and the α-D-NAG molecule was bound to the middle of subsites C and D. The N-acetyl and glucosamine groups of α-D-NAG were stabilized by hydrophobic interactions (Val117, Ala126, and Trp127), hydrogen bonds (Asn64, Asn78, Ala126, and Val128), and water bridges. Conformational changes of Arg80, Tyr81, Val128, and Arg131 of human lysozyme were observed due to the interactions of α-D-NAG with the active-site cleft. The binding configuration of α-D-NAG in human lysozyme was distinct compared with that of other sugar-bound lysozymes. Findings from this structural analysis provide a better understanding of the sugar recognition of human lysozyme during the immune response to microbial pathogens.
1. Introduction
In humans, lysozyme (muramidase or N-acetylmuramide glycanhydrolase) is found in several organs, tissues, and secretions. It is also found in the organs and secretions of other vertebrates, invertebrates, plants, and bacteria [1,2]. This enzyme is functionally conserved and exhibits antimicrobial activity that is critical to host defense [3]. Lysozymes are classified into three main families based on their amino acid sequence and biochemical properties. These families include the chicken or conventional type (c-type), goose type (g-type), and invertebrate type (i-type) [2,3]. In addition, phage type, bacterial type, and plant type lysozymes are also found in nature [4,5,6].
Bacterial cell walls are composed predominantly of peptidoglycan layers. Peptidoglycans consists of alternating units of N-acetylglucosamine (NAG or GlcNAc) and N-acetylmuramic acid (NAM or MurNAc). The peptide stems are covalently bound to a lactyl group in the NAM moiety [3]. Lysozymes enzymatically cleaves the peptidoglycan in bacterial cell walls by catalyzing the hydrolysis of the β-(1,4) linkages between the NAG and NAM saccharides [7]. These linkages are critical for the resistance of bacteria to osmotic stress, and their hydrolysis results in the loss of cellular membrane integrity and ultimately cell death by lysis [3,8,9]. Due to its broad antibacterial activity against pathogenic bacteria and viruses, lysozyme has been widely used in food, pharmaceutical, and clinical treatments, including various oral health care and antibiotic treatments [10,11].
In humans, lysozyme is highly expressed in breast milk and acts as an important natural non-specific immune protein that participates in the immune response of infants to bacterial [12]. For example, the concentration of lysozyme in the airway surface liquid of humans is estimated to be 20–100 µg/mL, which is sufficient to kill important lung pathogens, such as Gram-negative Pseudomonas aeruginosa and Gram-positive Staphylococcus aureus [13]. Lysozyme enhances the immune system [14,15], and in addition to antibacterial activity, it may also exhibit antitumor [16] and antiviral [17] activities. However, the molecular mechanism of these activities remains unclear.
Many crystal structures of human lysozyme (hLZM) have been reported, but most crystal structures of hLZM have been used as model samples in X-ray crystallographic studies. For instance, only one crystal structure of saccharide-bound hLZM (PDB ID: 5LSH, tetrasaccharide fragment from the O-chain of lipopolysaccharide from Klebsiella pneumonia) [15] has been deposited in the Protein Data Bank (PDB). Accordingly, how hLZM recognizes various types of saccharide molecules has not been fully elucidated. On the other hand, the crystal structure of hLZM complexed with hexa-N-acetyl-chitohexaose has previously been reported [18], but the coordinates and structural factors for this crystal structure are not deposited in PDB. Although the substrate recognition site of hLZM has been studied, it is not known how it recognizes single NAG or NAM sugar molecules constituting the substrate. The observation of single sugar molecule recognition of hLZM is expected to contribute to the initial first sugar molecule recognition when hLZM recognizes the actual substrate. In addition, it is unclear how hLZMs recognize sugars in bacterial cells and to what degree they are evolutionarily conserved with other lysozymes.
To better understand saccharide recognition of hLZM, the crystal structures of native hLZM and hLZM complexed with N-acetyl-α-d-glucosamine (α-D-NAG) were determined at 1.30 Å and 1.55/1.60 Å resolution, respectively. The α-D-NAG molecule was bound to the middle of subsites C and D in the active-site cleft of hLZM. The α-D-NAG molecule was stabilized by conserved hydrophobic interactions, hydrogen bonds, and water bridges. The α-D-NAG binding configuration of hLZM is unique when compared with crystal structure of sugar-bound lysozyme. Results from this study will provide a better understanding of saccharide recognition by hLZM and insight into the human immune response associated with hLZM.
2. Materials and Methods
2.1. Sample Preparation
The hLZM (cat. no. L1667) and N-acetyl-d-glucosamine (cat. no. A4106) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The hLZM was reconstituted in a solution containing 10 mM Tris-HCl, pH 8.0, and 200 mM NaCl. Protein crystallization was performed using the batch method as previously described [19]. Equal volumes (20 μL each) of lysozyme (50 mg/mL) and crystallization solution containing 0.1 M Na-acetate (pH 4.5), 5% (w/v) PEG 6000, and 1 M NaCl were transferred into a 1.5-mL microtube and immediately vortexed for 30 s. The sample mixture was incubated at 4 °C. The protein crystals were grown under protein precipitation for two weeks. The crystal dimensions were 0.25 × 0.25 × 0.10 mm3.
2.2. X-ray Diffraction Data Collection
The diffraction data were collected at Beamline 7A of the Pohang Accelerator Laboratory (Pohang, Korea). For native data collection, a single crystal of hLZM was soaked for 10 s in cryo-protectant solution (crystallization solution and 20% [v/v] ethylene glycol). The crystal was then mounted on the goniometer under a nitrogen gas stream at 100 K. For the hLZM-NAG complex, a single hLZM crystal was soaked for 30 min in a solution containing 0.1 M Na-acetate (pH 4.5), 5% (w/v) PEG 6000, 1 M NaCl, and 50 mM NAG. This crystal was then further soaked in a cryo-protectant solution and the diffraction data were collected. Indexing, integration, and processing of the diffraction data were performed using the HKL2000 program [20].
2.3. Structure Determination
The phases were obtained using the molecular replacement method with the MOLREP program [21]. The crystal structure of the chicken egg-white lysozyme (PDB ID: 7E02, sequence identity: 59%) [22] was used as the search model. The model was built using the program Coot [23]. Model refinement was performed using refine.phenix of the PHENIX program [24]. Data collection and refinement statistics are presented in Table 1. The geometry of the final models was validated using MolProbity [25]. The protein–ligand interaction was analyzed using PLIP [26]. Structure-based sequence alignments were generated using Clustal-Omega [27] and ESPript [28]. The structure was visualized using PyMOL (http://pymol.org).

Table 1.
Data collection and refinement statistics.
3. Results and Discussion
3.1. Overall Structure
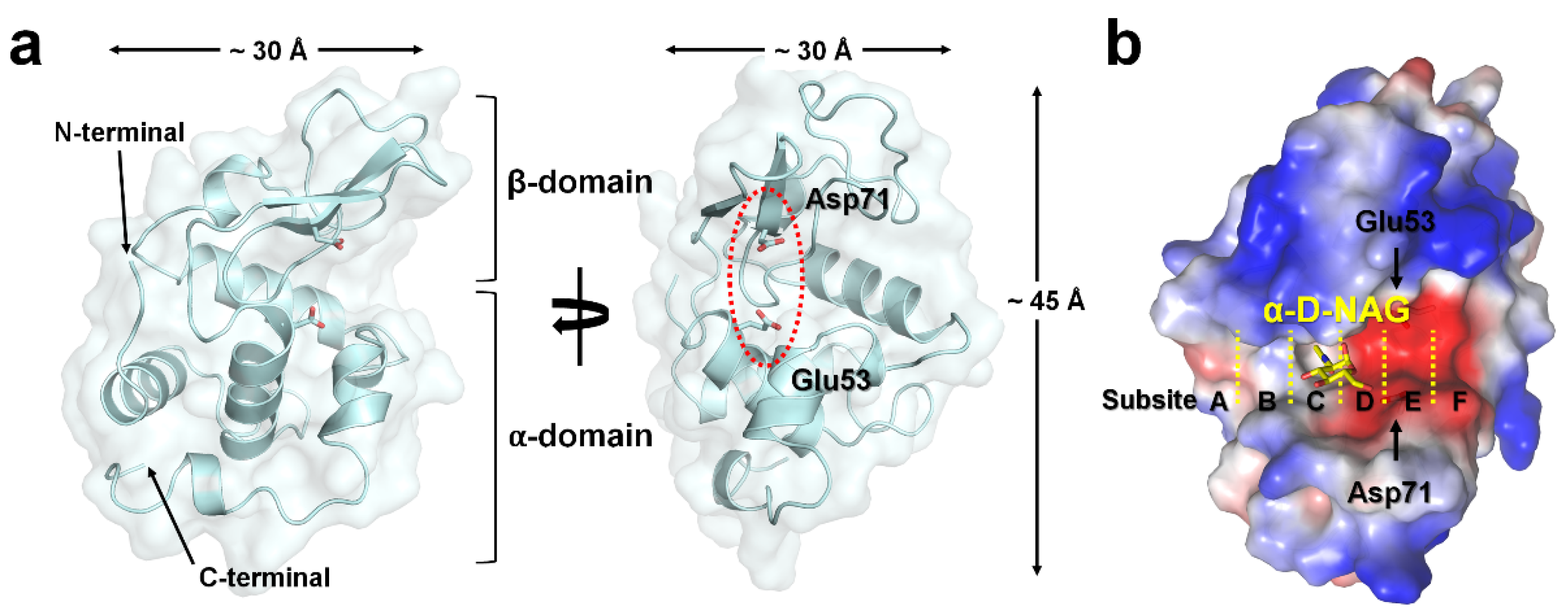
All hLZM crystals belonged to the orthorhombic space group P212121, occupying one molecule in an asymmetric unit. The crystal structures of a native and two NAG-bound hLZM crystals were determined at 1.30 and 1.55/1.60 Å resolution, respectively. The electron density was very clear for most of the residues between Lys19 and Val148 of all hLZM proteins, but not for Arg59–Ala60 of the native hLYZ. Validation of the geometry showed that >96% of the hLZM residues were within the most favored region of the Ramachandran plot and without outlier residues (Table 1). As hLZM contains eight cysteine residues that form disulfide bonds (Cys24–Cys146, Cys48–Cys134, Cys83–Cys99, and Cys95–Cys113), the lack of a significant negative Fo-Fc electron-density map indicated that all the hLZM structures determined in the current study were in an oxidized state. The hLZM was composed of seven α-helices and two β-strands, forming one α-domain and one β-domain (Figure 1a). The approximate dimensions of hLZM were 30 × 30 × 45 Å (Figure 1a). The active site of hLZM can accommodate six consecutive sugars at subsites A–F [2]. The catalytic residues Glu53 and Asp71 cleave the bond between subsites D and E (Figure 1b). The electrostatics of the active site of hLZM revealed a strong negatively charged surface (Figure 1b). The acetate ion of native hLZM was observed at subsite C and the α-D-NAG molecule of the hLZM-NAG complex was observed in the middle of subsites C and D (see below).
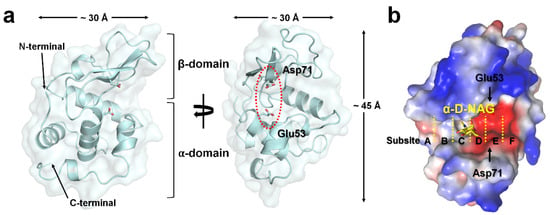
Figure 1.
Crystal structure of hLZM. (a) Cartoon and surface representation of hLZM. (b) Electrostatistics of the hLZM-NAG surface. Subsites in the active-site cleft of hLZM are indicated by the yellow dotted lines.
3.2. NAG Binding
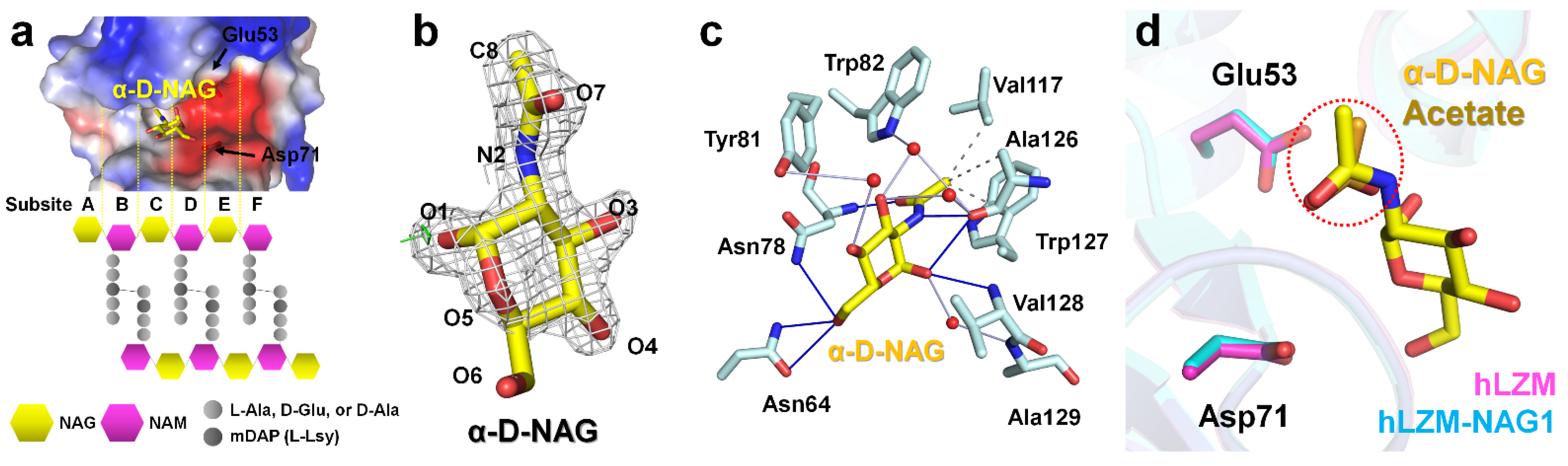
To investigate the hLZM recognition of the peptidoglycan component, NAG ligand-soaking experiments were performed with hLZM and two crystal structures of NAG-bound hLZM were determined. In the previous peptidoglycan cleavage model, NAG units of peptidoglycan were located at subsites A, C, and E, whereas NAM units were positioned at subsites B, D, and F (Figure 2a). Crystallographic results of the NAG-soaking experiment revealed the electron-density map corresponding NAG was only observed at the middle of the subsites C and D of hLZM (Figure 2b), with the other subsites being occupied by water molecules. The NAG reagent used in this experiment was a mixture of N-acetyl-α-d-glucosamine (α-D-NAG) and the N-acetyl-β-d-glucosamine (β-D-NAG). Analysis of the refinement results and electron-density map showed that α-D-NAG was bound to hLZM (Supplementary Figure S1). This indicated that hLZM preferred the interaction with α-D-NAG compared with that of β-D-NAG when a single NAG molecule was interacting hLZM. To confirm that the NAG molecule reproducibly binds to hLZM, two diffraction data sets of the hLZM crystal soaked with NAG were collected. In the two crystal structures of α-D-NAG-bound hLZM, the quality of the electron density for α-D-NAG and the binding site was almost identical, while the quality of the electron-density map for α-D-NAG surrounding water molecules was slightly different (Supplementary Figure S2). The superimposition of the two α-D-NAG -bound structures showed that the position of α-D-NAG was almost identical with the root-mean-square deviation (RMSD) analysis of 0.141 Å (Supplementary Figure S3). This indicated that a single α-D-NAG molecule was specifically bound in the middle of subsites C and D in hLZM. The binding of α-D-NAG in the two α-D-NAG-bound structures was analyzed using the crystal structure of hLZM-NAG1, which was superior to that of hLZM-NAG2 in terms of data quality and electron density clarity.
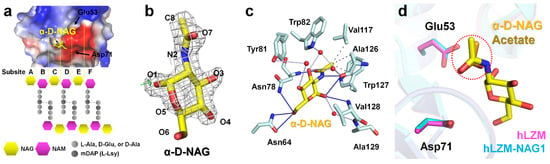
Figure 2.
NAG binding to the active site of lysozyme. (a) Active-site cleft of hLZM and schematic representation of peptidoglycan. In peptidoglycan, N-acetylmuramic acid (NAM) is covalently linked to a peptide chain (2–5 amino acid), including a dibasic amino acid [typically meso-diaminopimelic acid (mDAP) or l-lysine]. (b) 2mFo-DFc (1.2 σ, grey mesh) and mFo-DFc (+3 σ: green mesh, −3 σ: red mesh) electron-density maps of α-D-NAG in hLZM-NAG. (c) Interaction between α-D-NAG and hLZM. Hydrophobic interactions, hydrogen bonds, and water bridges are indicated by grey dotted-lines, blue lines, and light blue lines, respectively. (d) Superimposition of native hLZM (magenta) and hLZM-NAG (cyan). The N-acetyl group of α-D-NAG (yellow) and acetate ion (orange) are indicated by the red dots.
The electron-density map of N-acetyl and the ring of glucosamine were clearly observed in hLZM-NAG1, whereas the electron-density map of the C6-O6 region of glucosamine was weaker. The α-D-NAG molecule was stabilized by hydrophobic interactions (Val117, Ala126, and Trp127), hydrogen bonds (Asn64, Asn78, Ala126, and Val128), and water bridges (Tyr81, Trp82, Ala126, and Ala129). Specifically, the N-acetyl group of α-D-NAG was located at the main chains between residues Asn78 and Ala128 (Figure 2c). The C8 atom of the N-acetyl group of α-D-NAG interacted with the CG1 atom of Val117, CB atom of Ala126, and CD2 atom of Trp127 by hydrophobic interactions at distances of 3.60, 3.97, and 3.70 Å, respectively. The O7 and N2 atoms of the N-acetyl group of α-D-NAG interacted with the amine group of Asn78 and the carbonyl group of Ala126 at distances of 2.92 Å and 3.16 Å, respectively. The glucosamine group of α-D-NAG was located at the side chains between residues Asn78 and Val128 (Figure 2c). The O1 atom of the glucosamine of α-D-NAG interacted with amino group of Val128 at a distance of 3.27 Å. The distance between the side chain of Asn78 and ring center of glucosamine of α-D-NAG was 4.53 Å. In addition, five water bridges between α-D-NAG and hLZM were observed at distances of 2.79–4.02 Å. More detail information regarding the distances between hLZM and α-D-NAG are listed in Table 2.
Table 2.
Interaction distances between hLZM and α-D-NAG.
The electron-density map corresponding to the acetate ion of native hLZM was observed at subsite C (Supplementary Figure S4). The oxygen atom interacted with the amino group of Asn78 at a distance of 2.90 Å (Supplementary Figure S5). The methyl group of acetate was stabilized by hydrophobic interactions with Val117 and Ala126 (Supplementary Figure S5), showing similarities with the binding mode of the N-acetyl group of α-D-NAG in hLZM-NAG. Superimposition of the native hLZM and hLZM-NAG structures showed that the position of the acetate ion in native hLZM was similar to that of the N-acetyl group of α-D-NAG in hLZM-NAG (Figure 2d).
3.3. Comparison of Native hLZM and NAG-Bound hLZM
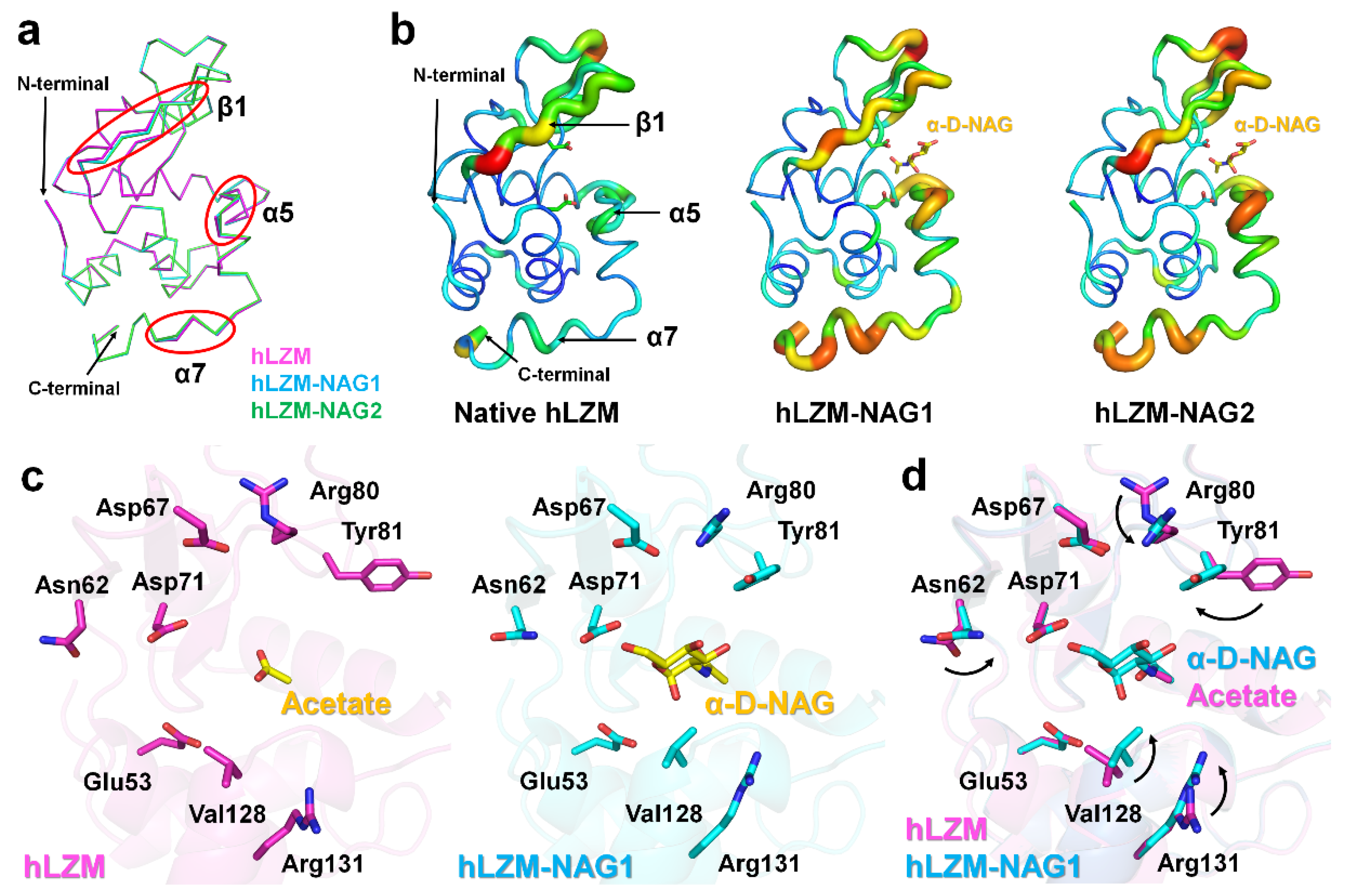
To investigate how hLZM recognizes the α-D-NAG molecule, the structures of native hLZM and hLZM-NAG were compared. Superimposition of their crystal structures showed a high level of structural similarity with RMSD of 0.093–0.128 Å, indicating there was no large conformational changes upon α-D-NAG binding to hLZM; however, there was a small shift of the main-chain of the β1-strand and C-terminal helix (Figure 3a). The B-factor representation revealed that the β1-strand, α5-helix, and C-terminal α7-helix of α-D-NAG-bound hLZMs were more flexible compared with that of native hLZM (Figure 3b). Structural analysis of ligand binding revealed conformation changes of Asn62, Arg80, Tyr81, Val128, and Arg131 of hLZM upon α-D-NAG binding (Figure 3c). In particular, the two methyl groups of the side chain of Val128 of NAG-bound hLZMs were directed more toward the α-D-NAG molecule compared with that in native hLZM. The side chain of Val110 was separated from the ring center of glucosamine of α-D-NAG by 0.2 Å, indicating that α-D-NAG-binding induced conformational changes of the Val128 side chain. The side chain of Tyr81 in native hLZM was positioned toward subsite E, but the side chain of Tyr81 of α-D-NAG-bound hLZM was positioned toward the α-D-NAG molecule at subsite D, due to rotation of the side change by approximately 106° (Figure 3d). The Tyr81 residue of α-D-NAG-bound hLZM interacted with α-D-NAG via a water bridge (Figure 2c). The smallest distance between the glucosamine of α-D-NAG and phenol ring of Tyr81 was approximately 5 Å, whereas the distance between the ring center of glucosamine of α-D-NAG and the phenol ring of Tyr81 was approximately 7 Å in parallel dispatch. On the other hand, the CZ atoms of Arg80 and Arg131 were separated from α-D-NAG by 6.40 Å and 6.65 Å, respectively, resulting in no direct interaction between these atoms and α-D-NAG. It was considered that the sidechain shifts of Arg80 and Arg131 may have been induced by conformational changes of neighboring residues Tyr81 and Val128. Meanwhile, no significant conformational changes of active-site residues Glu53 or Asp71 were observed. Taken together, this suggests that amino acids present in the active-site cleft undergo conformational changes and stabilize α-D-NAG when α-D-NAG binds to hLZM.
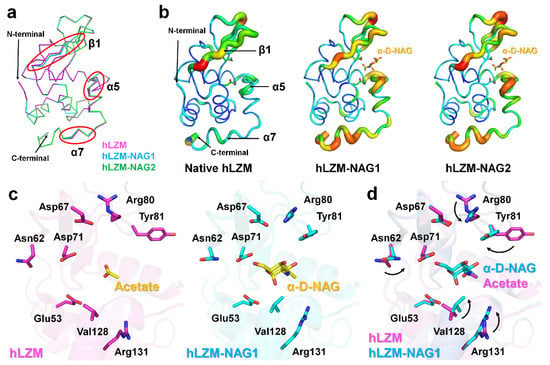
Figure 3.
Comparison of native hLZM and α-D-NAG-bound hLZM. (a) Superimposition of the main chains of native hLZM (magenta), hLZM-NAG1 (cyan), and hLZM-NAG2 (green). (b) B-factor putty representation of native hLZM and α-D-NAG-bound hLZMs. (c) Ligand binding in native hLZM and hLZM-NAG1. (d) Superimposition of acetate and α-D-NAG binding from native hLZM and α-D-NAG-bound hLZM.
3.4. Structural Comparison of hLZM with Other Lysozyme Structures
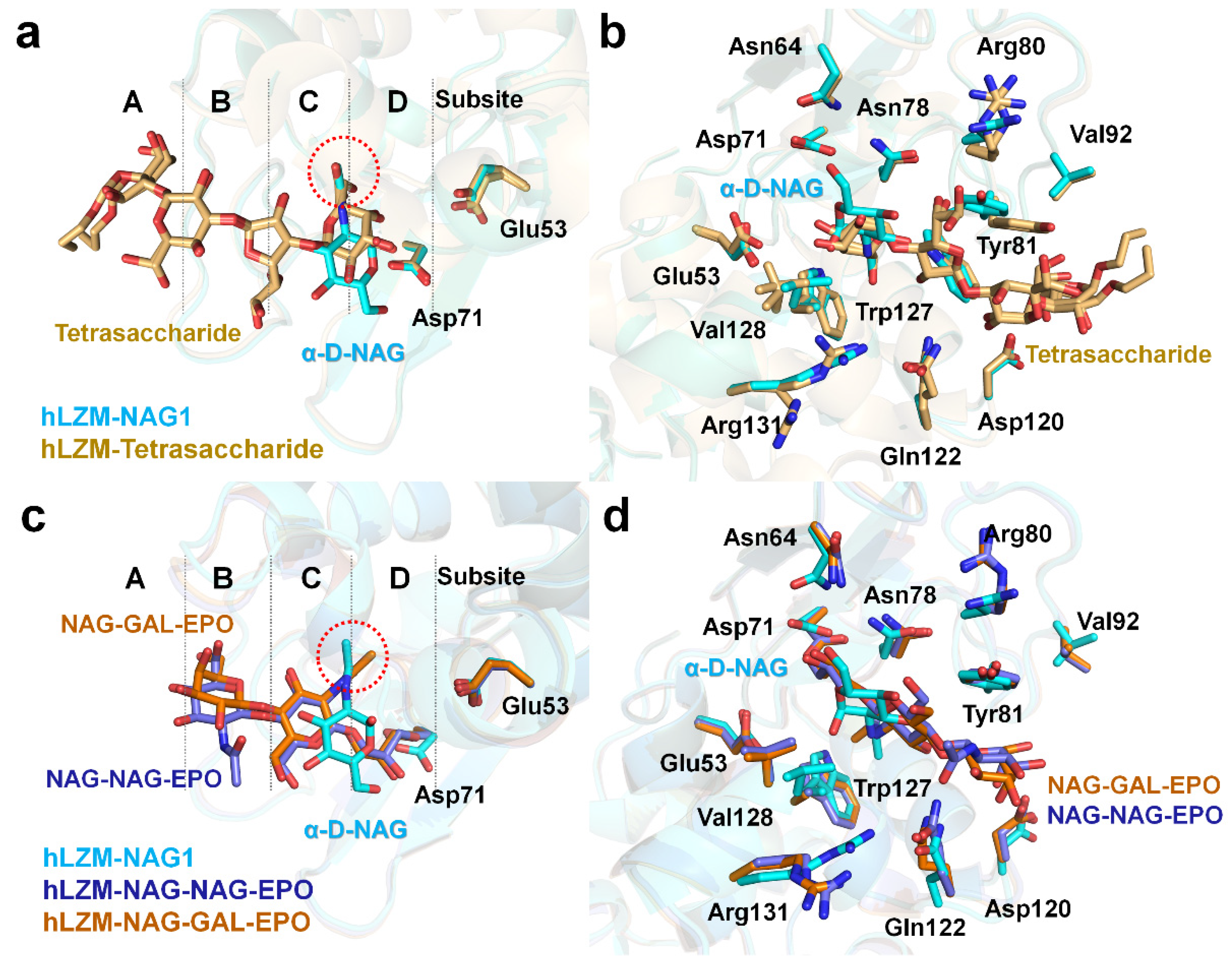
To better understand the saccharide recognition of hLZM, the crystal structure of hLZM-NAG was compared with that of a previously reported crystal structure of hLZM complexed with a tetrasaccharide fragment of the O-chain of lipopolysaccharide from Klebsiella pneumonia (PDB ID: 5LSH). In this crystal structure, tetrasaccharide bound to the substrate-binding cleft of hLZM with two different conformations (Figure 4a), in which tetrasaccharide was located at the subsites A–D of hLZM. The superimposition of α-D-NAG-bound and tetrasaccharide-bound hLZM showed a high degree of structural similarity with RMSD of 0.216 Å, and two O6 atoms of tetrasaccharide demonstrated positional similarity with that of the N-acetyl group of α-D-NAG (Figure 4a). Meanwhile, the ring positions of α-D-NAG and tetrasaccharide were different due to differences in chemical structure (Figure 4a). Superimposition of these hLZM structures showed that most of the amino acids interacting with saccharide molecules underwent large conformational changes of their side chains on the active-site cleft of hLZM (Figure 4b). Residues Tyr81 and Val128, in particular, demonstrated large differences in side-chain conformation.
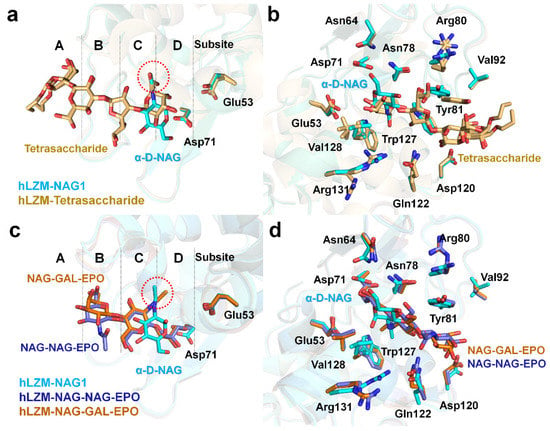
Figure 4.
Comparison of α-D-NAG-bound and tetrasaccharide-bound hLZM. (a) Superimposition of hLZM-NAG (cyan) with a tetrasaccharide fragment from the O-chain of lipopolysaccharide from Klebsiella pneumonia (orange; PDB ID: 5LSH). The O6 atoms of tetrasaccharide and the N-acetyl group of α-D-NAG are indicated by red dots. (b) Superimposition of saccharide residues interacting with hLZM complexed with α-D-NAG (cyan) and the tetrasaccharide fragment (orange). (c) Superimposition of hLZM-NAG (cyan) with hLZM complexed with NAG-NAG-EPO (blue; PDB ID: 1REY) and NAG-GAL-EPO (brown; PDB ID: 1REZ). The N-acetyl groups of α-D-NAG, NAG-NAG-EPO, and NAG-GAL-EPO are indicated by red dots. (d) Superimposition of saccharide residues interacting with hLZM complexed with α-D-NAG (cyan), NAG-NAG-EPO (blue), and NAG-GAL-EPO (brown).
Subsequently, the crystal structure of hLZM-NAG was compared with those of hLZM complexed with 2′,3′-epoxypropyl α-glycoside derivatives of the disaccharides [NAG-NAG-EPO (PDB ID: 1REY) or GAL-NAG-EPO (PDB ID: 1REZ)]. In these complex structures, the NAG molecule is located on subsite C (Figure 4c), which coincides with the substrate binding site model. The second sugar of the two ligands (either NAG or GAL) is located on subsite B, but with a slightly different conformation (Figure 4c). This indicates that the sugar recognition mode of hLZM depends on the number and type of sugar. Superimposition of these structures showed that, in the active-site cleft of hLZM, most amino acids interacting with saccharide molecules underwent small conformational changes of their side chains (Figure 4d).
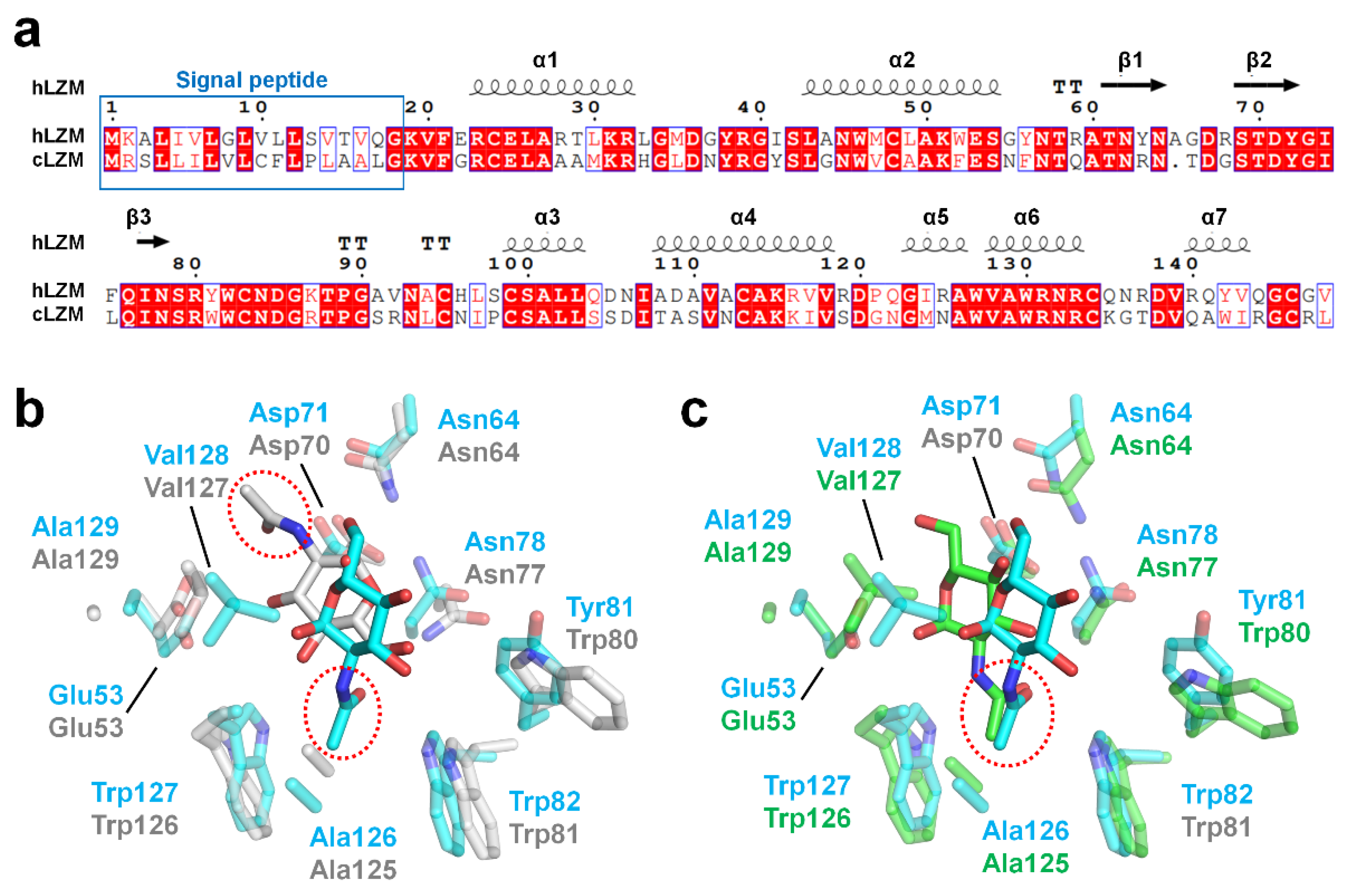
To evaluate the degree of conservation in the binding mode of α-D-NAG among the c-type lysozyme family, the crystal structure of hLZM-NAG was compared with that of the crystal structure of α-D-NAG-bound chicken egg-white lysozyme (cLZM), which is a well-studied model structure of c-type lysozyme. The antibacterial activity of hLZM is three-fold higher than that of the antibacterial activity of cLZM [1]. The sequence identity between hLZM and cLZM was 59% (Figure 5a). To date, three α-D-NAG-bound cLZM have been deposited in PDB. One of the crystal structures of cLZM (PDB ID: 1JA7) was determined in a powder diffraction study [29], while the other two crystal structure of cLZM (PDB ID: 7BHM and 7BHN) were determined by serial crystallography performed at room temperature [30]. In the case of the latter two crystal structures of α-D-NAG-bound cLZM, the results are highly similar due to only differences in the NAG mixing times being used. Accordingly, only one of these crystal structures of α-D-NAG-bound cLZM (7BHN) was compared in this study (the one that underwent the longer NAG-mixing time). Superimposition of the α-D-NAG-bound hLZM with that of the α-D-NAG-bound cLZM-1JA7 and cLZM-7BHN had an RMSD of 1.279 Å and 0.475 Å, respectively, for the main chain of hLZM. The glucosamine group of α-D-NAG was positioned toward to the active-site residues Asp35 and Glu71 in hLZM, whereas the N-acetyl group of α-D-NAG was positioned toward to active-site residues in cLZM-1JA7 (Figure 5b). As a result, a similarity in the α-D-NAG-binding modes for hLZM and cLZM-1JA7 was not observed. In contrast, in cLZM-7BHN, the glucosamine group of α-D-NAG was positioned toward the active site of cLZM, similar to that of α-D-NAG-bound hLZM (Figure 5c). Moreover, the binding mode of the N-acetyl group of α-D-NAG was similar between hLZM and cLZM (Figure 5c). Meanwhile, the position of the O6 atom in the glucosamine group of α-D-NAG from hLZM and cLZM differed by 3.95 Å.
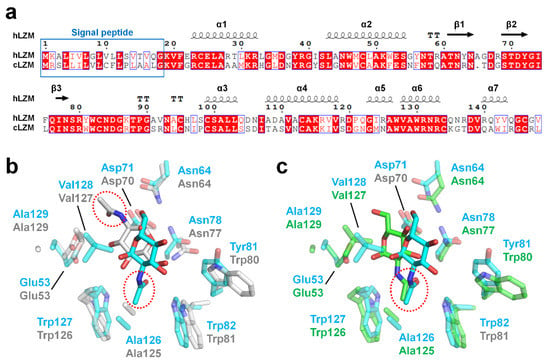
Figure 5.
Comparison of hLZM and cLZM. (a) Sequence alignment of hLZM (Uniprot: P61626) and cLZM (P00698). (b) Superimposition of the crystal structures of α-D-NAG-bound hLZM with cLZM (PDB ID: 1JA7), which was determined through a powder diffraction study. (c) Superimposition of the α-D-NAG-binding sites of hLZM and cLZM (PDB ID: 7BHN), which was determined through serial femtosecond crystallography.
4. Conclusions
The hydrolases hLZM plays a significant role in the antibacterial and antiviral immune responses in human infants. In this study, the high-resolution crystal structure of hLZM complexed with N-acetyl-α-d-glucosamine, a component of peptidoglycan, was reported for the first time. Six subsites for sugar binding were identified in the cleft of the active site of hLZM, with the α-D-NAG molecule being only observed in the middle of subsites C and D. This result indicates that a single α-D-NAG molecule prefers to bind near the active site of hLZM. Although the binding mode of α-D-NAG to hLZM is not fully consistent with that of other hLZM-tetrasaccharide or cLZM-α-D-NAG binding modes, the position of saccharide binding was similar. This indicates that the specific NAG binding to hLZM and cLZM are distinct, but the subsites C and D are positionally important for the recognition of a single NAG molecule. Meanwhile, the previous peptidoglycan binding and cleavage model of hLZM suggested that the NAG molecule may prefers to interact with subsites A, C, or F; however, the current actual experimental results located α-D-NAG only in the middle of subsites C and D. This indicates that the modes of binding of a single α-D-NAG molecule compared with that of the actual substrate molecule consisting of NAG-NAM may differ. Accordingly, further structural and biochemical experiments will be required to identify the interactions between hLZM and their substrates. Nevertheless, this study provided a greater understanding of how hLZM recognizes single α-D-NAG molecules, which is a component of peptidoglycan, and how it differentiates it from other sugar-binding LZM structures. The results from this study will provide new insights into sugar recognition of hLZM and the role hLZM plays in the immune system.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app12094363/s1, Figure S1: Electron-density map of hLZM after adding NAG and refinement, Figure S2: Electron-density map of hLZM-NAG1 and hLZM-NAG2, Figure S3: Superimposition of crystal structures of hLZM-NAG1 and hLZM-NAG2, Figure S4: Electron-density map of the acetate ion in native hLZM, Figure S5: Interaction between the acetate ion and hLZM.
Funding
This work was funded by the National Research Foundation of Korea (NRF) (NRF-2017M3A9F6029736 and NRF-2021R1I1A1A01050838) and the Korea Initiative for Fostering University of Research and Innovation (KIURI) Program of the NRF (NRF-2020M3H1A1075314).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The coordinates and structure factors for the protein crystals of this study are available in the Protein Data Bank (https://www.rcsb.org/) under entries 7XF6 (hLZM), 7XF7 (hLZM-NAG1), and 7XF8 (hLZM-NAG2).
Acknowledgments
I thank the staff at the 7A beamline at Pohang Accelerator Laboratory for their assistance with data collection.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef]
- Callewaert, L.; Michiels, C.W. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef]
- Bliska, J.B.; Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef]
- Jollès, P.; Jollès, J. What’s new in lysozyme research? Always a model system, today as yesterday. Mol. Cell. Biochem. 1984, 63, 165–189. [Google Scholar] [CrossRef]
- Beintema, J.J.; van Scheltinga, A.C.T. Plant lysozymes. In Lysozymes: Model Enzymes in Biochemistry and Biology; Experientia Supplementum; Springer: Berlin/Heidelberg, Germany, 1996; pp. 75–86. [Google Scholar] [CrossRef]
- Fastrez, J. Phage lysozymes. In Lysozymes: Model Enzymes in Biochemistry and Biology; Experientia Supplementum; Springer: Berlin/Heidelberg, Germany, 1996; pp. 35–64. [Google Scholar] [CrossRef]
- Uversky, V.N.; Wohlkönig, A.; Huet, J.; Looze, Y.; Wintjens, R. Structural Relationships in the Lysozyme Superfamily: Significant Evidence for Glycoside Hydrolase Signature Motifs. PLoS ONE 2010, 5, e15388. [Google Scholar] [CrossRef] [Green Version]
- Ellison, R.T.; Giehl, T.J. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef]
- Oliver, W.T.; Wells, J.E. Lysozyme as an alternative to growth promoting antibiotics in swine production. J. Anim. Sci. Biotechnol. 2015, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Tenovuo, J. Clinical applications of antimicrobial host proteins lactoperoxidase, lysozyme and lactoferrin in xerostomia: Efficacy and safety. Oral Dis. 2002, 8, 23–29. [Google Scholar] [CrossRef]
- Masschalck, B.; Michiels, C.W. Antimicrobial Properties of Lysozyme in Relation to Foodborne Vegetative Bacteria. Crit. Rev. Microbiol. 2008, 29, 191–214. [Google Scholar] [CrossRef]
- Lu, D.; Liu, S.; Ding, F.; Wang, H.; Li, J.; Li, L.; Dai, Y.; Li, N. Large-scale production of functional human lysozyme from marker-free transgenic cloned cows. Sci. Rep. 2016, 6, 22947. [Google Scholar] [CrossRef] [Green Version]
- Travis, S.M.; Conway, B.-A.D.; Zabner, J.; Smith, J.J.; Anderson, N.N.; Singh, P.K.; Peter Greenberg, E.; Welsh, M.J. Activity of Abundant Antimicrobials of the Human Airway. Am. J. Respir. Cell Mol. Biol. 1999, 20, 872–879. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Klein, P.; Morand, M.; Kiczka, W.; Studnicka, M. Immunostimulatory effects of dimerized lysozyme (KLP-602) on the nonspecific defense mechanisms and protection against furunculosis in salmonids. Vet. Immunol. Immunopathol. 1998, 61, 369–378. [Google Scholar] [CrossRef]
- Zhang, R.; Wu, L.; Eckert, T.; Burg-Roderfeld, M.; Rojas-Macias, M.A.; Lütteke, T.; Krylov, V.B.; Argunov, D.A.; Datta, A.; Markart, P.; et al. Lysozyme’s lectin-like characteristics facilitates its immune defense function. Q. Rev. Biophys. 2017, 50, e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osserman, E.F.; Klockars, M.; Halper, J.; Fischel, R.E. Effects of Lysozyme on Normal and Transformed Mammalian Cells. Nature 1973, 243, 331–335. [Google Scholar] [CrossRef]
- Lee-Huang, S.; Maiorov, V.; Huang, P.L.; Ng, A.; Lee, H.C.; Chang, Y.-T.; Kallenbach, N.; Huang, P.L.; Chen, H.-C. Structural and Functional Modeling of Human Lysozyme Reveals a Unique Nonapeptide, HL9, with Anti-HIV Activity. Biochemistry 2005, 44, 4648–4655. [Google Scholar] [CrossRef]
- Song, H.; Inaka, K.; Maenaka, K.; Matsushima, M. Structural Changes of Active Site Cleft and Different Saccharide Binding Modes in Human Lysozyme Co-crystallized with Hexa-N-acetyl-chitohexaose at pH 4.0. J. Mol. Biol. 1994, 244, 522–540. [Google Scholar] [CrossRef]
- Nam, K.H.; Cho, Y. Stable sample delivery in a viscous medium via a polyimide-based single-channel microfluidic chip for serial crystallography. J. Appl. Crystallogr. 2021, 54, 1081–1087. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef]
- Nam, K.H. Beef tallow injection matrix for serial crystallography. Sci. Rep. 2022, 12, 694. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Metoz, F. ESPript: Analysis of multiple sequence alignments in PostScript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Von Dreele, R.B. Binding of N-acetylglucosamine to chicken egg lysozyme: A powder diffraction study. Acta Crystallogr. D Biol. 2001, 57, 1836–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butryn, A.; Simon, P.S.; Aller, P.; Hinchliffe, P.; Massad, R.N.; Leen, G.; Tooke, C.L.; Bogacz, I.; Kim, I.-S.; Bhowmick, A.; et al. An on-demand, drop-on-drop method for studying enzyme catalysis by serial crystallography. Nat. Commun. 2021, 12, 4461. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).