3.1. Effect of EM and OMW Treatments on Plants Growth
In the present study, we tested the efficiency of Effective Microorganisms (EM) and Olive Mill Wastewater (OMW) on the growth, physiology, and quality parameters of costmary plants. Regarding to the growth parameters in general, the results showed that treatments with the two biostimulants had significant effects. Specifically, among assayed traits, statistically significant differences (
p ≤ 0.05) were observed respect to the type and the concentration of applied biostimulant (
Figure 1A–F;
Table 1); the treated plants showed better performances than the Control.
The ANOVA response for all growth factors detected huge differences in experimental results. The comparison of variance between and within experimental groups (rows “Model” and “Residual” in
Table 1) indicates that the variance of treatments is ever many times larger than unaccountable variance. Moreover the highly significant difference rejects for each growth features the null hypothesis that the means for the five treatments are the same (Column “PR(>F)”). Analyzing the paired results of the effects of different treatments, it appears that for all the growth variables there are always great differences between the EM, OMW and Control treatments and the Effect Size values always confirm a huge difference (
Table A2 and
Table A3). The Effect Size values tend to decrease when EM and OMW treatments at different concentrations are compared to each other. Both the
p-value values, corrected with the Bonferroni method (Column “p-corr”), confirm that for some growth features there are often no significant differences and when present, the Effect size values indicate low differences between treatments (
Table A2 and
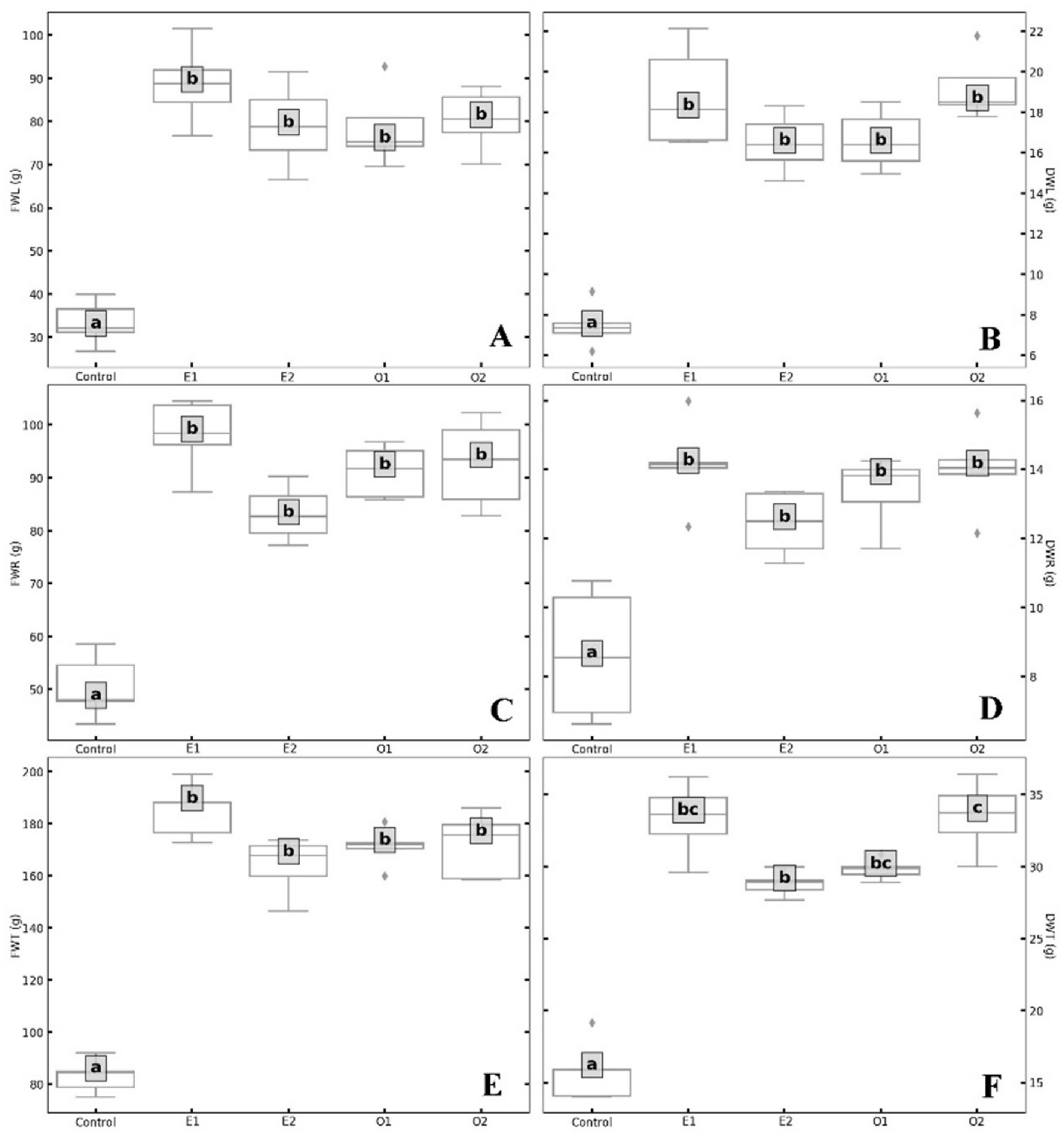
Table A3). Analysis of the different growth parameters showed statistically significant differences. The treated plants showed an average fresh leaves weight (81.6 g) that was about 48 g greater than the Control plants; and the higher value of FWL was observed in the E1 treatment with 88.7 g (
Figure 1A;
Table A4). Leaves dry weight (DWL) showed an increase from 120% (O1; E2) to about 150% (O2; E1) compared to the Control (
Figure 1B and
Table A4). The highest values of DWL were observed in E1 and O2, with 18.8 and 19.2 g respectively. Leaves fresh and dry weights showed no statistically significant differences among treatments with the two biostimulants (
Figure 1A,B,
Table A4).
Regarding the grown factors FWL and DWL, the pairwise analysis (
Table A2) showed that the practical significance, expressed in terms of Effect Size is always significant (EF < −5.3) in the comparison between Control and Treatments (E1, E2, O1, O2) and the most significant effects were recorded between Control and O2 (EF = −6.8 for FWL and EF = −7.8 for DWL) (
Table A2). Comparing the treatments in pairs, for FWL the higher value of EF was obtained for E1⇔O1 (EF = 1.01), while the minimum difference of response was obtained by comparing the groups E2⇔O1 (EF = 0.046). DWL showed a higher value of EF when comparing in pair E2⇔O2 (EF = −1.64), and the minimum difference in E2⇔O1 (EF = −0.094) (
Table A2). Concerning roots fresh and dry weights, they were significantly higher in EM and OMW treatments compared to the Control (
Figure 1C,D;
Table A4). The increase in fresh weight ranged from 65% (E2) to 94.4% (E1), and the highest value of FW roots was recorded in the E1 treatment with 98.0 g (
Figure 1C;
Table A4). For the dry weight, the best values were observed in E1 (14.1 g) and O2 (14.0 g) treatments, while the Control recorded 8.6 g (
Figure 1D). Roots fresh and dry weights showed no statistically significant differences between treatments with the two biostimulants (
Figure 1C,D). Comparing the Control with the treatments the pairwise analysis (
Table A2) showed that the maximum response to treatment for FWR was observed for pair Control⇔O1 (EF = −6.67), while among the treatments the maximum value of EF was observed for pair E1⇔E2 (2.17). Instead for DWR, the comparison between Control and treatments exhibited the maximum value for pair Control⇔E1 (EF = −3.08) and the minimum difference in Control⇔E2 (EF = −2.30). In treated plants maximum differences were observed for pair E1⇔E2 and the minimum in E1⇔O2, EF = 1.38 and EF = 0.10 respectively (
Table A2). The total fresh (FWT) and total dry weight (DWT) of OMW- and EM-treated plants were always higher in all treatments compared to Control. The treated plants showed an average FWT (172.9 g) that was about 89 g greater than the Control plants, and a higher value was observed in E1 (184.9 g) (
Table A4). The highest value in the pairwise analysis was EF = 11.4 in Control⇔O1 (
Table A2). The FWT did not show statistically significant differences among the four treatments (
Figure 1E). Regarding DWT, the treatments E1 and O2 registered the highest mean values (33.3 g and 33.5 g, respectively) and marginally significant differences were observed between the two biostimulants and the applied concentrations (
Figure 1F;
Table A4). Control in comparison to treated plants, showed the maximum value of EF (−8.07) for pair Control⇔O1, among treatments the minimum was for E1⇔O2 (EF = −0.06), and the highest was for E2⇔O2 (EF = −2.29) (
Table A2). The improved development of treated plants with EM and OMW is highlighted also in an increase in the total RGR index (
Figure 2).
At the end of the experiment (75 DTA), the total RGR of treated plants ranged from 50.5 to 53.1 mg g d, with the highest value in O2 treatment, while in the Control was 40.3 mg g d. In all plants the aerial part (leaves) exhibited the highest RGR and, in particular, the treated plants showed a significant increase of about 32% in leaves RGR compared to the Control; the highest value was observed in OMW treatment applied at the maximum concentration (O2; 63.0 mg g d).
The increased growth observed in
Tanacetum balsamita treated plants, can be attributed to the biostimulants influence and the effect is evident in both the rooting system and the aerial biomass development. In literature, several studies reported on horticultural and ornamental plants, the positive influence of microorganism activity (EM) on plant growth parameters [
36,
37,
38,
39]. In
Figure 3 the plants at T0 and T3 are shown, with details of leaves and roots.
The positive application of EM and OMW resulted also, in an increases in values of leaves number (LN) and leaf area (LA) (
Table A4;
Figure 3 and
Figure 4A,B). The average number of leaves in plants treated with OMW and EM (181.6) enhanced by 78.5% with respect to the Control; the highest values were observed applying the minor biostimulant concentrations, O1 (205 leaves) and in E1 (185.8 leaves) (
Figure 4A;
Table A4). Control plants registered a mean leaf area of 39.1 cm
while treatments determined an increase of leaf area ranging from about 72.0 cm
in E1 to 42.1 cm
in O1 (
Figure 4B;
Table A4).
Comparing the Control with the treatments the pairwise analysis (
Table A3) showed that the maximum value of EF for LN was −8.03 (Control⇔O1) and the minimum was −5.95 for Control⇔E2, however, the latter value highlights a strong response to treatment. Among the treatments, the maximum value was observed for E2⇔O1 (EF = −3.54) and the minimum was for E1⇔O2 with EF = 0.99 (
Table A3). Instead for LA, the comparison between Control and treatment exhibited the maximum value in Control⇔E1 (EF = −4.4) and the minimum difference in Control⇔O1 (EF = −0.51) (
Table A3). In treatment plants, maximum differences were observed in E1⇔O1 and the minimum in E1⇔O2, EF = 3.84, and EF = 0.69 respectively (
Table A3). Regarding these results, O1 treatment recorded a leaf area value similar to the Control, but the best result was in leaves number (205). This result is not conflicting, Hassanpouraghdam et al. [
14] had already observed an inverse relationship between leaf area and leaf number in
Tanacetum balsamita plants treated with different concentrations of N, and they considered this result to be not limiting to plant growth. The estimation of leaf number and leaf area is decisive for plant growth and development being the leaf an essential plant structure that allows gas exchange and the transformation of light energy into chemical energy useful for cellular functions [
40]. The effects of EM and OMW treatments on growth indices SLA (Specific Leaf Area), LAR (Leaf Area Ratio), and LWR (Leaf Weight Ratio), are reported in
Table A1 and
Figure 4C–E. SLA is a measure of leaf thickness, correlated with the number and/or size of leaf photosynthetic mesophyll cells [
41]. SLA index showed similar values for E2, O1, and O2 treatments and Control, while E1 treatment reported the highest value (715.2 cm
g
;
Figure 4C). LWR describes the efficiency of assimilation in the whole plant. LWR index ranged from 0.59 in O2 to 0.47 g g
in the Control and did not show differences between treated plants, which however showed values higher than the Control (
Table A4;
Figure 4D). LAR defines the leaf area produced per unit of biomass and therefore it is closely related to photosynthetic activity. In the present study LAR index values in treated plants were higher compared with the Control, and the highest data was recorded in E1 (414.9 cm
g
) followed by O2 treatment with 353.1 cm
g
(
Table A4;
Figure 4E). Comparing the Control with the treatments the pairwise analysis (
Table A3) showed that the maximum value of EF for SLA was −1.31 (Control⇔E1) and the minimum was 0.03 for Control⇔O1. Among the treatments, the maximum value was observed for E1⇔E2 (EF = 2.11) and the minimum was EF = −0.05 for E2⇔O1 (
Table A3). Instead for LWR, the comparison between Control and treatment exhibited the maximum value in Control⇔O2 (EF = −3.54) and the minimum difference in Control⇔O1 (EF = −2.20) (
Table A3). LAR showed among Control and treatment the maximum value of EF = −2.62 in Control⇔E1 and the minimum in Control⇔O1 (EF = −0.74). The highest value among treated plants was EF = 2.37 in E1⇔E2, and the lowest was in E2 ⇔O1 with EF = 0.27 (
Table A3).
In our experimental conditions, EM and OMW determined an increase in leaf number and leaf area but did not affect leaf thickness. These results agree with Scavroni et al. [
42] that used organic fertilizers in
Mentha piperita L. and with Wadas and Dziugieł [
43] for the use of biostimulants in
Solanum tuberosum L. The rise of LWR and LAR, due to the application of biostimulants, in combination with enlarged leaf size, can maximize the optical light interception efficiency and the greater acquisition of resources resulting in an increment of plant growth [
44,
45,
46].
The presence of trichomes on the leaf surface of
Tanacetum balsamita was observed, and thus effect of biostimulants on the number of trichomes was evaluated. Although trichomes have been described in some species of the genus
Tanacetum, this is the first time they are described in
Tanacetum balsamita. Costmary presented non-glandular and glandular trichomes on the adaxial and abaxial epidermis (
Figure 5A–F), even if they are both more abundant on the lower epidermis. Non-glandular trichomes are long, uniseriate T-shaped hair and consist of 2–3-foot cells and 2 acicular armed hair (
Figure 5A,B,G). Glandular trichomes consist of biseriate secretory cells and a large subcuticular cavity where secretory products are stored, like the peltate trichomes of the
Lamiaceae ([
47,
48] (
Figure 5C–E,G). Histochemical staining (Nadi Reagent) and fluorescence analysis highlighted the presence of terpenes and wall-bound hydroxycinnamates in glandular trichomes (
Figure 5D,E) [
49,
50]. Under our experimental conditions, no statistically significant difference among treatments was found in the density of glandular trichomes while in non-glandular trichomes, the number was significantly higher in the E2 (81.3) and O1 (78.7) treatments compared to the Control (64.7) (
Figure 5F–L). These results agree with the study of Saour [
51] who reported an increase in non-glandular hairs in potato plants after treatments with commercial organic biostimulant (Vitazyme™) speculating a positive relationship among the action of biostimulant, the resistance to pathogens and trichomes number.
Until now, there is no published research on the use of EM and OMW in costmary. Many studies reported a positive effect of EM on a wide range of fruits and leafy vegetables in pot and in field experiments [
52], such as apples [
53], beans [
54] and tomato [
55]. Similarly pot experiments under greenhouse, reported in
Allium cepa [
21] an increase in bulb growth parameters and in the medicinal plants
Bulbine frutescens [
22], a significant improvement in the number of new leaves, shoots, inflorescences per plant, and biomass weight. The efficiency of EM is attributed to the increase in the photosynthesis, in microbial biomass and to the improvement of water supply. EM also stimulates plant growth by improving physiological processes and plant resistance to abiotic stresses producing bioactive substances such as hormones and enzymes, accelerating the decomposition of organic materials, and controlling soil diseases [
56,
57,
58,
59]. Although most published studies show a positive effect of EM on plant growth, some researches reported negative or non-significant effects [
52], as well as in sweet corn [
60], seeds cotton [
61] and Chinese cabbage [
62]. Few information on the application of OMW on plants is available. OMW has a high content of organic compounds, and macro and microelements, making it valuable as biofertilizers and/or biostimulants. However, OMW obtained during oil production has a high level of acidity, salinity, and polyphenol content that make it harmful [
63]. Indeed, the application of undiluted OMW resulted in the suppression of seeds germination in
Triticum durum L. [
64],
Vicia faba L. and
Cicer arietinum L. [
65]. In addition, it was reported that the use of the unmodified OMW has a negative impact on physico-chemical and biological soil properties and ground waters [
65]. To make this product usable in agriculture, it is necessary to remove the detrimental products. Several methods have been developed for the improvement of OMW and the resulting product has been evaluated in different plants [
66]. In our experimental study, to reduce toxicity, OMWs were treated by a simple and easily repeatable method: decantation, dilution, and pH adjustment. Our findings are in agreement with [
23], where the filtered OMW applied to tomato plants increased growth parameters and fruit size. Other researchers have observed the same results in
Triticum aestivum var. Douma1 [
61], in
Vicia faba L. and
Cicer arietinum L. [
65], and
Phaseolus vulgaris L. [
67]. The positive effects of OMWs application are attributed to the large quantity of protein, polysaccharides, humic acids, and macro and micro mineral elements present in this compound [
23].
3.2. Mineral Composition of Leaves
The results of the mineral analysis, as tissue concentration, performed on costmary plants at 0 DAT (T0) and 75 DAT (T3) in Control, E1, E2, O1, O2, are shown in
Figure 6. The development of root, shoot, and leaves allowed more functional nutrient assimilation, and transport of mineral elements and physiological activators, as already seen by De Pascale et al. [
68].
After 75 DAT (T3), Control plants had a typical enhancement of the nutrient content (
Figure 6). The mineral element concentrations were differently affected by the type of treatment and treatment concentration. Total N showed the highest values in plants grown on E2 and O2 (28.79 ± 0.34 g Kg
DW and 30.23 ± 0.65 g Kg
DW, respectively), the highest concentrations used in this experiment. On the other hand, in these plants, N-NO
3 was significantly lower (0.93 ± 0.03 g Kg
DW and 0.82 ± 0.04 g Kg
DW, for E2 and O2, respectively) respect to the Control at T3 and the treated plants with the lowest concentrations (E1 and O1) indicating that these treatments could be useful to reduce nitrates in this edible plant. In fact, an excess consumption of nitrates is considered dangerous because nitrates could be converted into nitrite in the gut and nitrite can bind to hemoglobin preventing the blood from carrying enough oxygen or, in presence of ammine, may generate nitrosamines, known to have carcinogenic activity [
69]. For this reason, the reduction of anti-nutritional factors in horticultural products, such as nitrates in leafy vegetables, has become an important objective in agricultural research. Phosphate concentrations in plants treated with the highest concentrations of EM and OWM (E2, 1.10 ± 0.09 g Kg
DW; O2, 0.99 ± 0.22 g Kg
DW) were significantly minor to those of Control at T3 (2.13 ± 0.04 g Kg
DW), E1 (2.91 ± 0.10 g Kg
DW), and O1 (2.09 ± 0.23 g Kg
DW). Accordingly, K contents decreased significantly with the high EM and OMW concentrations (Control, 1.73 ± 0.01 g Kg
DW; E1, 2.01 ± 0.01 g Kg
DW; E2, 0.47 ± 0.03 g Kg
DW; O1, 2.19 ± 0.05 g Kg
DW; O2, 0.23 ± 0.02 g Kg
DW). A significant increase in sodium and magnesium was observed adding EM and OMW at the substrates compared with Control (Na, 0.13 ± 0.01 g Kg
DW; Mg, 1.03 ± 0.01 g Kg
DW), the application of biostimulant compounds seem to increase the availability of these mineral elements for the plant. Calcium was the only element affected simultaneously both by treatment and by the treatment concentrations. Specifically, it was higher in Control (11.60 ± 0.40 g Kg
DW). The other elements were all influenced by treatment and concentration.
The EM application has also been investigated in tomatoes [
70], and grass species [
71]. The authors showed that the addition of EM may enhance the amount of some elements, such as nitrogen, phosphorus, and potassium, promoting growth and productivity. These findings support the results of E1 and E2 treated plants which exhibited also higher growing parameter values with respect to the Control plants.
3.3. Physiological Parameters: Chlorophyll a Fluorescence
The trend of the JIP-test parameters is reported in
Figure 7. The values measured for the following parameters
; Mo; V
J;
Eo, in all the plants at T0 did not exhibit significant differences. These parameters are a useful tool to measure the electron transfer efficiency between the two photosystems of the photosynthetic apparatus, and their variations indicate how the level of this efficiency decreases, in the presence of stress. The
parameter is a measure of the maximum quantum yield of PSII for primary photochemistry, and a lowering of this value reflects a decrease in the ability to carry out photosynthesis. As shown in
Figure 7A the photosynthetic activity, expressed as
did not exhibit large variations during the period of the experiment (T0–T3). During the period of the treatment, at each time, no significant changes were detected among the Control and the treated plants, indicating that all the treated plants maintained efficient photosynthetic activity. However, it is interesting to underline that all the treated plants showed
values higher than Control plants, at any time, till T3. The highest values at T3 were detected in O1 and E1 (8.5% and 6.3%). The changes of the parameter Mo are reported in
Figure 7B. This parameter is a measure of the efficiency of the reaction centers to transfer electrons into the electron transport chain, and it increases when this efficiency is reduced. The results showed that, during the period of the treatments (T0–T3), all the treated plants exhibited a declined value of Mo. At T3 the highest changes resulted for E1, O1, and O2, whereas only a slight decline, by 2.10% and 4.41%, could be observed for E2 and Control, respectively
Figure 7B. Anyway, changes in this parameter indicated better performance of the reaction centers tendentially in the treated plants.
Figure 7C shows the changes in the VJ parameter. The VJ value reflects the reduction state of the photosynthetic apparatus, at the plastoquinone level, and it is a very useful tool to follow the level of accumulation of electrons in the electrons transport chain during the growth and especially under stress conditions which may affect the photosynthetic activity [
72]. The value of this parameter, during the examined period, from T0 to T3, resulted unchanged in E2 while it decreased in the other treated plants (O1, E1 and O2) (
Figure 7C), a significant change was observed only for O2. At T3 the differences among the VJ values were not significant among the plants. Anyway, the lowering of this parameter, indicated that, during the growth, both OMW and EM exercised a positive effect on photosynthetic electrons transport efficiency. The changes of the parameters
Eo are reported in
Figure 7D. The results indicated that it tendentially improved in all the plants at T3, except E2. The highest increase was observed in the following order: O2, O1, E1, and Control (23.87%, 16.90%, 15.22%, and 2.48% respectively). Comparison of the values at T3 showed significant differences for O1 vs. E2, O2 vs. E2, and E1 vs. E2. Also, for this parameter, the results tendentially showed a better performance in the treated plants. The implementation, although slight, of the photosynthetic efficiency in the treated plant with OMW and EM agrees with plant growth parameters, in particular the leaves number and leaf area (
Figure 1). The fluorescence parameters measurements showed that the addition of OMW to the plants was not toxic, at the assessed concentration.
The slight improvement of the photosynthetic efficiency in the treated plant with EM and OMW is in agreement with plant growth parameters, in particular the leaves number and leaf area (
Figure 1). The fluorescence indices measurement showed that the addition of OMW to the plants was not toxic, at the assessed concentrations. These results confirmed previous findings, reporting that the application of diluted filtered OMW, at neutral pH, incremented both growth and photosynthetic efficiency in tomato plants [
23] and in olive trees [
72]. The authors reported the increment of chlorophyll fluorescence parameters in olive trees (
Olea europaea L. cv. Chemlalì) when soil tillage was combined with OMW irrigation, they associated these results with the attenuation of oxidative stresses [
72]. Our findings agree with these results, as O1 and O2 treatments enhanced the photosynthetic activity, electron transport efficiency, and attenuation of reductive conditions at the photosynthetic apparatus level. Several protocols are used for the management of OMWs, in order to make them suitable for agriculture application, including mechanical steps of filtration and microbiological treatments [
23,
73]. In our study, the OMW treatment was very simple, very low time consuming and economically advantageous, as OMW was just kept settling for a week, the pH was adjusted to neutral value, and the right dilution rates were adopted. With this strategy, the toxicity due to the presence of polyphenolic compounds, conferring dark color and very low pH, was strongly reduced. Moreover, this procedure could not alter the organic and chemical composition of the OMW, which may have supplied nitrogen, phosphorus, and potassium [
74] with a positive effect of photosynthetic efficiency. The addition of EM may enhance the amount of absorbable nutrients, such as nitrogen, phosphorus, and potassium [
71], and maintain a high efficiency of PSII; these are two important processes for plant growth and productivity [
54]. These findings support the observed trend in E1 and E2 treated plants which exhibited a facilitated photosynthetic performance.
3.4. Total Polyphenolic Content and Individual Phenolic Determination
The values of the total phenolic content, measured on all the plants at T0, did not exhibit significant differences. In general, the extracts, of both the Control and treated plants, exhibited some changes in the total polyphenolic content during the period of the experiment (T0–T3) (
Table 2).
The highest total polyphenolic content significant increase was observed at T1 in the Control and O1 plants, resulting in 29% higher than at the initial time, under both conditions (
Table 2). For the other treatments, changes were not relevant, whereas the highest increment was observed after T2, by 7.5%, 18.0% and 19.1%, for O2, O1 and E1 extracts, respectively, while E2 did not induce any polyphenolic increase, with respect to the initial time. At the end of the growth period T3, in the Control and all the treated plants, the differences among the polyphenolic content in the extracts were not significant and the values were in the range of 18.6–20.9 mg g
. The individual phenolic composition was quantified, and it is reported in detail in
Table 3. Also in this case, the values measured on all the plants at T0 did not exhibit significant differences, for all the phenolic acids. At the initial time caffeic acid, isochlorogenic acid, rutin, and a very small amount of chlorogenic acid were found. As shown in
Table 3, these compounds resulted increased tendentially in all the treated plants, at the harvesting time. At T3 the extracts of the treated plants showed a significant higher amount of the phenolic compounds detected, with respect to the Control. For the chlorogenic acid, at a very low concentration at the initial time, the highest content followed the order: O2, E1, E2, O1, in the range of 2.16–2.45-fold higher than Control (
Table 3). The same trend could be observed for the caffeic acid, although the differences were less pronounced, in the range of 1.91-fold and 2.11-fold higher than Control. Whereas the amount of caffeic acid was almost the same in all the treatments, by contrast, isochlorogenic acid content showed significant differences among all the plants, except for E1 and Control, ranging from 1.02 to 2.44-fold the amount of Control. Rutin content in the extracts of the treated plants ranged in a value 1.28–1.50-fold the one of the Control extract (
Table 3). The phenolic composition resulted interestingly different among the plants. The higher amount of chlorogenic acid, caffeic acid, isochlorogenic acid, and rutin with respect to the Control plants evidenced how the improvement of the photosynthesis in these plants promoted a positive effect on the production of these antioxidant compounds.The value of total polyphenolic content may result non proportional to or higher than the value obtained by adding all the phenolic compounds quantified by HPLC analyses, and this is our case. The explanation is that the method to determine the total phenolic content may imply the reaction of a hydroxyl group of all phenolic compounds and their degradation product, which cannot be detected by HPLC analysis [
75].
It is generally reported that the increment of polyphenolic substances, such as other antioxidants, is induced by the occurrence of oxidative stresses, to counteract the effect of the presence of free radicals [
76,
77]. However, many officinal plants, such as
Tanacetum balsamita L., are constitutively able to produce a high amount of polyphenols also in absence of oxidative stress [
13,
19]. In the present study, stress in plants did not occur, neither in Control nor in treated plants, as revealed by fluorescence measurements. The most relevant result concerned the presence of important antioxidant phenolic compounds, in all the treated plants, at a concentration higher than in the Control ones and these results underline the importance of the utilization of both OMW and EM. The phenolic acids found in the extracts are well-known compounds synthesized in costmary, which can confer to the leaves anti-inflammatory, carminative and detoxifying properties [
19,
78].


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







