Featured Application
Screening for new yeast species may to ease the production of no/low-alcohol beers or may enable the obtaining of beers with unique sensory characteristics.
Abstract
Among many methods to produce low/no-alcohol beers, using special yeasts has gained a substantial interest in the brewing industry. This approach relies on the fact that many non-Saccharomyces yeasts do not utilize maltose, which is the main sugar found in brewer’s wort. Additionally, these yeasts may allow the production of a beer with unique sensory characteristics. The aim of the study was to evaluate the potential of 18 non-Saccharomyces yeast strains in the production of low-alcohol beer. As a control strain, S. cerevisiae US-05 was used. The study consisted of two parts: microbiological evaluation and small-scale fermentations. In the microbiological part, ability to ferment sugars found in a wort, resistance to stress factors, phenolic off-flavor production, and enzymatic activities of β-glucosidase and β-lyase were evaluated. In the second part of the study, yeasts were used to produce a beer from 9.3 °Plato wort. During the fermentation, its dynamics was analyzed. The obtained beers were analyzed regarding their alcohol content, pH, acidity, and color. All of the evaluated strains produced low levels of alcohol. Two of the evaluated strains were characterized by especially high β-glucosidase activity. Based on the obtained results, six of the evaluated strains are promising in brewing.
1. Introduction
In recent years, there has been a multidirectional growth of a brewing industry. One such direction is the so-called craft beer revolution. It is safe to say that nowadays, brewers and consumers are reaping the benefits of this movement. Prior to 2009, craft beer sales accounted for less than 2% globally, which nearly doubled to 3.9% in 2021 [1,2]. Thanks to that, consumers are continually looking for new, unique products. According to Anderson [2], the relatively new market of reduced/no alcohol beers can be seen as an extension of the craft beer revolution. Although these beers have a relatively low market share, it is constantly growing [3]. According to Kokole et al. [4], non-alcoholic beer (containing up to 0.5% ethanol) accounted for 3.8% of European beer volume in 2019. Anderson [2] states that no-lo beers already have 3% of the global market, with further growths expected. With such a potential, the brewing industry is looking for new ways to produce these beers. Producing a beer with a normal alcohol content and then removing that alcohol to obtain desired levels is not energetically viable and problematic in many regards. An interesting alternative to these methods is to produce a beer with reduced alcohol levels in the first place using biological methods. Among the methods which enable this, using non-Saccharomyces yeast has gained a great interest [5].
For a long time, Saccharomyces cerevisiae was the main yeast species used in most of fermentative industries. This is related to their unique biological characteristics of high fermentation capacity, good alcohol production, and elevated resistance to adverse environmental conditions [6]. The other yeast species found in fermentation environments, non-Saccharomyces yeast, were thought to be a source of microbiological spoilage rather than a desired organism. However, in recent decades this view has begun to shift. For example, they are now seen as a factor which can add a complexity to the flavor profile of wines [7]. The brewing science has also expressed interest in these yeasts, as their application can result in beers with unique sensory characteristics. This is related to the high bioflavoring potential of some of the strains. Individual strains can be characterized by desirable profiles of volatile compounds [8,9,10]. As an extension of bioflavoring potential, some strains possess higher enzymatic activities of β-glucosidase and β-lyase than S. cerevisiae [11]. These enzymatic activities can result in a beer with more pronounced fruity, citrous, or tropical aromas, due to the release of flavor compounds from their precursors [12].
However, the main problem with non-Saccharomyces yeasts is the fact that the majority of them are poor fermenters, which makes them problematic to use in standard beer fermentations. This has been evident in wine studies and is even more problematic in the case of the brewing industry. The fermentative sugars found in brewers wort are glucose (10–15%), fructose (1–2%), sucrose (1–2%), maltose (50–60%), and maltotriose (15–20%) [13]. S. cerevisiae yeast are characterized by a high fermentative capacity, as they readily utilize sugars found in wort (except for the maltotriose, which utilization is strain-dependent). As maltose is the major sugar found in the brewers’ wort, using yeasts which are unable to ferment this sugar would result in a beer with a reduced alcohol content. The ability to utilize maltose is rather scarce among non-Saccharomyces yeasts. However, this potential drawback can also be seen as their biggest advantage regarding the production of reduced-alcohol beer. This approach would be easy to implement in most breweries as the only factor that would change about the production process is the applied yeast species [3,14,15,16]. Yeasts used in the production of low-alcohol beers should not be able to ferment maltose, be able to grow in the presence of iso-AA’s, not produce phenolic off-flavors (POF-), flocculate easily, and, most importantly, be consumer-safe [17].
The aim of the study was to evaluate the suitability of selected yeast strains in the brewing industry. The first part of the study was the microbiological characterization of the strains, conducted by determining their ability to ferment sugars found in the wort, resistance to stress factors, and enzymatic activities: phenolic off flavor production, β-glucosidase, and β-lyase activities. In the second part of the study, the yeasts were used in small-scale beer fermentations. The course of the fermentation process and basic physicochemical characteristics of resulting beers were determined.
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals
The following chemicals were used in the study: bromothymol blue (analytical grade, EuroChem Group, Zug, Switzerland), carbonate buffer (Arcos Organics, Geel, Belgium), citric acid (analytical grade, STANLAB, Lublin, Poland), dextrose (analytical grade, STANLAB, Poland), disodium phosphate (analytical grade, EuroChem Group, Switzerland), ethanol (96%, STANLAB, Poland), ferulic acid (analytical grade, Ambeed, Arlington Heights, IL, USA), glucose (analytical grade, STANLAB, Poland), Isomerized Hop Extract 30% (30% of iso-α-acids, supplied by Hopsteiner, New York, NY, USA), lactose (analytical grade, EuroChem Group, Switzerland), maltose (analytical grade, STANLAB, Poland), p-coumaric acid (purity ≥ 98.0%, Sigma-Aldrich, Overijse, Belgium), pyridoxal-5′-phosphate (purity ≥ 98.0%, Sigma-Aldrich, Madrid, Spain), S-Methyl-L-cysteine (purity ≥ 98.0%, Sigma-Aldrich, Spain), sodium chloride (analytical grade, EuroChem Group, Switzerland), sodium hydroxide (Tarchem, Tarnowskie Góry, Poland), sucrose (analytical grade, Chempur, Piekary Śląskie, Poland), and 4-Nitrophenyl β-D-glucopyranoside—pNPG (purity ≥ 98.0%, Pol-Aura, Dywity, Poland).
The following microbiological substrates were used in the study: bacteriological agar (Biomaxima, Lublin, Poland), peptone (Thermo Scientific, Waltham, MA, USA), Sabouraud agar (Biocorp, Warsaw, Poland), Sabouraud Broth (Biocorp, Poland), Yeast Carbon Base (HiMedia Laboratories, Thane, India), and yeast extract (Pol-Aura, Poland).
2.1.2. Microorganisms
A total of 18 strains of non-Saccharomyces yeast and 1 control strain of Saccharomyces cerevisiae were used in the study. Non-Saccharomyces strains used in the study were obtained from the pure culture collection of the Department of Fermentation Technology and Microbiology of the University of Agriculture in Kraków. The cultures were previously isolated from cold climate grapes by Cioch-Skoneczny et al. [18,19,20]. The non-Saccharomyces cultures were Hanseniaspora uvarum MG971252 (HU1), H. uvarum MG971255 (HU2), Kluyveromyces lactis MG971263 (KL1), Metschnikowia pulcherrima MG971245 (MP1), M. pulcherrima MG971247 (MP2), M. pulcherrima MG971249 (MP3), M. pulcherrima MG971250 (MP4), M. pulcherrima MG971256 (MP5), M. pulcherrima MG971257 (MP6), M. pulcherrima MG971260 (MP7), M. pulcherrima MG971262 (MP8), M. pulcherrima MG971264 (MP9), Nakazawaea ishiwadae MG971259 (NI1), Torulaspora delbrueckii MG971248 (TD1), Wickerhamomyces anomalus MG971261 (WA1), W. onychis MG971246 (WO1), Zygoascus meyerae MG970696 (ZM1), and Z. meyerae MG971244 (ZM2).
Additionally, as a control culture, Fermentis SafAle™ US-05 Saccharomyces cerevisiae strain (S.C.) was used.
2.2. Microbial Analyses
2.2.1. Sugar Utilization
The analysis was conducted according to Jimoh et al. [18] and Kurtzman et al. [19]. The ability of the yeast to ferment specific sugars was determined using the following sugars: glucose, sucrose, maltose, and lactose. The basal medium contained 0.5% (w/v) yeast extract, 0.75% (w/v) peptone, 2% of sugar, and bromothymol blue indicator. The inoculated growth medium tubes were incubated at a temperature of 28 °C for 10 days. Thus, gas production and medium color change indicated the fermentative activity of the yeast strains.
Interpretation of results:
- -: no gas bubble,
- +/-: inconclusive (gas bubble in some repetitions),
- +: gas bubble filling less than half of the Durham test tube in more than a half of the cases,
- ++: gas bubble filling more than half the Durham test tube in more than a half of the cases.
2.2.2. Resistance to Stress Factors
The analysis was conducted according to Hong et al. [20]. The medium was based on 2% YPD medium (pH 5.4), with different gradients of ethanol (addition of 96% ethanol) and iso-α-acids (addition of ethanolic solution of Iso-Extract 30%). The composition of YPD medium was 1% yeast extract, 2% (w/v) peptone and 2% (w/v) dextrose. The ethanol grades were 1.5, 3.0, 4.5, 6.0, 7.5, and 9.0% by volume, while the iso-α-acids grades were 50 and 90 IBU (ppm iso-α-acids). A strain was considered to have specific tolerance when the value of OD600 ≥ 0.1.
2.2.3. Phenolic Off-Flavor Production
The analysis was conducted according to a modified method of Mertens et al. [21]. The modification was related to a higher volume of medium used. Strains were inoculated into 3 mL of standard growth medium in test tubes (YPD 2%) supplemented with 100 mg/L of ferulic acid or 100 mg/L of p-coumaric acid. The tubes were sealed with a plastic cork and incubated for five days at 30 °C, with shaking (120 rpm). After incubation, the contents of the test tubes were centrifuged (3 min, 3000 rpm), and 2 mL of supernatant was collected. Next, absorbance was measured at a 325 nm wavelength for ferulic acid and 285 nm for p-coumaric acid (Beckman DU-650 UV–Vis, Triad Scientific, Manasquan, NJ, USA). During the experiment, a positive control (Saccharomyces cerevisiae SafAle™ T-58, Fermentis, Rue Gabriel Péri, France) was included, as well as blank measurements (noninoculated YPD 2% + ferulic acid or p-coumaric acid medium). Strains were regarded as POF+ if the measured amount of ferulic acid or p-coumaric acid was below the 90% confidence interval of the blank.
2.2.4. β-Glucosidase Activity
The analysis was conducted according to Daenen et al. [22]. A loopful of yeast cells was inoculated from YPD slants to 5 mL YPD at pH 5 and incubated at 25 °C. Then, the yeast was inoculated at a cell concentration of 106 cells/mL in Erlenmeyer flasks filled to 20% of their volume with YPD and grown at 20 °C on an orbital shaker (120 rpm) until reaching the later exponential growth phase.
Extracellular activity
The yeast cell suspension (1 mL) was centrifuged (4 °C, 3000× g, 10 min) and 0.2 mL of supernatant was kept for the determination of extracellular activity on 4-Nitrophenyl β-D-glucopyranoside—pNPG.
Cell-associated activity
The remaining pellet was washed with 1 mL saline solution (sterile, cold, 7 g/L NaCl). After centrifugation (4 °C, 3000× g, 10 min) the supernatant was removed and 0.2 mL McIlvaine buffer pH 5 (0.1 mol/L citric acid and 0.2 mol/L Na2HPO4) was added to the pellet.
Enzyme assay on pNPG for determination of the extracellular and cell-associated activity
A total of 0.2 mL of pNPG solution (5 mmol/L) in McIlvaine buffer (pH 5) was added to 0.2 mL supernatant and to the pellet in 0.2 mL buffer. After incubation at 30 °C for 1 h, 0.8 mL carbonate buffer (pH 10.2) was added and the tubes were centrifuged (4 °C, 3000× g, 10 min). The increase in absorbance by the released pNP was measured spectrophotometrically at 400 nm (Beckman DU-650 UV–Vis). Blanks were prepared for the substrate and the different fractions.
One unit (U) of enzyme activity was defined as the quantity of enzyme that released 1 µmol of pNP per minute under the above experimental conditions, expressed per g of dry weight [23].
Yeast dry mass
The dry yeast mass was determined on a moisture analyzer Radwag MAC-50 (Zakład Mechaniki Produkcyjnej, Łódź, Poland).
2.2.5. β-Lyase Activity
The analysis was conducted according to Belda et al. [24]. The YCB-SMC medium was 0.1% (w/v) S-methyl-L-cysteine, 0.01% (w/v) pyridoxal-5′-phosphate, and 1.2% (w/v) Yeast Carbon Base. For solid medium, 2% agar was added. Potential errors due to nitrogen reserves were eliminated by making two serial transfers in the YCB-SMC liquid medium. When the first transfer was three days old, one loopful was transferred to a second test tube containing YCB-SMC medium containing the same source of nitrogen. After that, one loopful of medium was spread on a Petri dish with YCB-SMC-agar. Colony growth was checked after 72 h at 20 °C.
Interpretation of results:
- -: no growth,
- +: individual colonies grown on the plate,
- ++: many colonies grown on the plate.
2.3. Brewing Tests
2.3.1. Wort Preparation and Yeast Pitching
The wort (supplied by the Browar Górniczo-Hutniczy brewery—Kraków, Poland) used in the study had an extract content of 9.3%.
The propagation of yeast cultures was carried out according to Cioch-Skoneczny et al. [25]. Pure yeast cultures were passaged in triplicate. In the first stage, the strains were grown on the Sabouraud agar slants for 24 h at 28 °C. Then, the strains were transferred to 10 mL of Sabouraud Broth. After another 24 h, dynamic propagation of the strains was conducted in 200 mL of Sabouraud Broth on a water bath shaker (120 rpm, 28 °C) for 48 h. After that, the yeast concentration in the obtained yeast slurry was counted (using a Thoma cell counting chamber).
The pitching rate of yeasts was 106 cells per mL of wort per 1 °Plato. The adequate amount of growth media with yeasts was aseptically collected and centrifuged (5 min, 10,000× g). Sediment obtained from centrifugation was washed with sterile water and centrifuged again. Such cells were used in pitching. Then, 220 mL of beer wort and adequate amount of yeasts were added to a fermentation vessel, fitted with an airlock, and secured with a parafilm.
2.3.2. Course of Fermentation
The course of fermentation was measured as a function of weight loss of evaporating CO2 during the fermentation process. The masses of fermenting setups were measured daily, using the RADWAG WPS 600/C scale (Radwag Balances and Scales, Radom, Poland).
2.3.3. Alcohol, Real and Apparent Extract Content, Real and Apparent Attenuation, Caloric Content
The analysis was performed according to Ciosek et al. [26]. A DMA 4500 M Anton Paar beer analyzer was used to determine the alcohol content, real and apparent extract content, real and apparent attenuation, and caloric content. Before measurement, samples were degassed and centrifuged (3000× g, 5 min).
2.3.4. Color
The beer was filtered using a 0.22 μm syringe filter (Biosens, Warsaw, Poland). The color of the filtered beer was measured spectrophotometrically, using a Beckman DU-650 UV-Vis spectrophotometer at a wavelength of 430 nm, according to Analytica EBC method 8.5 Color of wort: Spectrophotometric Method (IM) [27].
2.3.5. Titratable Acidity and pH
The analysis was performed according to Cioch et al. [28]. The titratable acidity was determined by taking 25 mL of a degassed sample into a beaker. The sample’s pH was evaluated and later monitored using a pH meter. The sample was titrated with 0.1 M NaOH solution until it reached pH = 8.0. The total acidity was calculated as the amount of 1 M NaOH per 100 mL of a beer.
2.4. Statistical Analysis
The results are presented as an arithmetic mean of three repetitions, with standard deviation included. Additionally, a repeated-measures ANOVA and a Tukey’s (HSD) multiple range test at the significance level of α = 0.05 were performed. Analyses were performed using Statistica (StatSoft Poland, Kraków, Poland) software (v.13.3).
3. Results and Discussion
3.1. Microbial Analyses
3.1.1. Sugar Utilization
All of the examined yeast species were found to ferment glucose to varying degrees. Regarding the sucrose, most of the studied yeast showed the ability to at least partially ferment this sugar (Table 1).

Table 1.
Sugar utilization across the yeasts, phenolic off-flavors, and β-lyase screening results.
As stated in the literature data, M. pulcherrima strains should not be able to ferment maltose or sucrose [19]. The lack of maltose fermentation is consistent with the literature data. However, some authors, such as Drosou et al. [29] and Einfalt [30], have found strains of M. pulcherrima which are able to ferment maltose. Other authors, such as Postigo et al. [31], report a lack of maltose fermentation ability in these yeast species. It suggests that these yeasts are characterized by some interspecies variability regarding the utilization of this sugar. Regarding the sucrose, the yeast of Metschnikowia pulcherrima species was highly varied in this regard, where 6/9 of the strains fermented sucrose, which is not consistent with the data provided by Kurtzman et al. [19], where this species should not ferment this sugar. Of the H. uvarum species, one strain was found to ferment sucrose, and only to a low degree. According to the literature, H. uvarum species should not ferment sucrose [19]. Z. meyerae species are known to show variable utilization of sucrose, and maltose [32]. The studied strains utilized glucose, but did not utilize maltose. Lactose was not fermented by any of the yeast species, even by K. lactis (KL1) yeast strain. Not all of the K. lactis yeasts show the ability to ferment lactose. The genes responsible for that are LAC4 and LAC12, which encode β-galactosidase and lactose permease, which probably were missing or non-functional in the evaluated strain [33,34].
Overall, maltose was not fermented by any of the non-Saccharomyces yeast species studied, except WA1 (Table 1). In S. cerevisiae yeasts, maltose fermentation ability is linked to an active transport of this sugar via the plasma membrane, followed by its intracellular hydrolysis by maltases. The ability to ferment maltose is dependent on a series of unlinked telomere-associated MAL loci. However, there is very little known on the biochemical and genetic background of non-Saccharomyces yeast maltose fermentation ability [35].
In summary, the aim was to find yeast species which do not ferment maltose but can ferment glucose and saccharose. Such yeasts should ensure a beer with low levels of alcohol. The inclusion of lactose in the tests was mainly due to K. lactis strain, which was found to not ferment this sugar. This sugar is sometimes used as an additive in some types of beers [36].
3.1.2. Phenolic off Flavor Production
Ferulic and p-coumaric acids are the predominant phenolic acids in brewers’ wort. They are extracted from plant materials (primarily malt) during the brewing process. These compounds can be decarboxylated by yeasts with adequate enzymatic abilities to form 4-vinylguaicol and 4-vinylphenol, respectively. These compounds bestow a beer with clove, spice, smoky, phenolic, medicinal, and spicy aromas [37]. In S. cerevisiae, the ability to decarboxylase these compounds is regulated by the PAD1 and FDC1 genes [38]. Although the used US-05 yeasts were supposed to be a negative control and not produce phenolic off-flavor, the obtained results show that they were POF (+) for ferulic acid but not for p-coumaric acid (Table 1). This might have been caused by the high temperature of cultivation (30 °C), which is much higher than the range suggested by the manufacturer (18–26 °C) [39]. Overall, the POF test using ferulic acid was found to be positive for WA1 and WO1 and inconclusive for KL1 and NI1 (Table 1). W. anomalus was reported by other authors to produce these off-flavors. Larroque et al. [40] report a similar ability of Z. meryae, but the strains used in this study lacked the ability to produce these compounds.
Regarding p-coumaric acid, very few strains showed the ability to degrade this compound. It was found only for WO1 and WA1. These strains were also found to degrade ferulic acid. No ability to reduce this compound was found for control strain T-58 (Table 1). Similarly to the results obtained by Shinohara et al. [41], the ability to utilize p-coumaric acid was scarcer than for ferulic acid. Many wine-isolated yeast species typically have the ability to utilize these compounds [41,42].
3.1.3. Enzymatic Activities
β-lyase
Because the S-methyl-L-cysteine is the sole nitrogen source in the medium used, only yeasts which possess β-lyase activity should express the growth on the medium. To eliminate the influence of the possible nitrogen reserves in the cells, the yeasts were transferred three times on the medium [24].
Except for HU2 and some M. pulcherrima species, all of the studied yeasts showed mostly good growth on the studied YCB medium, which proves their β-lyase potential (Table 1). The yeast cultures used were isolated from a winery environment. According to Ruiz et al. [43], wine-related strains have low β-lyase potential. The obtained results show that this is not always the case. The M. pulcherrima species were characterized by high variability in the studied enzymatic activity. This variability was higher than in a study by Barbosa et al. [44], where most of the studied yeasts showed at least some degree of growth on the YCB-SMC medium. However, Zott et al. [45] found similar variability in β-lyase activity in the studied M. pulcherrima strains.
Interestingly, the control strain studied showed a very good growth on the medium (Table 1). The β-lyase activity of S. cerevisiae brewing yeast is corroborated by Krogerus et al. [46] and Michel et al. [47]. The β-lyase activity is, according to current knowledge, related to the ability of yeasts to release polyfunctional thiols from their cysteinated and glutathionylated precursors. These compounds are characterized by very low sensory threshold, and fruity, tropical aromas. The liberation of these compounds may improve the aroma of low-alcohol beers. The source of these compounds are malt and hops [48,49].
β-glucosidase activity
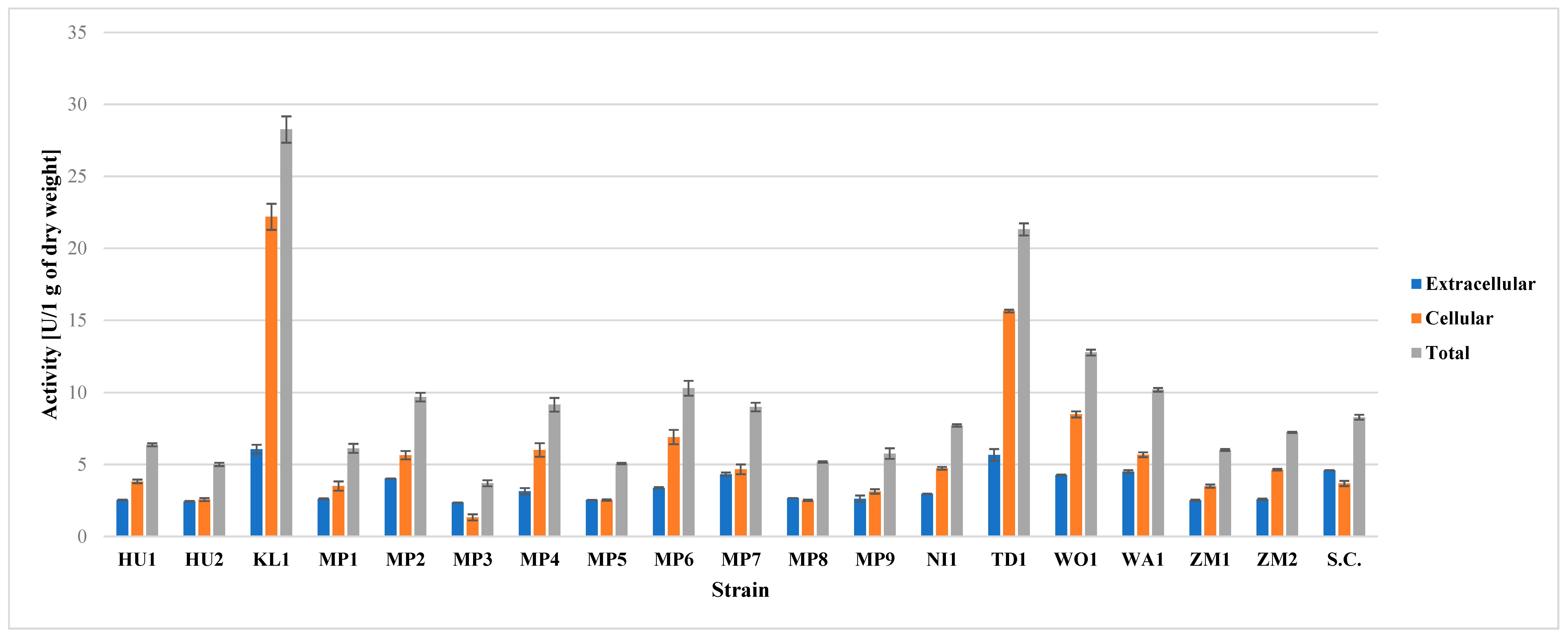
The highest degree of β-glucosidase activity was found in KL1 and TD1 (Figure 1). High potential of T. delbrueckii was also found by Schober et al. [50], whereas this is the first report of such high activity of this enzyme in K. lactis. High, but much lower activity than in two mentioned species was found in WO1. Other yeast species were found to have similar and moderate degree of this enzymatic activity (Figure 1). The moderate β-glucosidase potential of S. cerevisiae strains is confirmed by Daenen et al. [22] and Kanauchi and Bamforth [51]. All of the studied M. pulcherrima strains possessed this enzymatic activity, which is consistent with literature reports [52]. Some literature reports state that some strains of M. pulcherrima should be characterized by high enzymatic activity, which was not observed in this study [44].
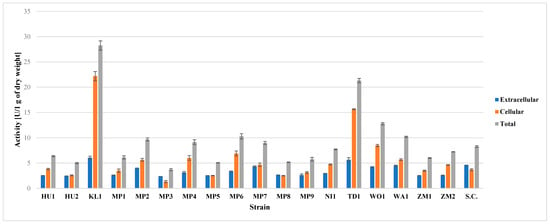
Figure 1.
Results of β-glucosidase activity assay on pNPG. Total activity is the sum of extracellular and cellular activity.
In all of the strains, except MP3 and US-05, the enzyme activity was mostly cell-bound (Figure 1). The occurrence of most of the β-glucosidase activity within the cell wall is corroborated by the other authors [22,53]. Similarly to β-lyase enzyme activity, β-glucosidase activity may enhance the aroma of a beer. According to the current knowledge, β-glucosidase is at least partially responsible for the liberation of aroma-active aglycones from their glycoside-bound aroma substances. The action of this enzyme might enhance the fruity and floral aromas in a beer [54].
3.1.4. Resistance to Stress Factors
The growth of nearly all of the studied yeasts was unaffected by the iso-α-acids, even at the higher concentration studied (90 IBU). The only exception was NI1 yeasts. However, the higher concentration studied did not arrested the growth of the yeast further (Table 2). Yang et al. [55] reports the inhibited growth of wild S. cerevisiae strain by iso-a-acids, although at a very high concentration (3 g/L). The IBU value of most of the beers on the market ranges from 1 to about 100 IBU’s. Most beers have an IBU value in the lower range, with the exception of some highly hopped double IPA’s or American barley wines [36]. Iso-α-acids are known to not affect the growth and functioning of S. cerevisiae yeast, which is related to many adaptations of these yeasts to weak acids. The studied yeast probably possesses some adaptations to weak acids as well [56,57]. The resistance to iso-α-acids is a requirement for yeasts in beer production [17].
Table 2.
Resistance to stress factors.
Most of the studied strains managed to grow in an environment of up to 4.5% ethanol. The exceptions were MP4, WO1, TD1, WI1, WA1, and KA1. The only non-Saccharomyces strain that grew at 7.5% was WA1. S. cerevisiae, expectedly, grew across all the studied alcohol ranges (Table 2).
The studied M. pulcherrima strains were characterized by lower tolerance to ethanol than in a study by Barbosa et al. [44], where most of the strains managed to tolerate at least 6% ethanol. However, the obtained results are in line with the studies of other authors, who found that M. pulcherrima survives up to 4–5% ethanol [58]. All of the studied yeasts showed a satisfactory tolerance to ethanol in the environment.
3.2. Fermentative Analyses
3.2.1. Fermentation Course
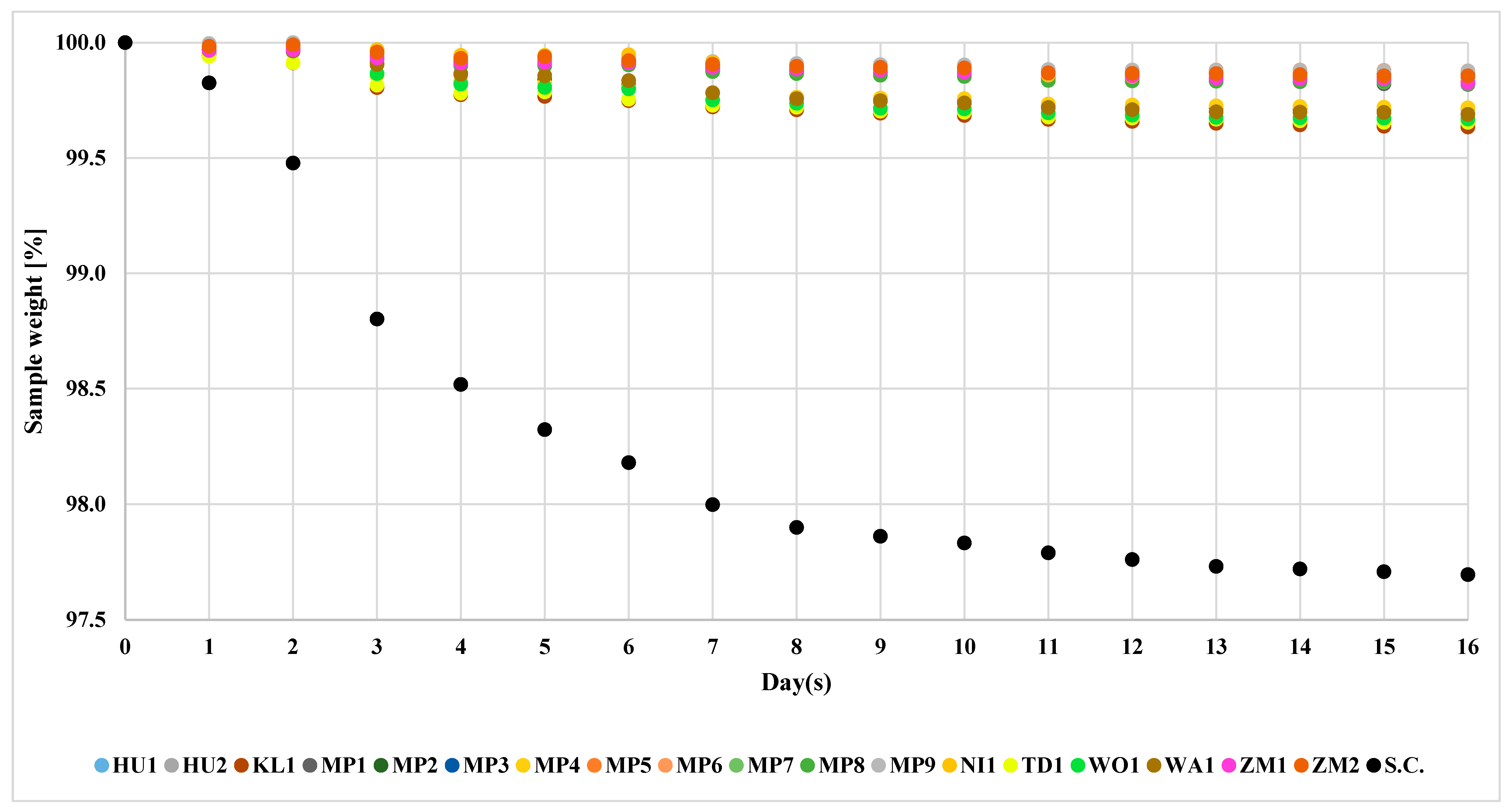
The fermentation process was conducted over 17 days. Control strain, S. cerevisiae US-05, conducted the fermentation at the fastest pace among the studied strains. The final mass amounted to 97.8% of the initial mass (Figure 2).
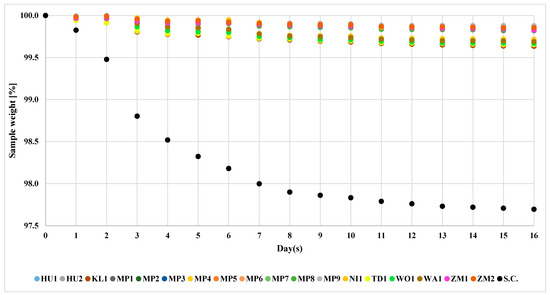
Figure 2.
Fermentation course of S. cerevisiae US-05 and non-Saccharomyces yeasts. The standard deviation of individual measurements is below 0.5%.
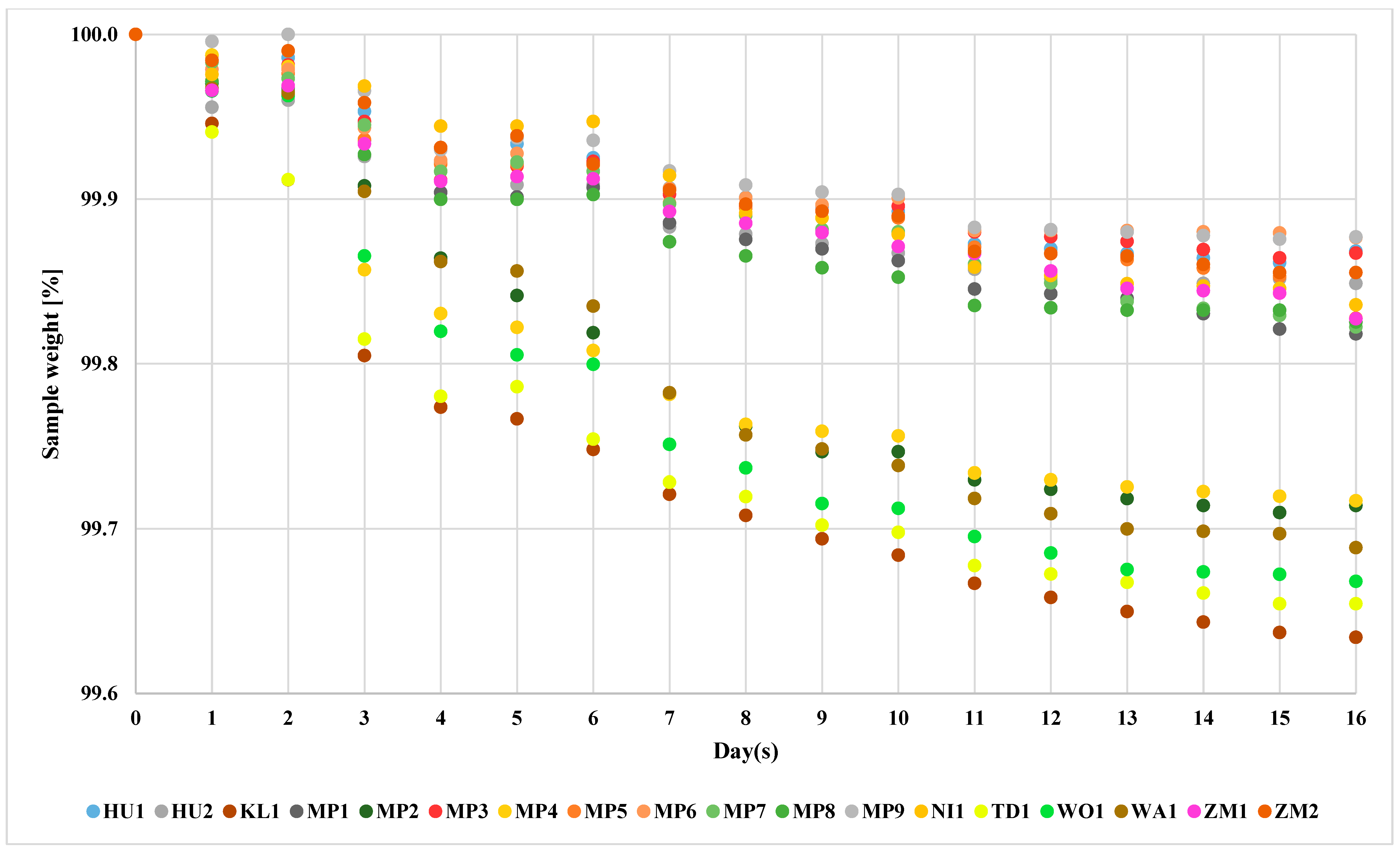
The non-Saccharomyces yeast species used much lower amounts of sugars during the fermentation (Figure 3). At the third day of the fermentation, the yeasts divided into two groups. That divergence remained till the end of the fermentation process. The yeasts KL1, MP2, MP4, TD1, WA1 and WO1 showed higher loss of the mass. The remaining yeasts showed lower loss. Interestingly, the strain WA1 was found to conduct the fermentation process similarly to other non-Saccharomyces yeasts, despite the results suggesting its ability to ferment maltose (Table 1). It is then possible that the identified ability to ferment maltose was a false-positive result. This result might be related to the different amino acid compositions of brewer’s wort and YPD medium, which is known to influence the yeast’s ability to utilize sugars [59].
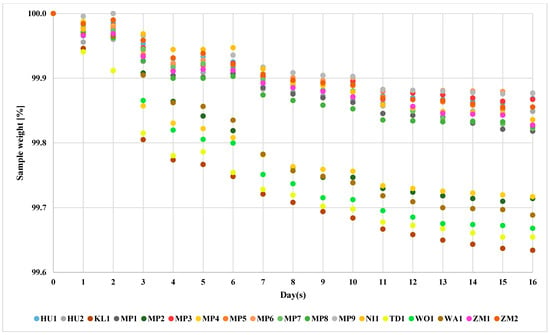
Figure 3.
Fermentation course of non-Saccharomyces yeasts. The standard deviation of individual measurements is below 0.5%.
3.2.2. Beer and Wort Physicochemical Characteristics
The non-Saccharomyces yeast species produced beers with the alcohol content in the range of 0.33–0.83% (v/v). The control strain of S. cerevisiae US-05 produced significantly higher level of alcohol (3.61% (v/v)). Based on the results, the non-Saccharomyces yeast probably did not utilize the maltose or maltotriose. Most of the studied strains, with the exception of KL1, MP2, MP4, TD1, WO1, and WA1, produced less than 0.5% (v/v) ethanol (Table 3). These are the same strains which showed lower losses of mass during the fermentation process. Looking at the alcohol content, along with the sugar fermentation tests (Table 1), all of the strains show potential in obtaining a no-alcohol beer (<0.5%) without any significant process modifications. In case of using the strains that reach higher attenuation, a wort of lower extract might be necessary. Comparing with the other authors, Einfalt [30] obtained a beer with significantly higher alcohol content from 12.5 °Plato wort. The strains evaluated by the author, that is, M. pulcherrima, produced 33.8 g/L of alcohol, whereas the T. delbrueckii strain produced 29.2 g/L of alcohol [30]. Low alcohol levels (~1% v/v) in a beer obtained from 11.1 °Plato wort were found by Postigo et al. [31] for W. anomalus, T. delbrueckii, and M. pulcherrima yeast strains. Similarly, Canonico et al. [60] reports 1.53% (v/v) beer from 12.3 °Plato wort fermented by W. anomalus. The obtained results indicate that wine-related non-Saccharomyces strains may be of particular interest in a screening of maltose-negative yeasts.
Table 3.
Physicochemical characterization of beers and wort (mean of three replicates ± standard deviation).
Regarding the attenuation rate, the control strain US-05 had an apparent attenuation of 81.8%, which is consistent with the manufacturer’s data [39]. All of the non-Saccharomyces yeasts had attenuation in the range of 7.68–17.5% (Table 3). This further confirms that the studied yeasts did not utilize maltose nor maltotriose. Such attenuation rates are in line with the attenuation range of low- and no-alcohol yeasts found on the market.
The real extract of the beers obtained with non-Saccharomyces yeasts was expectedly high, in the range of 7.35–8.46 (w/w) (Table 3). Such a high content of residual extract is typical for low-alcohol beers obtained with biological methods [5]. The yeasts used about 1.0–1.5 °Plato of the extract (Table 3).
The color of most of the samples did not differ significantly from the color of the reference sample (S.C.). The only exception was a beer produced with the MP6 strain, which had the deepest color among the evaluated samples. This may have been related to the unique ability of some of the M. pulcherrima strains to produce a red dye, called pulcherrimin [61]. The color of all of the beers was lighter compared to wort (Table 3). Lightening of a wort after a fermentation is typical [62].
Typically, the beer should have a pH of about 4.0–4.5 [63]. No-alcohol beers produced using microbiological methods are often characterized by excessively high pH levels [15]. The low pH is one of the microbiological hurdles which make a beer a microbiologically safe product. Munford et al. [64] states that the lack of microbial growth in studied beer samples can be attributed to the low pH (below 4.5) of evaluated beers. The low pH also makes the beer more refreshing and hides unwanted off-flavors. Therefore, no-lo beers are often acidified to amend their flavor [15,65]. In that regard, yeast strains could produce a beer with relatively low pH, such as MP1, MP5, MP9, and NI1, are especially interesting. The titratable acidity of beers was in a range of 1.18–3.64 mL 1 M NaOH/100 mL of beer. The acidity of the beers did not differ significantly between the samples (Table 3). The beer obtained by Wzorek et al. [66] from 12.5 °Blg wort had an acidity of 2.50 mL NaOH/100 mL of beer.
3.3. Limitations and Insights of a Study
The studied non-Saccharomyces yeasts may offer several benefits for the production of low-alcohol beers. Due to low ethanol production, the studied yeast strains may be used as primary fermenters in beer production, which would simplify the production of such beverages. Physical dealcoholization processes inherently remove volatile compounds from a beer, which negatively impacts its flavor. Directly obtaining a beer with a lower alcohol content might allow brewers to retain more of the flavor compounds in a finished beer. Some of the studied strains (such as TD1 or KL1) are characterized by higher enzymatic activities regarding the biotransformation. If such a yeast would allow more of the flavor precursors to be liberated in the fermentation conditions, they might allow brewers to obtain a beer with richer flavor. Additionally, the strains with high bioflavoring potential might be used in co-fermentation with brewing strains of S. cerevisiae to obtain higher-alcohol beers with richer aromas.
The main limitation of this study is the limited physicochemical analysis of the obtained beers. The study focused mainly on the biological analysis of the yeast strains. To evaluate the potential of the yeasts studied, a broader analysis of beers, such as sensory analysis, the composition of volatile compounds, acids, and sugars is required. However, such approach was by design. In a continuation of this research, a broader analysis focusing only on the physiochemical parameters and sensory characteristics of the obtained beers is planned. Additionally, if any of the studied strains were to be used in the production of beer, significant process parameters (fermentation temperature and time, pitching rate, etc.) would be required.
4. Conclusions
The evaluated yeast species isolated from a winery environment are especially interesting in the production of reduced-alcohol and no-alcohol beers, as they do not utilize maltose. It naturally restricts the levels of alcohol in resulting beers. Most of the strains did not produce phenolic off-flavors and allowed us to obtain a beer with <0.5% alcohol by volume. The lack of phenolic off-flavor production is important, as the aromas bestowed by these compounds are mostly seen as disadvantage. Two of the evaluated strains, K. lactis MG971263 and T. delbrueckii MG971248, were characterized by a high β-glucosidase activity, which might potentially positively influence the aroma of the resulting beers. Additionally, the yeasts were resistant to low levels of alcohol in the environment and reduced the pH of a wort during a fermentation. Further studies focusing on beer physicochemical and sensory characteristics are required.
Author Contributions
Conceptualization, K.K. and M.C.-S.; methodology, K.K.; software, K.K.; validation, K.K., M.C.-S. and A.P.; formal analysis, K.K. and M.C.-S.; investigation, K.K.; resources, K.K., M.C.-S. and A.P.; data curation, K.K.; writing—original draft preparation, K.K.; writing—review and editing, K.K., M.C.-S. and A.P.; visualization, K.K.; supervision, M.C.-S. and A.P.; project administration, K.K.; funding acquisition, K.K. All authors have read and agreed to the published version of the manuscript.
Funding
The research was partially founded by Agricultural University of Kraków grant number AD-50.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Garavaglia, C.; Swinnen, J. Economics of the Craft Beer Revolution: A Comparative International Perspective. In Economic Perspectives on Craft Beer; Springer International Publishing: Cham, Switzerland, 2018; pp. 3–51. [Google Scholar]
- Anderson, K. The Emergence of Lower-Alcohol Beverages: The Case of Beer. J. Wine Econ. 2023, 18, 66–86. [Google Scholar] [CrossRef]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Kokole, D.; Jané Llopis, E.; Anderson, P. Non-alcoholic Beer in the European Union and UK: Availability and Apparent Consumption. Drug Alcohol. Rev. 2022, 41, 550–560. [Google Scholar] [CrossRef]
- Brányik, T.; Silva, D.P.; Baszczyňski, M.; Lehnert, R.; Almeida E Silva, J.B. A Review of Methods of Low Alcohol and Alcohol-Free Beer Production. J. Food Eng. 2012, 108, 493–506. [Google Scholar] [CrossRef]
- Parapouli, M.; Vasileiadi, A.; Afendra, A.-S.; Hatziloukas, E. Saccharomyces cerevisiae and Its Industrial Applications. AIMS Microbiol. 2020, 6, 1–32. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could Non-Saccharomyces Yeasts Contribute on Innovative Brewing Fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by Non-Conventional Yeasts in Sequential Beer Fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Buiatti, S.; Tat, L.; Natolino, A.; Passaghe, P. Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review. Fermentation 2023, 9, 327. [Google Scholar] [CrossRef]
- Tolosa, J.J.M.; Prieto, S.M. Non-Saccharomyces Yeasts: An Enzymatic Unexplored World to Be Exploited. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 433–450. [Google Scholar]
- Svedlund, N.; Evering, S.; Gibson, B.; Krogerus, K. Fruits of Their Labour: Biotransformation Reactions of Yeasts during Brewery Fermentation. Appl. Microbiol. Biotechnol. 2022, 106, 4929–4944. [Google Scholar] [CrossRef]
- Palmer, J.J. How to Brew: Everything You Need to Know to Brew Great Beer Every Time, 4th ed.; Kristi Switzer: Gilbert, AZ, USA, 2017. [Google Scholar]
- Magalhães, F.; Vidgren, V.; Ruohonen, L.; Gibson, B. Maltose and Maltotriose Utilisation by Group I Strains of the Hybrid Lager Yeast Saccharomyces pastorianus. FEMS Yeast Res. 2016, 16, fow053. [Google Scholar] [CrossRef]
- Preedy, V.R. (Ed.) Beer in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Methner, Y.; Hutzler, M.; Matoulková, D.; Jacob, F.; Michel, M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation 2019, 5, 101. [Google Scholar] [CrossRef]
- Yabaci Karaoglan, S.; Jung, R.; Gauthier, M.; Kinčl, T.; Dostálek, P. Maltose-Negative Yeast in Non-Alcoholic and Low-Alcoholic Beer Production. Fermentation 2022, 8, 273. [Google Scholar] [CrossRef]
- Jimoh, S.O.; Ado, S.A.; Ameh, J.A.; Whong, C.M.Z. Osmotolerance and Fermentative Pattern of Brewer’s Yeast. World J. Life Sci. Med. Res. 2012, 2, 59–64. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W.; Teun, B. (Eds.) The Yeasts. A Taxonomic Study, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Hong, M.; Li, J.; Chen, Y. Characterization of Tolerance and Multi-Enzyme Activities in Non-Saccharomyces Yeasts Isolated from Vidal Blanc Icewine Fermentation. J. Food Biochem. 2019, 43, e13027. [Google Scholar] [CrossRef]
- Mertens, S.; Steensels, J.; Gallone, B.; Souffriau, B.; Malcorps, P.; Verstrepen, K.J. Rapid Screening Method for Phenolic Off-Flavor (POF) Production in Yeast. J. Am. Soc. Brew. Chem. 2017, 75, 318–323. [Google Scholar] [CrossRef]
- Daenen, L.; Saison, D.; Sterckx, F.; Delvaux, F.R.; Verachtert, H.; Derdelinckx, G. Screening and Evaluation of the Glucoside Hydrolase Activity in Saccharomyces and Brettanomyces Brewing Yeasts. J. Appl. Microbiol. 2008, 104, 478–488. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.A.; Van Broock, M.; Valles, S.; Ramón, D.; Caballero, A.C. Screening and Typing of Patagonian Wine Yeasts for Glycosidase Activities. Proc. J. Appl. Microbiol. 2004, 96, 84–95. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of Aromatic Thiol Release through the Selection of Yeasts with Increased β-Lyase Activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- Cioch-Skoneczny, M.; Cichoń, N.; Satora, P.; Skoneczny, S. Physicochemical Characteristics of Beer with Grape Must Addition Produced Using Non-Saccharomyces Yeasts. Eur. Food Res. Technol. 2023, 249, 903–912. [Google Scholar] [CrossRef]
- Ciosek, A.; Rusiecka, I.; Poreda, A. Sour Beer Production: Impact of Pitching Sequence of Yeast and Lactic Acid Bacteria. J. Inst. Brew. 2020, 126, 53–58. [Google Scholar] [CrossRef]
- European Brewery Convention (1998–2007) Analytica-EBC; Fachverlag Hans Carl: Nürberg, Germany, 2007.
- Cioch-Skoneczny, M.; Bajerski, M.; Klimczak, K.; Satora, P.; Skoneczny, S. Influence of Oak Chips Addition on the Physicochemical Properties of Beer. Eur. Food Res. Technol. 2023, 249, 183–197. [Google Scholar] [CrossRef]
- Drosou, F.; Mamma, D.; Tataridis, P.; Dourtoglou, V.; Oreopoulou, V. Metschnikowia pulcherrima in Mono or Co-Fermentations in Brewing. BrewingScience 2022, 75, 69–78. [Google Scholar] [CrossRef]
- Einfalt, D. Barley-Sorghum Craft Beer Production with Saccharomyces cerevisiae, Torulaspora delbrueckii and Metschnikowia pulcherrima Yeast Strains. Eur. Food Res. Technol. 2021, 247, 385–393. [Google Scholar] [CrossRef]
- Postigo, V.; Sanz, P.; García, M.; Arroyo, T. Impact of Non-Saccharomyces Wine Yeast Strains on Improving Healthy Characteristics and the Sensory Profile of Beer in Sequential Fermentation. Foods 2022, 11, 2029. [Google Scholar] [CrossRef]
- Smith, M.T.; Robert, V.; Poot, G.A.; Epping, W.; de Cock, A.W.A.M. Taxonomy and Phylogeny of the Ascomycetous Yeast Genus Zygoascus, with Proposal of Zygoascus meyerae sp. Nov. and Related Anamorphic Varieties. Int. J. Syst. Evol. Microbiol. 2005, 55, 1353–1363. [Google Scholar] [CrossRef]
- Breunig, K.D.; Bolotin–Fukuhara, M.; Bianchi, M.M.; Bourgarel, D.; Falcone, C.; Ferrero, I.; Frontali, L.; Goffrini, P.; Krijger, J.J.; Mazzoni, C.; et al. Regulation of Primary Carbon Metabolism in Kluyveromyces lactis. Enzym. Microb. Technol. 2000, 26, 771–780. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Barrio, E.; Querol, A. Genetic and Molecular Study of the Inability of the Yeast Kluyveromyces lactis Var. Drosophilarum to Ferment Lactose. Microbiology 2006, 75, 248–252. [Google Scholar] [CrossRef]
- Alves-Jr, S.L.; Herberts, R.A.; Hollatz, C.; Miletti, L.C.; Stambuk, B.U. Maltose and Maltotriose Active Transport and Fermentation by Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2007, 65, 99–104. [Google Scholar] [CrossRef]
- Oliver, G.; Colicchio, T. The Oxford Companion to Beer-Oxford University Press, 1st ed.; Oliver, G., Ed.; Oxford University Press Inc.: Oxford, UK, 2012. [Google Scholar]
- Lentz, M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation 2018, 4, 20. [Google Scholar] [CrossRef]
- Mukai, N.; Masaki, K.; Fujii, T.; Kawamukai, M.; Iefuji, H. PAD1 and FDC1 Are Essential for the Decarboxylation of Phenylacrylic Acids in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2010, 109, 564–569. [Google Scholar] [CrossRef]
- Fermentis SafAleTM US-05—Versatile Brewer’s Yeast for Neutral Ales—Fermentis. Available online: https://fermentis.com/en/product/safale-us%e2%80%9105/ (accessed on 2 August 2023).
- Larroque, M.N.; Carrau, F.; Fariña, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and Non-Saccharomyces Native Yeasts on Beer Aroma Compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef]
- Shinohara, T.; Kubodera, S.; Yanagida, F. Distribution of Phenolic Yeasts and Production of Phenolic Off-Flavors in Wine Fermentation. J. Biosci. Bioeng. 2000, 90, 90–97. [Google Scholar] [CrossRef]
- Fernández-González, M.; Briones, A.I. Study of Yeast Populations and Their Enological Properties in Guijoso Appellation of Origin (Spain). Ann. Microbiol. 2013, 63, 371–379. [Google Scholar] [CrossRef]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical Impact of Metschnikowia pulcherrima in the Volatile Profile of Verdejo White Wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and Phenotypic Characterization of Metschnikowia pulcherrima Strains from Douro Wine Region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The Grape Must Non-Saccharomyces Microbial Community: Impact on Volatile Thiol Release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef]
- Krogerus, K.; Rettberg, N.; Gibson, B. Increased Volatile Thiol Release during Beer Fermentation Using Constructed Interspecies Yeast Hybrids. Eur. Food Res. Technol. 2023, 249, 55–69. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Stretz, D. Screening of Brewing Yeast β-Lyase Activity and Release of Hop Volatile Thiols from Precursors during Fermentation. BrewingScience 2019, 72, 179–186. [Google Scholar] [CrossRef]
- Simon, M.; Christiaens, R.; Janssens, P.; Collin, S. Unexpected Behavior of a Maltose-Negative Saccharomyces cerevisiae Yeast: Higher Release of Polyfunctional Thiols from Glutathionylated Than from Cysteinylated S-Conjugates. Fermentation 2024, 10, 276. [Google Scholar] [CrossRef]
- Chenot, C.; Donck, W.; Janssens, P.; Collin, S. Malt and Hop as Sources of Thiol S-Conjugates: Thiol-Releasing Property of Lager Yeast during Fermentation. J. Agric. Food Chem. 2022, 70, 3272–3279. [Google Scholar] [CrossRef]
- Schober, D.; Wacker, M.; Schmarr, H.-G.; Fischer, U. Understanding the Contribution of Co-Fermenting Non-Saccharomyces and Saccharomyces Yeasts to Aroma Precursor Degradation and Formation of Sensory Profiles in Wine Using a Model System. Fermentation 2023, 9, 931. [Google Scholar] [CrossRef]
- Kanauchi, M.; Bamforth, C.W. β-Glucoside Hydrolyzing Enzymes from Ale and Lager Strains of Brewing Yeast. Cerevisia 2013, 38, 60. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of Non-Saccharomyces Yeasts with Enzymatic Activities of Interest in Wine-Making. Int. J. Food Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, F.; Yan, X.; Qin, Y.; Jiang, J.; Liu, Y.; Song, Y. Characterization of the β-Glucosidase Activity in Indigenous Yeast Isolated from Wine Regions in China. J. Food Sci. 2021, 86, 2327–2345. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Qin, Q.; Li, C.; Zhao, X.; Song, F.; An, M.; Chen, Y.; Wang, X.; Huang, W.; Zhan, J.; et al. Application of Non-Saccharomyces Yeasts with High β-Glucosidase Activity to Enhance Terpene-Related Floral Flavor in Craft Beer. Food Chem. 2023, 404, 134726. [Google Scholar] [CrossRef]
- Yang, G.; Nie, C.; Zhang, H.; Sun, S.; Wang, X.; Zhang, J.; Xu, H.; Liu, J. The Tolerance of Brewing-Related Microorganisms to Isomerized Hop Products and the Effect on Beer Stability and Quality. Eur. Food Res. Technol. 2021, 247, 555–567. [Google Scholar] [CrossRef]
- Mira, N.P.; Teixeira, M.C.; Sá-Correia, I. Adaptive Response and Tolerance to Weak Acids in Saccharomyces cerevisiae: A Genome-Wide View. OMICS 2010, 14, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Walsh, M.C.; Pronk, J.T.; Daran, J.-M. Involvement of Vacuolar Sequestration and Active Transport in Tolerance of Saccharomyces cerevisiae to Hop Iso-α-Acids. Appl. Environ. Microbiol. 2010, 76, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The Genus Metschnikowia in Enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Batistote, M.; Cruz, S.H.; Ernandes, J.R. Altered Patterns of Maltose and Glucose Fermentation by Brewing and Wine Yeasts Influenced by the Complexity of Nitrogen Source. J. Inst. Brew. 2006, 112, 84–91. [Google Scholar] [CrossRef]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of Three Non-Conventional Yeast Species in the Brewing Process. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, E.; Kolesińska, B.; Nowacka, M.; Kregiel, D. A New Approach to Producing High Yields of Pulcherrimin from Metschnikowia Yeasts. Fermentation 2020, 6, 114. [Google Scholar] [CrossRef]
- Narziß, L.; Back, W.; Gastl, M.; Zarnkow, M. Applied Malting and Brewing Science: A Weihenstephan Compendium; Wiley: Hoboken, NJ, USA, 2023. [Google Scholar]
- Krogerus, K.; Gibson, B.R. 125th Anniversary Review: Diacetyl and Its Control during Brewery Fermentation. J. Inst. Brew. 2013, 119, 86–97. [Google Scholar] [CrossRef]
- Munford, A.R.G.; Alvarenga, V.O.; do Prado-Silva, L.; Crucello, A.; Campagnollo, F.B.; Chaves, R.D.; Oteiza, J.M.; Sant’Ana, A.S. Sporeforming Bacteria in Beer: Occurrence, Diversity, Presence of Hop Resistance Genes and Fate in Alcohol-Free and Lager Beers. Food Control 2017, 81, 126–136. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Hutzler, M.; Zarnkow, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Investigation into the Potential of Lachancea fermentati Strain KBI 12.1 for Low Alcohol Beer Brewing. J. Am. Soc. Brew. Chem. 2019, 77, 157–169. [Google Scholar] [CrossRef]
- Wzorek, W.; Haberowa, H.; Jędrysiak, P. Zastosowanie Chitozanu w Kombinacji z Innymi Preparatami Do Stabilizacji Piwa Jasnego. Żywność Nauka Technol. Jakość 2001, 8, 43–56. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).