
Practices of Organic Medium Enclosed Trough and Amaranth Species Variation Enhanced Growth, Nutritional Composition and Bioactive Compounds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site, and Treatments of the OMET System
2.2. Establishment of Seedling on the OMET System
2.3. Growth and Yield Data Collection
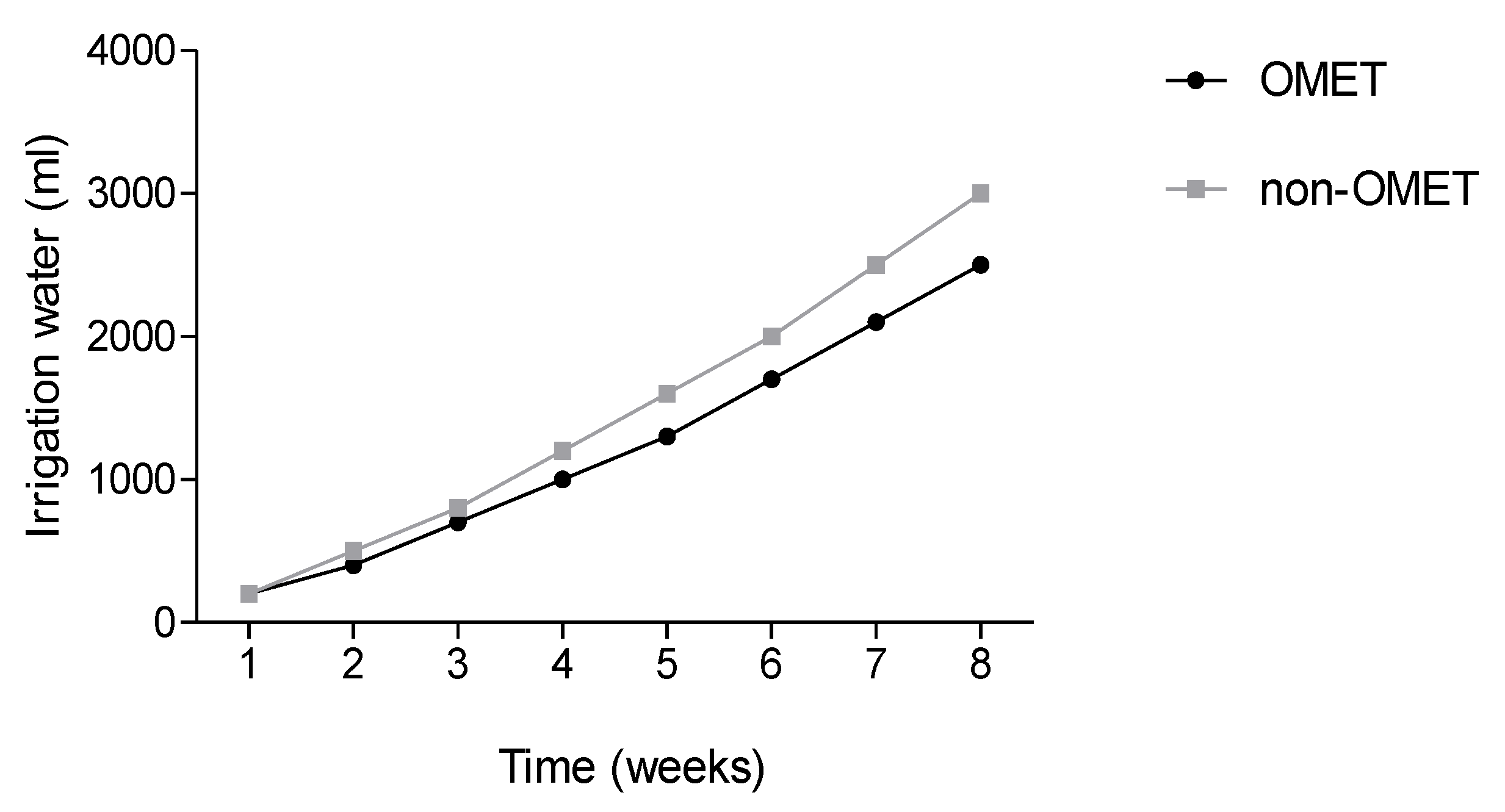
2.4. Determination of Total Irrigation Water
2.5. Determination of Bioactive Compounds and HPLC-MS-QTOF Untargeted Metabolites
2.6. Determination of Protein
2.7. Determination of Minerals or Trace Elements
2.8. Determination of Amino Acids
2.9. Data Analysis
3. Results
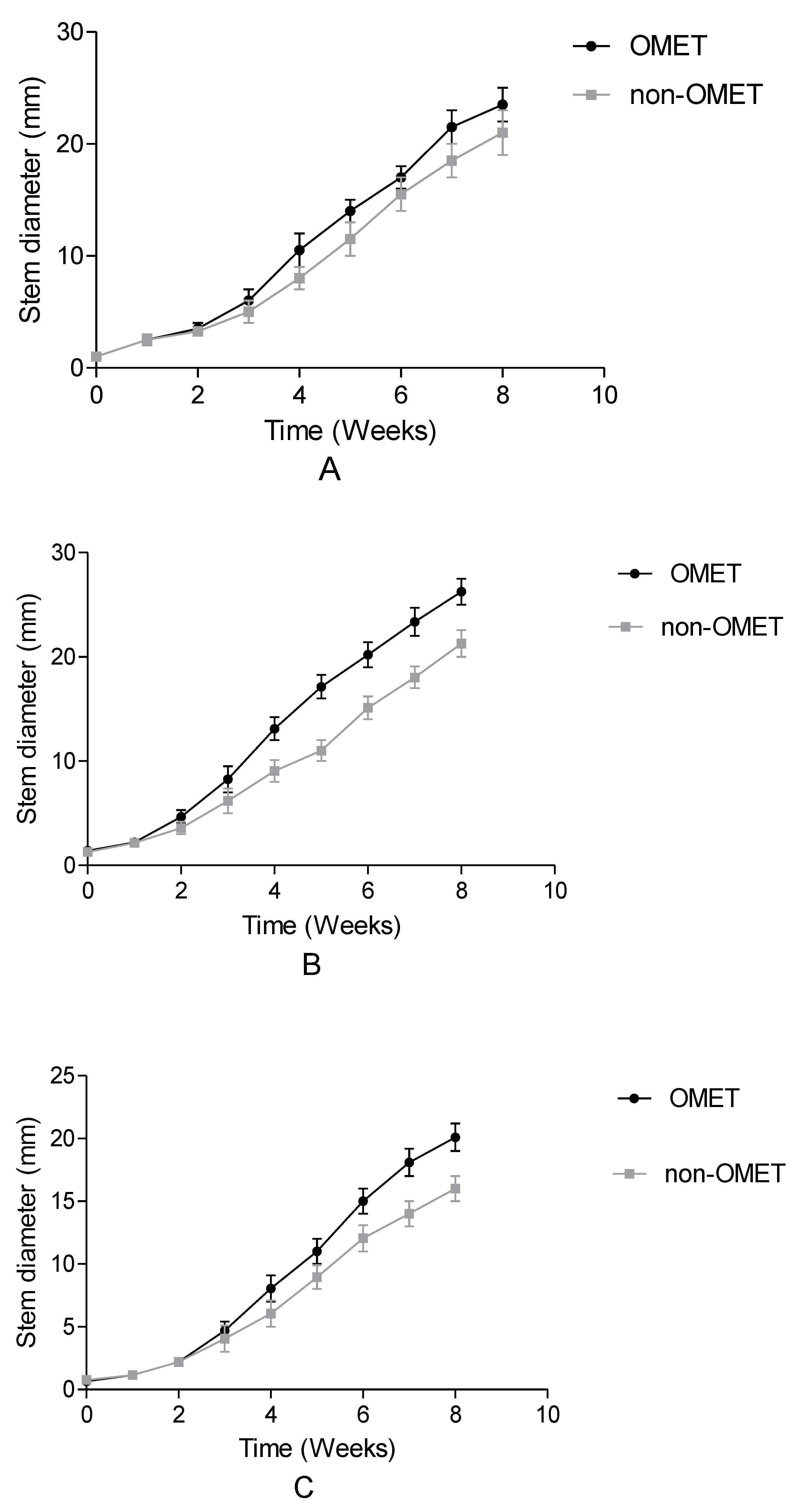
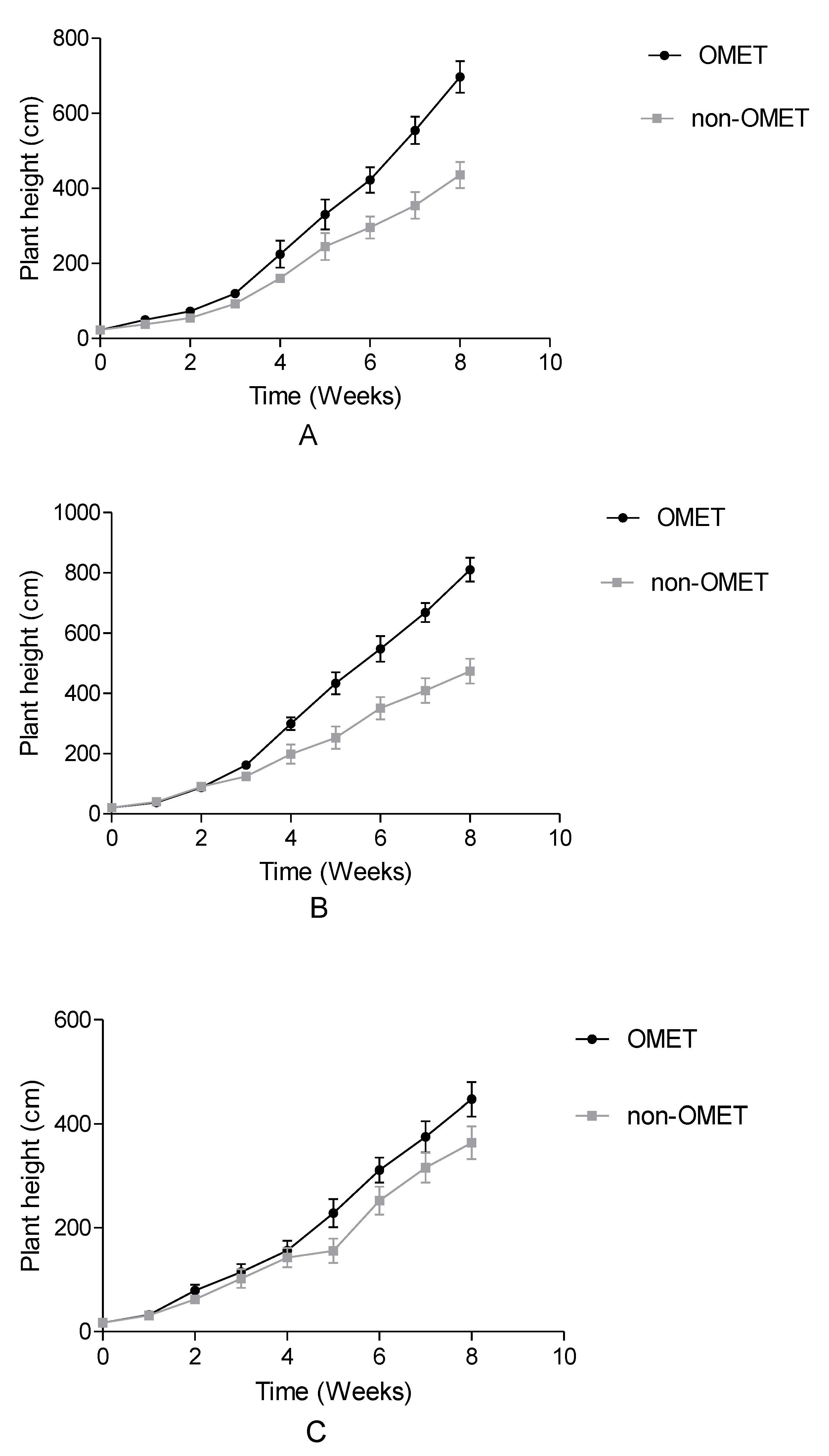
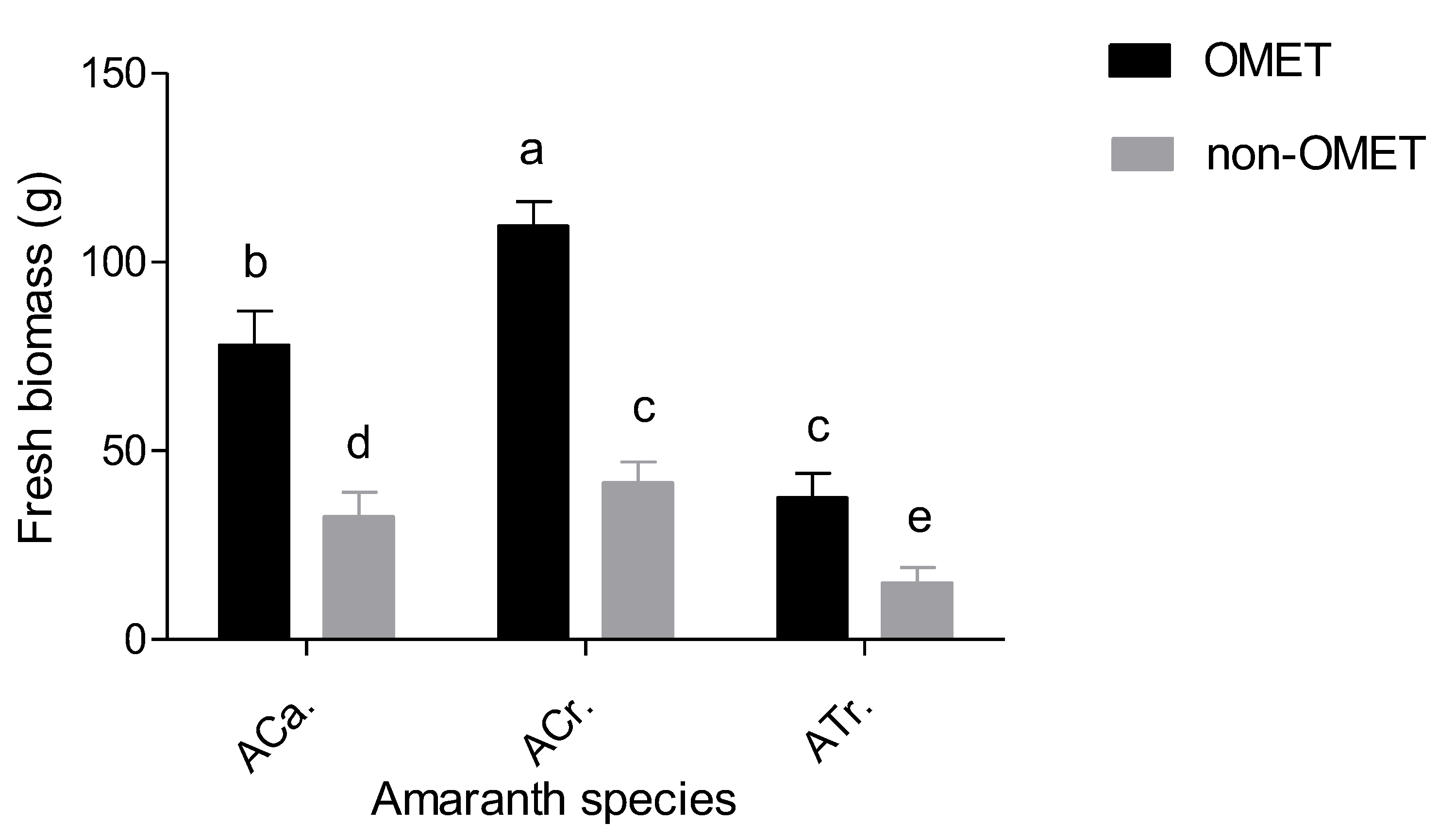
3.1. Effect between Growth Conditions and Species on Growth, Flowering, Irrigation Water and Yield Attributes of Three Amaranth Species
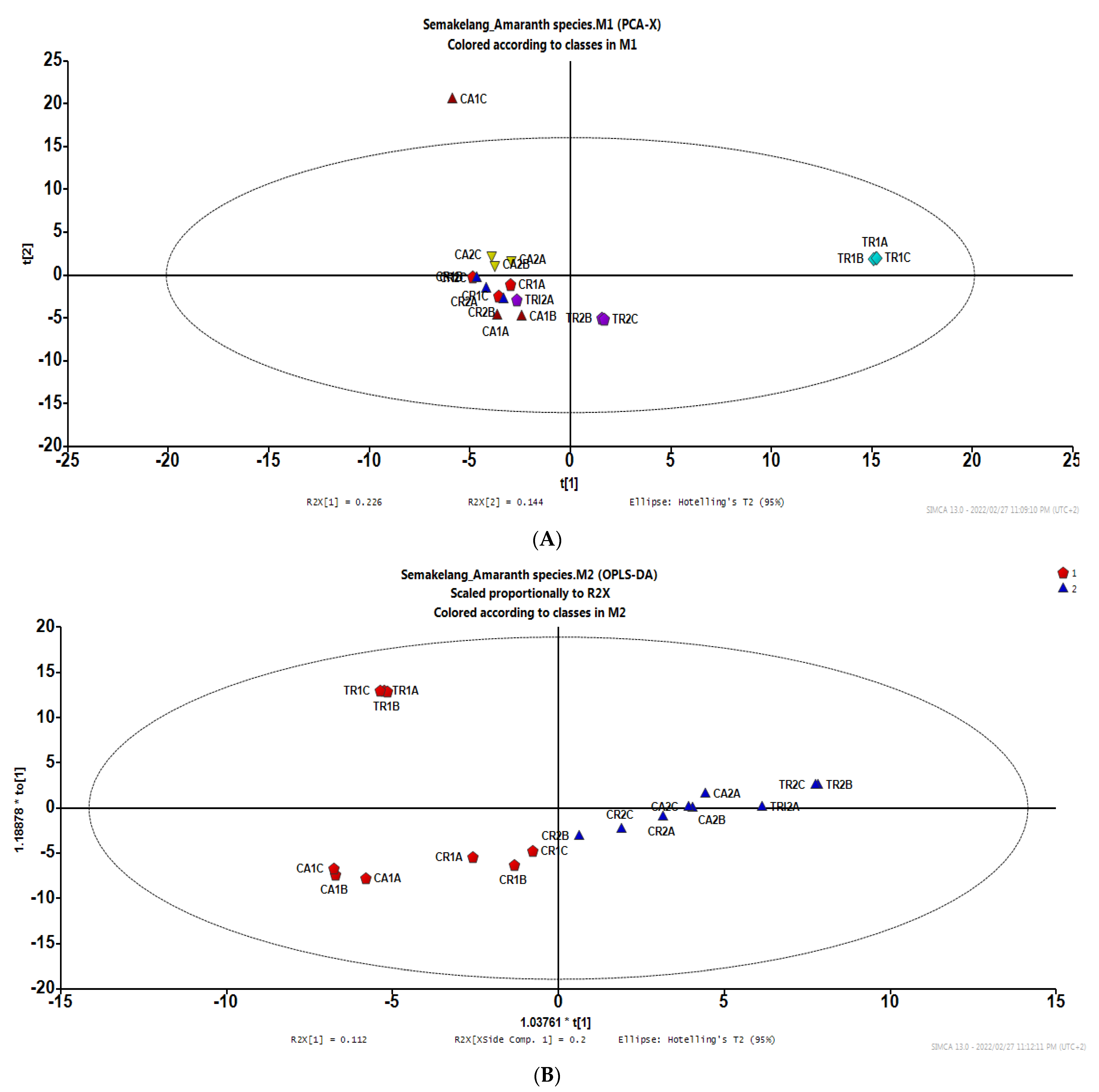
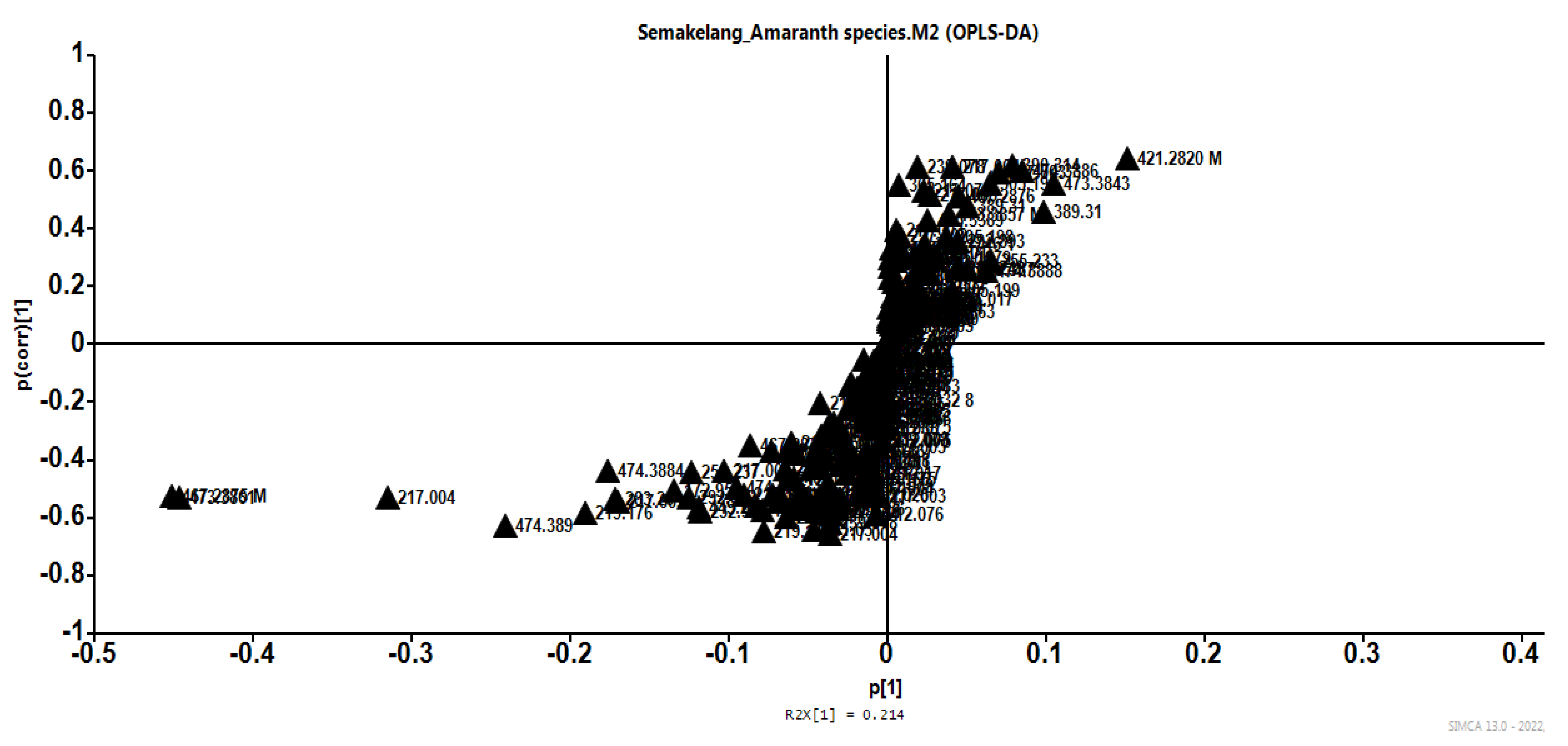
3.2. Interactive Effect between Growth Condition and Species on Bioactive Compounds and HPLC-MS-QTOF Metabolites
3.2.1. Bioactive Compounds
3.2.2. HPLC-MS-TOF Metabolites
3.3. Interactive Effect between Growth Condition and Species on Nutritional Composition
3.3.1. Protein and Nutritive Minerals
3.3.2. Amino Acid Composition
4. Discussion
5. Conclusions and Recommendations
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bvenura, C.; Afolayan, A.J. The role of wild vegetables in household food security in South Africa: A review. Food Res. Int. 2015, 76, 1001–1011. [Google Scholar] [CrossRef]
- South-East Asia Region; WHO. Regional Nutrition Strategy: Addressing Malnutrition and Micronutrient Deficiencies; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Taia, W.K.; Shehata, A.A.; Manaser, M.I.; El-Shamy, I.M. Vegetative morphological variations within some Egyptian Amaranthus L. species. Jordan J. Biol. Sci. 2021, 14, 137–146. [Google Scholar]
- Sarker, U.; Oba, S. Salinity stress enhances colour parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Maces, L.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Villagómez-González, B.B. Edible leafy plants from Mexico as sources of antioxidant compounds, and their nutritional, nutraceutical and antimicrobial potential: A review. Antioxidants 2020, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Suitability of Amaranthus species for alleviating human dietary deficiencies. S. Afr. J. Bot. 2018, 115, 65–73. [Google Scholar] [CrossRef]
- Ngoroyemoto, N.; Gupta, S.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Effect of organic biostimulants on the growth and biochemical composition of Amaranthus hybridus L. S. Afr. J. Bot. 2019, 124, 87–93. [Google Scholar] [CrossRef]
- Sarmadi, B.; Rouzbehan, Y.; Rezaei, J. Influences of growth stage and nitrogen fertilizer on chemical composition, phenolics, in situ degradability and in vitro ruminal variables in Amaranth forage. Anim. Feed. Sci. Technol. 2016, 215, 73–84. [Google Scholar] [CrossRef]
- Ferreira, J. OMET Farming Better than Hydroponics? Farmer’s Weekly. 2013. Available online: https://www.farmersweekly.co.za (accessed on 10 March 2021).
- Mokgalabone, T.T.; Mpai, S.; Ndhlala, A.R. Organic Medium Enclosed Trough Growing Technique Improves Abelmoschus esculentus (Okra) Growth, Yield and Some Nutritional Components. Appl. Sci. 2023, 13, 5645. [Google Scholar] [CrossRef]
- Mpai, S.; Mokganya, L.M.; Raphoko, L.; Masoko, P.; Ndhlala, A.R. Untargeted metabolites and chemometric approach to elucidate the response of growth and yield attributes on different concentrations of an amino acid based biostimulant in two lettuce cultivars. Sci. Hortic. 2022, 306, 111478. [Google Scholar] [CrossRef]
- Mpai, S.; Du Preez, R.; Sultanbawa, Y.; Sivakumar, D. Phytochemicals and nutritional composition in accessions of Kei-apple (Dovyalis caffra): Southern African indigenous fruit. Food Chem. 2018, 253, 37–45. [Google Scholar] [CrossRef]
- Mpai, S.; Sivakumar, D. Influence of growing seasons on metabolic composition, and fruit quality of avocado cultivars at ‘ready-to-eat stage’. Sci. Hortic. 2020, 265, 109159. [Google Scholar] [CrossRef]
- Makkar, H.P.; Siddhuraju, P.; Becker, K. Plant Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007; Volume 393, pp. 1–122. [Google Scholar]
- Tambe, V.D.; Bhambar, R.S. Estimation of total phenol, tannin, alkaloid, and flavonoid in Hibiscus tiliaceus Linn. Wood extracts. J. Pharmacogn. Phytochem. 2014, 2, 2321–6182. [Google Scholar]
- Managa, M.G.; Sultanbawa, Y.; Sivakumar, D. Effects of different drying methods on untargeted phenolic metabolites, and antioxidant activity in Chinese cabbage (Brassica rapa L. subsp. chinensis) and nightshade (Solanum retroflexum Dun.). Molecules 2020, 25, 1326. [Google Scholar] [CrossRef] [PubMed]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Rockville, MD, USA, 1990. [Google Scholar]
- Mathipa, M.M.; Mphosi, M.S.; Masoko, P. Phytochemical Profile, Antioxidant Potential, Proximate and Trace Elements Composition of Leaves, Stems and Ashes from 12 Combretum spp. Used as Food Additives. Int. J. Plant Biol. 2022, 13, 561–578. [Google Scholar] [CrossRef]
- Yahaya, Y.; Birnin-Yauri, U.A.; Bagudo, B.U.; Noma, S.S. Quantification of macro and micro elements in selected green vegetables and their soils from Aliero agricultural fields in Aliero, Kebbi State, Nigeria. J. Soil Sci. Environ. Manag. 2012, 3, 207–215. [Google Scholar]
- Manyelo, T.G.; Sebola, N.A.; Mabelebele, M. Nutritional and phenolic profile of early and late harvested Amaranth leaves (Amaranthus cruentus) grown under cultivated conditions. Agriculture 2020, 432, 5. [Google Scholar]
- Saeed, R.; Ahmad, R. Vegetative growth and yield of tomato as affected by the application of organic mulch and gypsum under saline rhizosphere. Pak. J. Bot. 2009, 41, 3093–3105. [Google Scholar]
- Ferrini, F.; Fini, A.; Frangi, P.; Amoroso, G. Mulching of ornamental trees: Effects on growth and physiology. Arboric Urban Forum 2008, 34, 157. [Google Scholar] [CrossRef]
- Ngala, J.M.; Ndiso, J.B.; Mundi, E.M. Effects of selected organic mulches on growth and yield of Amaranth in Kilifi country. Int. J. Agric. Environ. Bioresearch 2019, 4, 6. [Google Scholar]
- Department of Agriculture, Forestry and Fisheries (DAFF). Amaranthus Production Guideline; Yumpu: Pretoria, South Africa, 2010.
- Scharenbroch, B.C.; Lloyd, J.E. Particulate organic matter and soil N availability in urban landscapes. Arboric Urban Forum 2006, 32, 180. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Hallmann, E.; Rembiałkowska, E. Effects of organic and conventional production systems on the content of bioactive substances in four species of medicinal plants. Biol. Agric. Hortic. 2015, 31, 118–127. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S., III. Resource availability and plant anti-herbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Ferreira, V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2017, 33, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Skotnica, J.; Matoušková, M.; Nauš, J.; Lazár, D.; Dvořák, L. Thermoluminescence and fluorescence study of changes in Photosystem II phytochemistry in desiccating barley leaves. Photosynth. Res. 2000, 65, 29–40. [Google Scholar] [CrossRef]
- Kwenin, W.K.J.; Wolli, M.; Dzomeku, B.M. Assessing the nutritional value of some African indigenous green leafy vegetables in Ghana. J. Anim. Plant Sci. 2011, 10, 1300–1305. [Google Scholar]
- Hanif, R.; Iqbal, Z.; Iqbal, M.; Hanif, S.; Rasheed, M. Use of vegetables as nutritional food: Role in human health. J. Agric. Biol. Sci. 2006, 1, 18–22. [Google Scholar]
- Wijewardana, C.; Reddy, K.R.; Bellaloui, N. Soybean seed physiology, quality, and chemical composition under soil moisture stress. Food Chem. 2019, 278, 92–100. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Fisher, D.K.; Abel, C.A. Soybean seed composition constituents as affected by drought and Phomopsis in phomopsis susceptible and resistant genotypes. J. Crop Improv. 2012, 26, 428–453. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B.S.M.A.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Amaranth Species | Time of Flowering (Weeks) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| OMET system | Amaranthus caudatus | X | |||||||
| Amaranthus cruentus | X | ||||||||
| Amaranthus tricolor | X | ||||||||
| non-OMET system | Amaranthus caudatus | X | |||||||
| Amaranthus cruentus | X | ||||||||
| Amaranthus tricolor | X | ||||||||
| Amaranth Species & Treatment | Total Phenolic (mg GAE/100 g DW) | Total Flavonoids (mg CA/100 g DW) | Total Tannins (mg GAE/100 g DW) | |
|---|---|---|---|---|
| A. caudatus | OMET | 210.24 ± 0.033 d | 391.26 ± 0.046 d | 4.09 ± 0.042 d |
| non-OMET | 259.68 ± 0.046 b | 453.34 ± 0.036 b | 9.60 ± 0.022 a | |
| A. cruentus | OMET | 209.65 ± 0.035 d | 382.60 ± 0.047 e | 3.98 ± 0.043 d |
| non-OMET | 262.45 ± 0.028 a | 473.28 ± 0.021 a | 8.78 ± 0.031 b | |
| A. tricolor | OMET | 207.70 ± 0.0.036 e | 364.24 ± 0.038 f | 3.61 ± 0.038 d |
| non-OMET | 257.24 ± 0.040 c | 447.59 ± 0.042 c | 6.89 ± 0.040 c | |
| Chl a (mg/kg) | Chl b (mg/kg) | Chl a + b (mg/kg) | ||
|---|---|---|---|---|
| A. caudatus | OMET | 0.306 ± 0.005 a | 47.779 ± 0.004 a | 48.085 ± 0.005 a |
| non-OMET | 0.301 ± 0.0047 b | 47.142 ± 0.003 a | 47.443 ± 0.007 a | |
| A. cruentus | OMET | 0.301 ± 0.0048 b | 47.142 ± 0.008 a | 47.443 ± 0.008 a |
| non-OMET | 0.296 ± 0.0034 c | 46.306 ± 0.003 b | 46.603 ± 0.004 a | |
| A. tricolor | OMET | 0.289 ± 0.044 d | 45.253 ± 0.048 c | 45.543 ± 0.045 a |
| non-OMET | 0.197 ± 0.036 e | 30.749 ± 0.041 d | 30.946 ± 0.039 b | |
| Retention Time (min) | Exact Mass (g/mol) | Mass Generated ESI (-) TOF MS (g/mol) | Fragmentation | Chemical Formula | Tentative Structural Assignment |
|---|---|---|---|---|---|
| 15.658 | 474.4557 | 474.08423 | 474.08758:246 475 | C21H30O12 | 6-Feruloylglucose 2,3,4-trihydroxy-3-methylbutylglycoside |
| 8093 | 219.23 | 218.10211 | 218.10655:4843 219 | C9H17NO5 | Pantothenic acid |
| 21.15 | 468.46 | 467.15897 | 467.13629:1292 468 | C22H28O11 | 6-O-(4-Hydroxybenzoyl)-ajugol |
| 17.63 | 420.41 | 420.3132 | 268.03810:269.04341 | C21H24O9 | Apigenin 7-O-glucoside |
| Amaranth Species & Treatment | Protein (%) | |
|---|---|---|
| A. caudatus | OMET | 24.1 ± 0.039 b |
| non-OMET | 21.4 ± 0.044 d | |
| A. cruentus | OMET | 28.6 ± 0.46 a |
| non-OMET | 24.1 ± 0.040 b | |
| A. tricolor | OMET | 22.7 ± 0.036 c |
| non-OMET | 20.3 ± 0.042 d | |
| p-value | 0.04 | |
| 3A | ||||||
|---|---|---|---|---|---|---|
| Amaranth Species & Treatment | Macro Elements (mg/kg DW) | |||||
| Ca | Mg | K | p | |||
| A. caudatus | OMET | 104 ± 0.043 b | 77.3 ± 0.032 b | 251 ± 0.041 c | 29.7 ± 0.045 b | |
| non-OMET | 104 ± 0.043 b | 62.6 ± 0.030 d | 244 ± 0.038 d | 21.8 ± 0.037 c | ||
| A. cruentus | OMET | 130 ± 0.038 a | 63.3 ± 0.029 d | 254 ± 0.046 b | 34.7 ± 0.043 a | |
| non-OMET | 88.3 ± 0.033 d | 56.6 ± 0.025 e | 217 ± 0.033 f | 22.2 ± 0.039 c | ||
| A. tricolor | OMET | 104 ± 0.039 b | 82.5 ± 0.038 a | 276 ± 0.05 a | 30.3 ± 0.041 b | |
| non-OMET | 90.5 ± 0.041 c | 66.2 ± 0.032 c | 226 ± 0.035 e | 21.5 ± 0.036 c | ||
| 3B | ||||||
| Microelements (mg/kg DW) | ||||||
| Cu | Mn | Fe | Se | Zn | ||
| A. caudatus | OMET | 0.89 ± 0.023 c | 1.73 ± 0.046 b | 2.62 ± 0.05 b | 8.03 ± 0.049 a | 1.11 ± 0.024 c |
| non-OMET | 0.84 ± 0.021 d | 1.46 ± 0.043 d | 2.5 ± 0.050 c | 7.01 ± 0.048 d | 1.09 ± 0.036 c | |
| A. cruentus | OMET | 1.04 ± 0.034 a | 1.78 ± 0.048 b | 2.52 ± 0.04 c | 8.13 ± 0.050 a | 1.66 ± 0.048 a |
| non-OMET | 0.89 ± 0.023 c | 1.17 ± 0.041 e | 2.51 ± 0.04 c | 7.69 ± 0.05 b | 1.28 ± 0.045 b | |
| A. tricolor | OMET | 0.93 ± 0.025 b | 2.16 ± 0.05 a | 3.41 ± 0.39 a | 7.28 ± 0.05 c | 1.27 ± 0.047 b |
| non-OMET | 0.78 ± 0.022 e | 1.64 ± 0.04 c | 2.47 ± 0.05 c | 6.36 ± 0.048 e | 0.95 ± 0.039 d | |
| Essential Amino Acids | |||||||
|---|---|---|---|---|---|---|---|
| Threonine | Valine | Isoleucine | Lysine | Leucine | Phenylalanine | ||
| A. caudatus | OMET | 0.92 ± 0.024 b | 1.09 ± 0.043 c | 0.97 ± 0.031 b | 2.25 ± 0.05 a | 1.66 ± 0.044 b | 2.36 ± 0.05 a |
| non-OMET | 0.66 ± 0.025 d | 0.84 ± 0.027 d | 0.76 ± 0.026 c | 1.3 ± 0.034 b | 1.25 ± 0.038 c | 1.14 ± 0.035 c | |
| A. cruentus | OMET | 1.15 ± 0.034 a | 1.45 ± 0.041 a | 1.29 ± 0.027 a | 2.37 ± 0.05 a | 2.21 ± 0.042 a | 2.54 ± 0.051 a |
| non-OMET | 0.93 ± 0.036 b | 1.22 ± 0.042 b | 1.12 ± 0.039 a | 2.19 ± 0.050 a | 1.92 ± 0.051 b | 1.95 ± 0.043 b | |
| A. tricolor | OMET | 0.88 ± 0.021 c | 1.08 ± 0.034 c | 0.98 ± 0.028 b | 1.62 ± 0.036 b | 1.66 ± 0.041 b | 1.47 ± 0.038 c |
| non-OMET | 0.43 ± 0.021 e | 0.52 ± 0.026 d | 0.46 ± 0.022 d | 0.8 ± 0.012 c | 0.74 ± 0.024 d | 0.71 ± 0.023 d | |
| Non-essential amino acids | |||||||
| Arginine | Serine | Glycine | Aspartate | Glutamate | |||
| A. caudatus | OMET | 1.12 ± 0.047 b | 0.97 ± 0.034 b | 1.37 ± 0.05 b | 1.2 ± 0.041 c | 1.52 ± 0.05 c | |
| non-OMET | 0.86 ± 0.031 c | 0.7 ± 0.030 c | 0.95 ± 0.041 c | 0.96 ± 0.042 c | 1.29 ± 0.049 c | ||
| A. cruentus | OMET | 1.37 ± 0.048 a | 0.98 ± 0.036 b | 1.33 ± 0.044 b | 1.33 ± 0.044 b | 1.75 ± 0.05 b | |
| non-OMET | 1.54 ± 0.05 a | 1.16 ± 0.045 a | 1.46 ± 0.048 a | 2.02 ± 0.05 a | 2.55 ± 0.05 a | ||
| A. tricolor | OMET | 1.12 ± 0.049 b | 0.91 ± 0.042 b | 1.21 ± 0.047 b | 1.53 ± 0.049 b | 1.95 ± 0.05 b | |
| non-OMET | 0.53 ± 0.028 d | 0.46 ± 0.022 c | 0.59 ± 0.029 d | 0.6 ± 0.030 d | 0.73 ± 0.032 d | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mopai, M.G.; Mpai, S.; Ndhlala, A.R. Practices of Organic Medium Enclosed Trough and Amaranth Species Variation Enhanced Growth, Nutritional Composition and Bioactive Compounds. Appl. Sci. 2023, 13, 12574. https://doi.org/10.3390/app132312574
Mopai MG, Mpai S, Ndhlala AR. Practices of Organic Medium Enclosed Trough and Amaranth Species Variation Enhanced Growth, Nutritional Composition and Bioactive Compounds. Applied Sciences. 2023; 13(23):12574. https://doi.org/10.3390/app132312574
Chicago/Turabian StyleMopai, Maeleletse Glas, Semakaleng Mpai, and Ashwell R. Ndhlala. 2023. "Practices of Organic Medium Enclosed Trough and Amaranth Species Variation Enhanced Growth, Nutritional Composition and Bioactive Compounds" Applied Sciences 13, no. 23: 12574. https://doi.org/10.3390/app132312574
APA StyleMopai, M. G., Mpai, S., & Ndhlala, A. R. (2023). Practices of Organic Medium Enclosed Trough and Amaranth Species Variation Enhanced Growth, Nutritional Composition and Bioactive Compounds. Applied Sciences, 13(23), 12574. https://doi.org/10.3390/app132312574