

Upgrading In Vitro Digestion Protocols with Absorption Models


Abstract
:1. Introduction
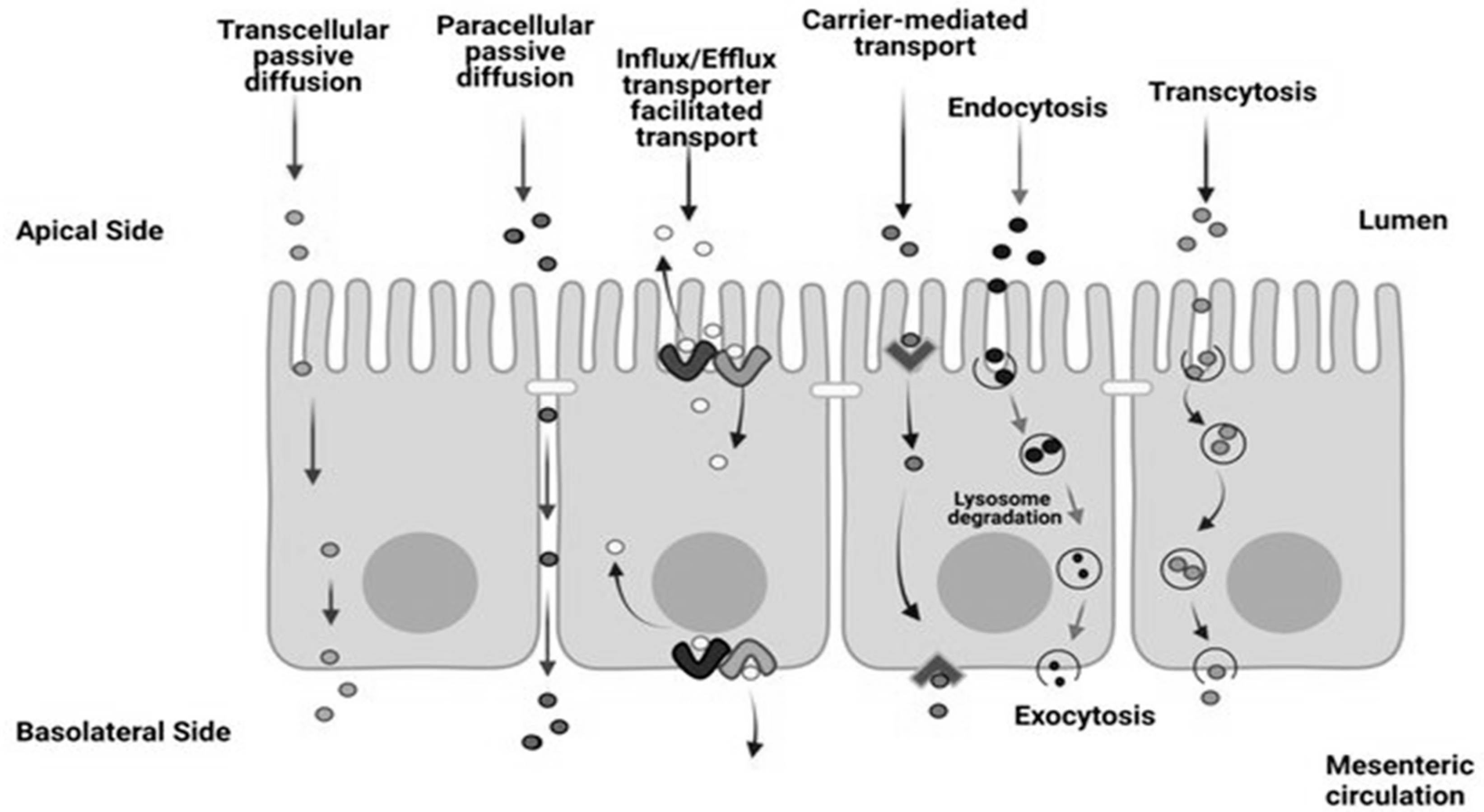
1.1. Description of Intestinal Epithelium
1.2. Challenges Regarding the Design of Intestinal Epithelium Models Used for Absorption Studies
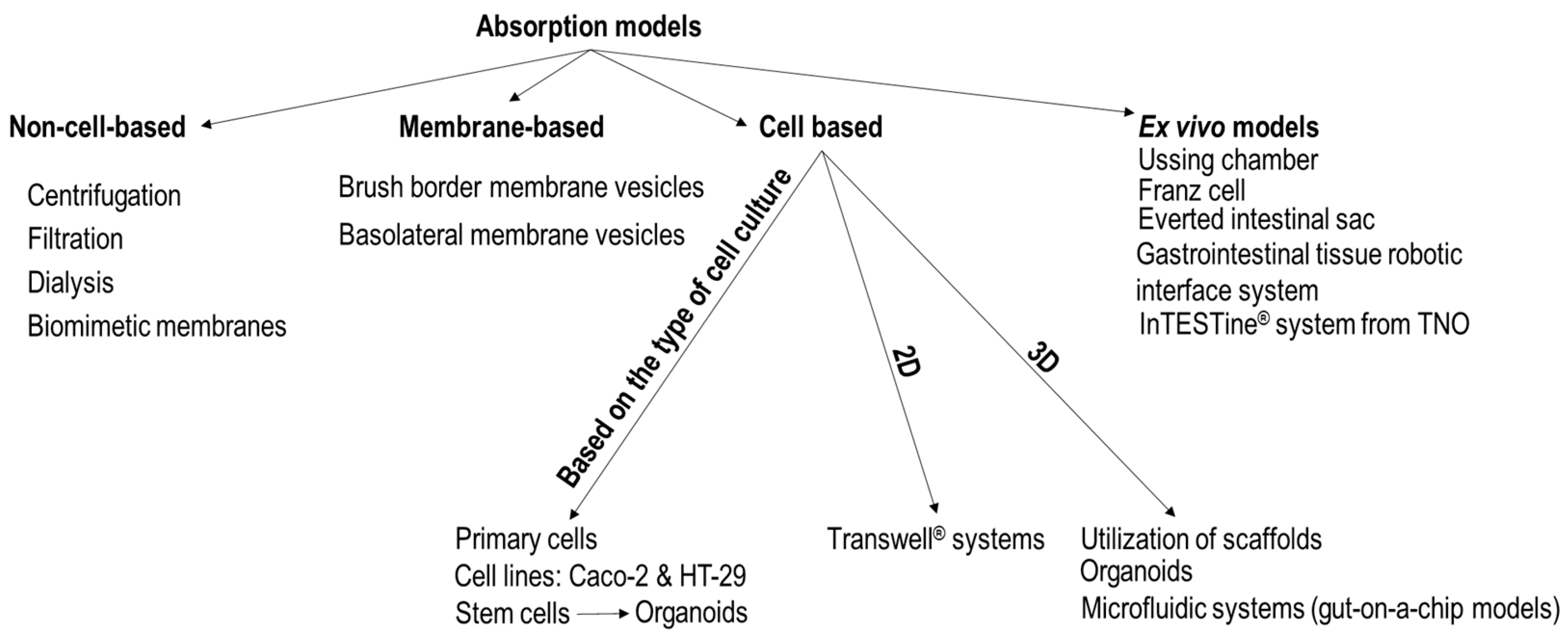
2. Technological Approaches of Intestinal Epithelium Models
2.1. Non-Cell-Based Transport Models
- parallel artificial membrane permeability (PAMPA): a filter is infused with an organic solvent that contains phospholipids, in a manner that it imitates the lipid composition of the intestinal membrane [22].
- vesicle-based permeation assay (PVPA): liposomes are deposited in the pores and on the surface of the filter [22].
- PermeaPad®: two cellulose-hydrate membranes act as support layers in a sandwich-like structure, where the middle layer is made of dry phospholipids. This structure hinders the erosion of the middle layer and prevents the leakage of lipids into the solutions used during the experiment [22].
2.2. Membrane-Based Transport Models
- (i)
- “Tris disruption procedure, followed by density gradient centrifugation”;
- (ii)
- the hypotonic EDTA procedure which can be complemented with density gradient centrifugations;
- (iii)
- the Ca2+ precipitation protocol. In the presence of Ca2+, endoplasmic reticulum and mitochondria are converted to larger particles due to aggregation. These larger particles can be easily separated by centrifugation at 2000× g. Following this, the brush border fragments are separated by centrifugation of the supernatant at 20,000× g [38];
- (iv)
- “isotonic homogenization and density gradient centrifugation”;
- (v)
- “isolation of enterocytes, homogenization and density gradient centrifugation or free flow electrophoresis”.
2.3. Cell Cultures Used in Epithelium Models
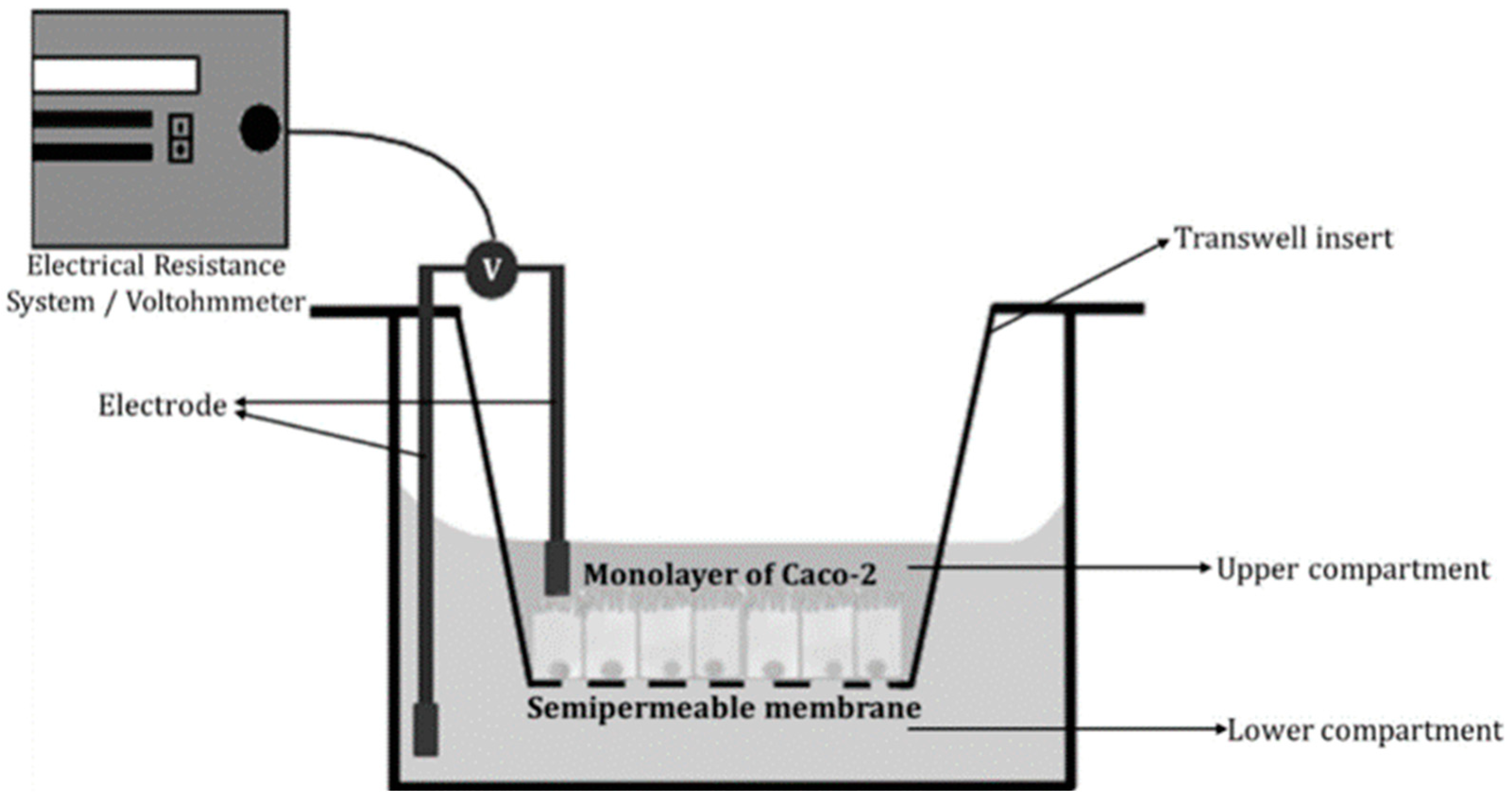
2.3.1. Transwell® Systems
2.3.2. Verification of Cell-Based Absorption Models
2.3.3. Utilization of Scaffolds
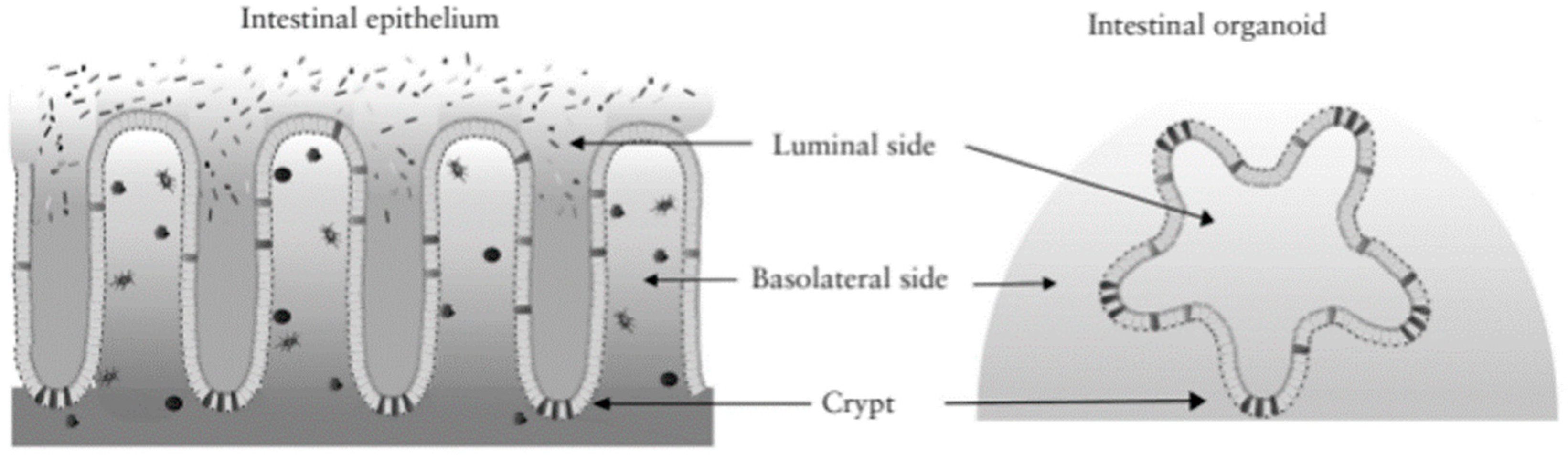
2.3.4. Organoid Cultures
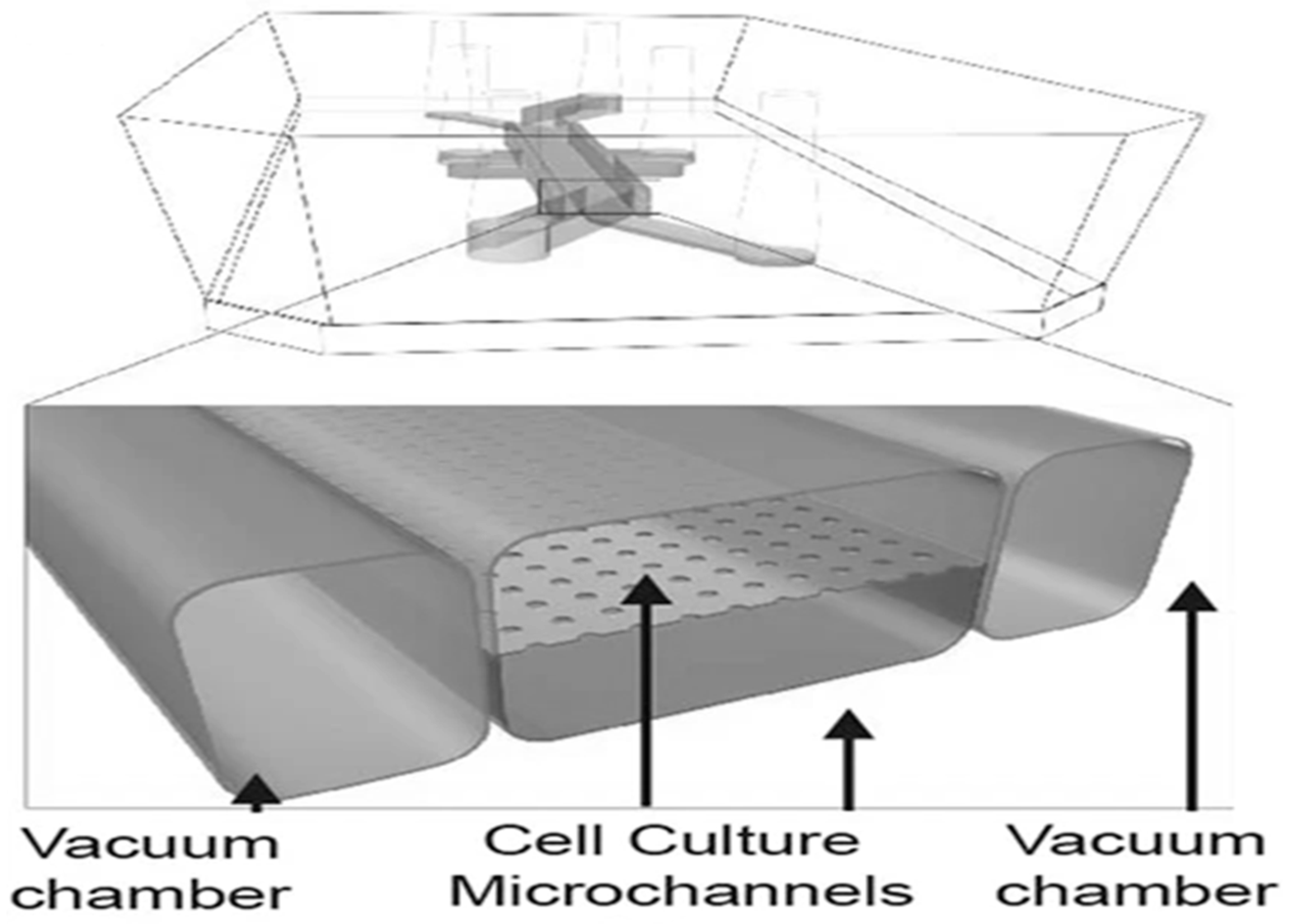
2.3.5. Microfluidic Systems That May Be Used in Absorption Studies
2.4. Ex Vivo Models
2.5. Advantages and Disadvantages of Models of the Intestinal Barrier
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Verhoeckx, K.; Cotter, P.; López-Expósito, I.; Kleiveland, C.; Lea, T.; Mackie, A.; Requena, T.; Swiatecka, D.; Wichers, H. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland; Berlin/Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2015. [Google Scholar]
- Antal, O.; Némethné Szerdahelyi, E.; Takács, K. In vitro humán emésztési modellek alkalmazása a táplálkozástudomány területén. (Appl. Vitr. Hum. Dig. Models Int He Field Nutr. Sci.) Élelmiszervizsgálati Közlemények J. Food Investig. 2020, 66, 31–57. [Google Scholar]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Tormási, J.; Abrankó, L. Lipid digestibility of sour cream and its analogue during in vitro digestion simulation and co-consumption with cooked pasta. Int. J. Food Sci. Technol. 2023, 58, 6330–6341. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, X.; Shang, Y.; Ding, Y. Microfluidic intestine-on-a-chip: Current progress and further perspectives of probiotic-foodborne pathogen interactions. Trends Food Sci. Technol. 2023, 134, 207–221. [Google Scholar] [CrossRef]
- Haddad, M.J.; Sztupecki, W.; Delayre-Orthez, C.; Rhazi, L.; Barbezier, N.; Depeint, F.; Anton, P.M. Complexification of in vitro models of intestinal barriers, a true challenge for a more accurate alternative approach. Int. J. Mol. Sci. 2023, 244, 3595. [Google Scholar] [CrossRef]
- Kondrashina, A.; Arranz, E.; Cilla, A.; Faria, M.A.; Santos-Hernández, M.; Miralles, B.; Hashemi, N.; Rasmussen, M.K.; Young, J.F.; Barberá, R.; et al. Coupling in vitro food digestion with in vitro epithelial absorption; recommendations for biocompatibility. Crit. Rev. Food Sci. Nutr. 2023, 26, 1–19. [Google Scholar] [CrossRef]
- Dutton, J.S.; Hinman, S.S.; Kim, R.; Wang, Y.; Allbritton, N.L. Primary cell-derived intestinal models: Recapitul. Physiol. Trends Biotechnol. 2019, 377, 744–760. [Google Scholar] [CrossRef]
- Grzymajlo, K. The game for three: Salmonella-host-microbiota interaction models. Front. Microbiol. 2022, 13, 854112. [Google Scholar] [CrossRef]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Pantoja-Feliciano, I.G.; Zachos, N.C.; Racicot, K. Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front. Physiol. 2018, 9, 1584. [Google Scholar] [CrossRef]
- Lechanteur, A.; das Neves, J.; Sarmento, B. The role of mucus in cell-based models used to screen mucosal drug delivery. Advandced Drug Deliv. Rev. 2018, 124, 50–63. [Google Scholar] [CrossRef]
- Fedi, A.; Vitale, C.; Ponschin, G.; Ayehunie, S.; Fato, M.; Scaglione, S. In vitro models replicating the human intestinal epithelium for absorption and metabolism studies: A systematic review. J. Control Release 2021, 335, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Suzuki, K.; Sifuentes-Dominguez, L.; Miyata, N.; Song, J.; Lopez, A.; Starokadomskyy, P.; Gopal, P.; Dozmorov, I.; Tan, S.; et al. Paneth cell-derived growth factors support tumorigenesis in the small intestine. Life Sci. Alliance. 2020, 43, e202000934. [Google Scholar] [CrossRef] [PubMed]
- Creff, J.; Malaquin, L.; Besson, A. In vitro models of intestinal epithelium: Toward bioengineered systems. J. Tissue Eng. 2021, 12, 2041731420985202. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.; Salzman, N. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Hendel, S.K.; Kellermann, L.; Hausmann, A.; Bindslev, N.; Jensen, K.B.; Nielsen, O.H. Tuft cells and their role in intestinal diseases. Front. Immunol. 2022, 13, 822867. [Google Scholar] [CrossRef]
- Steinway, S.N.; Saleh, J.; Koo, B.K.; Delacour, D.; Kim, D.H. Human microphysiological models of intestinal tissue and gut microbiome. Front. Bioeng. Biotechnol. 2020, 8, 725. [Google Scholar] [CrossRef]
- Siwczak, F.; Loffet, E.; Kaminska, M.; Koceva, H.; Mahe, M.M.; Mosig, A.S. Intestinal stem cell-on-chip to study human host-microbiota interaction. Front. Immunol. 2021, 12, 798552. [Google Scholar] [CrossRef]
- Tormási, J.; Abrankó, L. Impact of grape seed powder and black tea brew on lipid digestion-An in vitro co-digestion study with real foods. Nutrients 2023, 1510, 2395. [Google Scholar] [CrossRef]
- Tümer, E.; Bröer, A.; Balkrishna, S.; Jülich, T.; Bröer, S. Enterocyte-specific regulation of the apical nutrient transporter SLC6A19 B0AT1 by transcriptional and epigenetic networks. J. Biol. Chem. 2013, 28847, 33813–33823. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Martinez, M.N.; Dao, K.; Gabriel, V.; Zdyrski, C.; Jergens, A.E.; Atherly, T.; Iennarella-Servantez, C.A.; Burns, L.E.; Schrunk, D.; et al. Canine intestinal organoids as a novel in vitro model of intestinal drug permeability: A proof-of-concept study. Cells 2023, 129, 1269. [Google Scholar] [CrossRef]
- Xu, Y.; Shrestha, N.; Préat, V.; Beloqui, A. An overview of in vitro, ex vivo and in vivo models for studying the transport of drugs across intestinal barriers. Adv. Drug Deliv. Rev. 2021, 175, 113795. [Google Scholar] [CrossRef] [PubMed]
- Ashammakhi, N.; Nasiri, R.; Barros, N.R.; Tebon, P.; Thakor, J.; Goudie, M.; Shamloo, A.; Martin, M.G.; Khademhosseini, A. Gut-on-a-chip: Current progress and future opportunities. Biomaterials 2020, 255, 120196. [Google Scholar]
- Shim, K.Y.; Lee, D.; Han, J.; Nguyen, N.T.; Park, S.; Sung, J.H. Microfluidic gut-on-a-chip with three-dimensional villi structure. Biomed. Microdevices 2017, 192, 37. [Google Scholar] [CrossRef] [PubMed]
- Gościniak, A.; Eder, P.; Walkowiak, J.; Cielecka-Piontek, J. Artificial gastrointestinal models for nutraceuticals research-achievements and challenges: A practical review. Nutrients 2022, 1413, 2560. [Google Scholar] [CrossRef]
- Biagini, F.; Calvigioni, M.; Lapomarda, A.; Vecchione, A.; Magliaro, C.; De Maria, C.; Montemurro, F.; Celandroni, F.; Mazzantini, D.; Mattioli-Belmonte, M.; et al. A novel 3D in vitro model of the human gut microbiota. Sci. Rep. 2020, 10, 21499. [Google Scholar] [CrossRef]
- Hentschel, V.; Seufferlein, T.; Armacki, M. Intestinal organoids in coculture: Redefining the boundaries of gut mucosa ex vivo modeling. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 3216, G693–G704. [Google Scholar] [CrossRef]
- O’Farrell, C.; Stamatopoulos, K.; Simmons, M.; Batchelor, H. In vitro models to evaluate ingestible devices: Present status and current trends. Adv. Drug Deliv. Rev. 2021, 178, 113924. [Google Scholar] [CrossRef]
- Costa, J.; Ahluwalia, A. Advances and current challenges in intestinal in vitro model engineering: A Digest. Front. Bioeng. Biotechnol. 2019, 7, 144. [Google Scholar] [CrossRef]
- Kang, T.H.; Kim, H.J. Farewell to animal testing: Innovations on human intestinal microphysiological systems. Micromachines 2016, 77, 107. [Google Scholar] [CrossRef]
- Li, C.; Yu, W.; Wu, P.; Chen, X.D. Current in vitro digestion systems for understanding food digestion in human upper gastrointestinal tract. Trends Food Sci. Technol. 2020, 96, 114–126. [Google Scholar] [CrossRef]
- Sensoy, I. A review on the food digestion in the digestive tract and the used in vitro models. Curr. Res. Food Sci. 2021, 4, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Park, H.J.; Xiu, H.; Akoh, C.C.; Kong, F. Predicting intestinal effective permeability of different transport mechanisms: Comparing ex vivo porcine and in vitro dialysis models. J. Food Eng. 2023, 338, 111256. [Google Scholar] [CrossRef]
- González-Arias, C.A.; Marín, S.; Sanchis, V.; Ramos, A.J. Mycotoxin bioaccessibility/absorption assessment using in vitro digestion models: A review. World Mycotoxin J. 2013, 62, 167–184. [Google Scholar] [CrossRef]
- Lefebvre, D.E.; Venema, K.; Gombau, L.; Valerio, L.G., Jr.; Raju, J.; Bondy, G.S.; Bouwmeester, H.; Singh, R.P.; Clippinger, A.J.; Collnot, E.M.; et al. Utility of models of the gastrointestinal tract for assessment of the digestion and absorption of engineered nanomaterials released from food matrices. Nanotoxicology 2015, 94, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.; Preiser, H.; Maestracci, D.; Ghosh, B.K.; Cerda, J.J.; Crane, R.K. Purification of the human intestinal brush border membrane. Biochim. Biophys. Acta BBA-Biomembr. 1973, 3231, 98–112. [Google Scholar] [CrossRef]
- Kessler, M.; Acuto, O.; Storelli, C.; Murer, H.; Müller, M.; Semenza, G. A modified procedure for the rapid preparation of efficiently transporting vesicles from small intestinal brush border membranes. Their use in investigating some properties of D-glucose and choline transport systems. Biochim. Biophys. Acta BBA 1978, 5061, 136–154. [Google Scholar] [CrossRef]
- Patel, G.; Misra, A. 10—Oral Delivery of Proteins and Peptides: Concepts and Applications. In Challenges in Delivery of Therapeutic Genomics and Proteomics, 1st ed.; Misra, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 481–529. [Google Scholar]
- Said, H.M.; Redha, R.; Nylander, W. Biotin transport in basolateral membrane vesicles of human intestine. Gastroenterology 1988, 945, 1157–1163. [Google Scholar] [CrossRef]
- Carter, M.; Essner, R.; Goldstein, N.; Iyer, M. Guide to Research Techniques in Neuroscience, 3rd ed.; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Anjum, M.; Laitila, A.; Ouwehand, A.C.; Forssten, S.D. Current perspectives on gastrointestinal models to assess probiotic-pathogen interactions. Front. Microbiol. 2022, 13, 831455. [Google Scholar] [CrossRef]
- Kasendra, M.; Tovaglieri, A.; Sontheimer-Phelps, A.; Jalili-Firoozinezhad, S.; Bein, A.; Chalkiadaki, A.; Scholl, W.; Zhang, C.; Rickner, H.; Richmond, C.A.; et al. Development of a primary human Small Intestine-on-a-Chip using biopsy-derived organoids. Sci. Rep. 2018, 81, 2871. [Google Scholar] [CrossRef]
- Morelli, M.; Kurek, D.; Ng, C.P.; Queiroz, K. Gut-on-a-chip models: Current and future perspectives for host-microbial interactions research. Biomedicines 2023, 112, 619. [Google Scholar] [CrossRef]
- Corning Incorporated. Transwell®, SnapwellTM, Netwell, and Falcon® Permeable Supports. Available online: https://www.corning.com/au/en/products/life-sciences/products/permeable-supports/transwell-snapwell-netwell-falcon-permeable-supports.html (accessed on 12 March 2024).
- Bein, A.; Shin, W.; Jalili-Firoozinezhad, S.; Park, M.H.; Sontheimer-Phelps, A.; Tovaglieri, A.; Chalkiadaki, A.; Kim, H.J.; Ingber, D.E. Microfluidic organ-on-a-chip models of human intestine. Cell. Mol. Gastroenterol. Hepatol. 2018, 54, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Roupar, D.; Berni, P.; Martins, J.T.; Caetano, A.C.; Teixeira, J.A.; Clarisse Nobre, C. Bioengineering approaches to simulate human colon microbiome ecosystem. Trends Food Sci. Technol. 2021, 112, 808–822. [Google Scholar] [CrossRef]
- Rahman, S.; Ghiboub, M.; Donkers, J.M.; van de Steeg, E.; van Tol, E.A.F.; Hakvoort, T.B.M.; de Jonge, W.J. The Progress of intestinal epithelial models from cell lines to gut-on-chip. Int. J. Mol. Sci. 2021, 2224, 13472. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Failla, M.L. Intestinal cell models for investigating the uptake, metabolism and absorption of dietary nutrients and bioactive compounds. Curr. Opin. Food Sci. 2021, 41, 169–179. [Google Scholar] [CrossRef]
- Sontheimer-Phelps, A.; Chou, D.B.; Tovaglieri, A.; Ferrante, T.C.; Duckworth, T.; Fadel, C.; Frismantas, V.; Sutherland, A.D.; Jalili-Firoozinezhad, S.; Kasendra, M.; et al. Human colonon-a-chip enables continuous in vitro analysis of colon mucus layer accumulation and physiology. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Langerholc, T.; Maragkoudakis, P.A.; Wollgast, J.; Gradisnik, L.; Cencic, A. Novel and established intestinal cell line models—An indispensable tool in food science and nutrition. Trends Food Sci. Technol. 2011, 22, S11–S20. [Google Scholar] [CrossRef]
- Cencič, A.; Langerholc, T. Functional cell models of the gut and their applications in food microbiology—A review. Int. J. Food Microbiol. 2010, 141, S4–S14. [Google Scholar] [CrossRef]
- Jensen, H.H.; Holst, M.R.; Login, F.H.; Morgen, J.J.; Nejsum, L.N. Ectopic expression of aquaporin-5 in noncancerous epithelial MDCK cells changes cellular morphology and actin fiber formation without inducing epithelial-to-mesenchymal transition. Am. J. Physiol. Cell Physiol. 2018, 3146, C654–C661. [Google Scholar] [CrossRef] [PubMed]
- Bédard, P.; Gauvin, S.; Ferland, K.; Caneparo, C.; Pellerin, È.; Chabaud, S.; Bolduc, S. Innovative human three-dimensional tissue-engineered models as an alternative to animal testing. Bioengineering 2020, 73, 115. [Google Scholar] [CrossRef]
- Pimenta, J.; Ribeiro, R.; Almeida, R.; Costa, P.F.; da Silva, M.A.; Pereira, B. Organ-on-chip approaches for intestinal 3D in vitro modeling. Cell. Mol. Gastroenterol. Hepatol. 2022, 132, 351–367. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, X.; Dowbaj, A.M.; Sljukic, A.; Bratlie, K.; Lin, L.; Fong, E.L.S.; Balachander, G.M.; Chen, Z.; Soragni, A.; et al. Organoids. Nat. Rev. Methods Prim. 2022, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Li, H.; Collins, J.J.; Ingber, D.E. Contributions of microbiome and mechanical deformation to intestinal bacterial overgrowth and inflammation in a human gut-on-a-chip. Proc. Natl. Acad. Sci. USA 2016, 113, E7–E15. [Google Scholar] [CrossRef] [PubMed]
- Donkers, J.M.; Amirabadi, H.E.; van de Steeg, E. Intestine-on-a-chip: Next level in vitro research model of the human intestine. Curr. Opin. Toxicol. 2021, 25, 6–14. [Google Scholar] [CrossRef]
- Thomas, D.P.; Zhang, J.; Nguyen, N.T.; Ta, H.T. Microfluidic gut-on-a-chip: Fundamentals and challenges. Biosensors 2023, 131, 136. [Google Scholar] [CrossRef]
- Zhang, D.; Qiao, L. Intestine-on-a-chip for intestinal disease study and pharmacological research. View 2023, 4, 20220037. [Google Scholar] [CrossRef]
- Barrila, J.; Radtke, A.L.; Crabbé, A.; Sarker, S.F.; Herbst-Kralovetz, M.M.; Ott, C.M.; Nickerson, C.A. Organotypic 3D cell culture models: Using the rotating wall vessel to study host-pathogen interactions. Nat. Rev. Microbiol. 2010, 8, 791–801. [Google Scholar] [CrossRef]
- Costello, C.M.; Phillipsen, M.B.; Hartmanis, L.M.; Kwasnica, M.A.; Chen, V.; Hackam, D.; Chang, M.W.; Bentley, W.E.; March, J.C. Microscale bioreactors for in situ characterization of GI epithelial cell physiology. Sci. Rep. 2017, 7, 12515. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Chen, Y.G. Generation of 3D human gastrointestinal organoids: Principle and applications. Cell Regen. 2020, 91, 6. [Google Scholar] [CrossRef]
- Stanton, J.E.; Grabrucker, A.M. The use of organoids in food research. Curr. Opin. Food Sci. 2023, 49, 100977. [Google Scholar] [CrossRef]
- Poletti, M.; Arnauts, K.; Ferrante, M.; Korcsmaros, T. Organoid-based models to study the role of host-microbiota interactions in IBD. J. Crohn’s Colitis 2021, 157, 1222–1235. [Google Scholar] [CrossRef]
- Bossink, E.G.B.M.; Segerink, L.I.; Odijk, M. Organ-on-chip technology for aerobic intestinal host—anaerobic microbiota research. Organs–A-Chip 2022, 4, 100013. [Google Scholar] [CrossRef]
- Xavier, M.; Rodrigues, P.M.; Neto, M.D.; Guedes, M.I.; Calero, V.; Pastrana, L.; Gonçalves, C. From mouth to gut: Microfluidic in vitro simulation of human gastro-intestinal digestion and intestinal permeability. Analyst 2023, 14814, 3193–3203. [Google Scholar] [CrossRef] [PubMed]
- Imura, Y.; Asano, Y.; Sato, K.; Yoshimura, E. A microfluidic system to evaluate intestinal absorption. Anal. Sci. 2009, 25, 1403–1407. [Google Scholar] [CrossRef]
- Workman, M.J.; Gleeson, J.P.; Troisi, E.J.; Estrada, H.Q.; Kerns, S.J.; Hinojosa, C.D.; Hamilton, G.A.; Targan, S.R.; Svendsen, C.N.; Barrett, R.J. Enhanced utilization of induced pluripotent stem cell-derived human intestinal organoids using microengineered chips. Cell. Mol. Gastroenterol. Hepatol. 2017, 54, 669–677.e2. [Google Scholar] [CrossRef] [PubMed]
- Valiei, A.; Aminian-Dehkordi, J.; Mofrad, M.R.K. Gut-on-a-chip models for dissecting the gut microbiology and physiology. APL Bioeng. 2023, 71, 011502. [Google Scholar] [CrossRef]
- Marrero, D.; Pujol-Vila, F.; Vera, D.; Gabriel, G.; Illa, X.; Elizalde-Torrent, A.; Alvarez, M.; Villa, R. Gut-on-a-chip: Mimicking and monitoring the human intestine. Biosens. Bioelectron. 2021, 181, 113156. [Google Scholar] [CrossRef]
- Greenhalgh, K.; Ramiro-Garcia, J.; Heinken, A.; Ullmann, P.; Bintener, T.; Pacheco, M.P.; Baginska, J.; Shah, P.; Frachet, A.; Halder, R.; et al. Integrated in vitro and in silico modeling delineates the molecular effects of a synbiotic regimen on colorectal-cancer-derived cells. Cell Rep. 2019, 275, 1621–1632.e9. [Google Scholar] [CrossRef] [PubMed]
- Marzorati, M.; Vanhoecke, B.; De Ryck, T.; Sadaghian Sadabad, M.; Pinheiro, I.; Possemiers, S.; Van den Abbeele, P.; Derycke, L.; Bracke, M.; Pieters, J.; et al. The HMI™ module: A new tool to study the Host-Microbiota Interaction in the human gastrointestinal tract in vitro. BMC Microbiol. 2014, 14, 133. [Google Scholar] [CrossRef]
- Jalili-Firoozinezhad, S.; Gazzaniga, F.S.; Calamari, E.L.; Camacho, D.M.; Fadel, C.W.; Bein, A.; Swenor, B.; Nestor, B.; Cronce, M.J.; Tovaglieri, A.; et al. A complex human gut microbiome cultured in an anaerobic intestine-on-a-chip. Nat. Biomed. Eng. 2019, 3, 520–531. [Google Scholar] [CrossRef]
- Shin, W.; Wu, A.; Massidda, M.W.; Foster, C.; Thomas, N.; Lee, D.W.; Koh, H.; Ju, Y.; Kim, J.; Kim, H.J. A robust longitudinal co-culture of obligate anaerobic gut microbiome with human intestinal epithelium in an anoxic-oxic interface-on-a-chip. Front. Bioeng. Biotechnol. 2019, 7, 13. [Google Scholar] [CrossRef]
- de Haan, P.; Santbergen, M.J.C.; van der Zande, M.; Bouwmeester, H.; Nielen, M.W.F.; Verpoorte, E. A versatile, compartmentalised gut-on-a-chip system for pharmacological and toxicological analyses. Sci. Rep. 2021, 111, 4920. [Google Scholar] [CrossRef] [PubMed]
- Kalungwana, N.; Marshall, L.; Mackie, A.; Boesch, C. An ex vivo intestinal absorption model is more effective than an in vitro cell model to characterise absorption of dietary carotenoids following simulated gastrointestinal digestion. Food Res. Int. 2023, 166, 112558. [Google Scholar] [CrossRef] [PubMed]
- Gijzen, L.; Marescotti, D.; Raineri, E.; Nicolas, A.; Lanz, H.L.; Guerrera, D.; van Vught, R.; Joore, J.; Vulto, P.; Peitsch, M.C.; et al. An intestine-on-a-chip model of plug-and-play modularity to study inflammatory processes. SLAS Technol. 2020, 256, 585–597. [Google Scholar] [CrossRef] [PubMed]
- von Erlach, T.; Saxton, S.; Shi, Y.; Minahan, D.; Reker, D.; Javid, F.; Lee, Y.L.; Schoellhammer, C.; Esfandiary, T.; Cleveland, C.; et al. Robotically handled whole-tissue culture system for the screening of oral drug formulations. Nat. Biomed. Eng. 2020, 45, 544–559. [Google Scholar] [CrossRef]
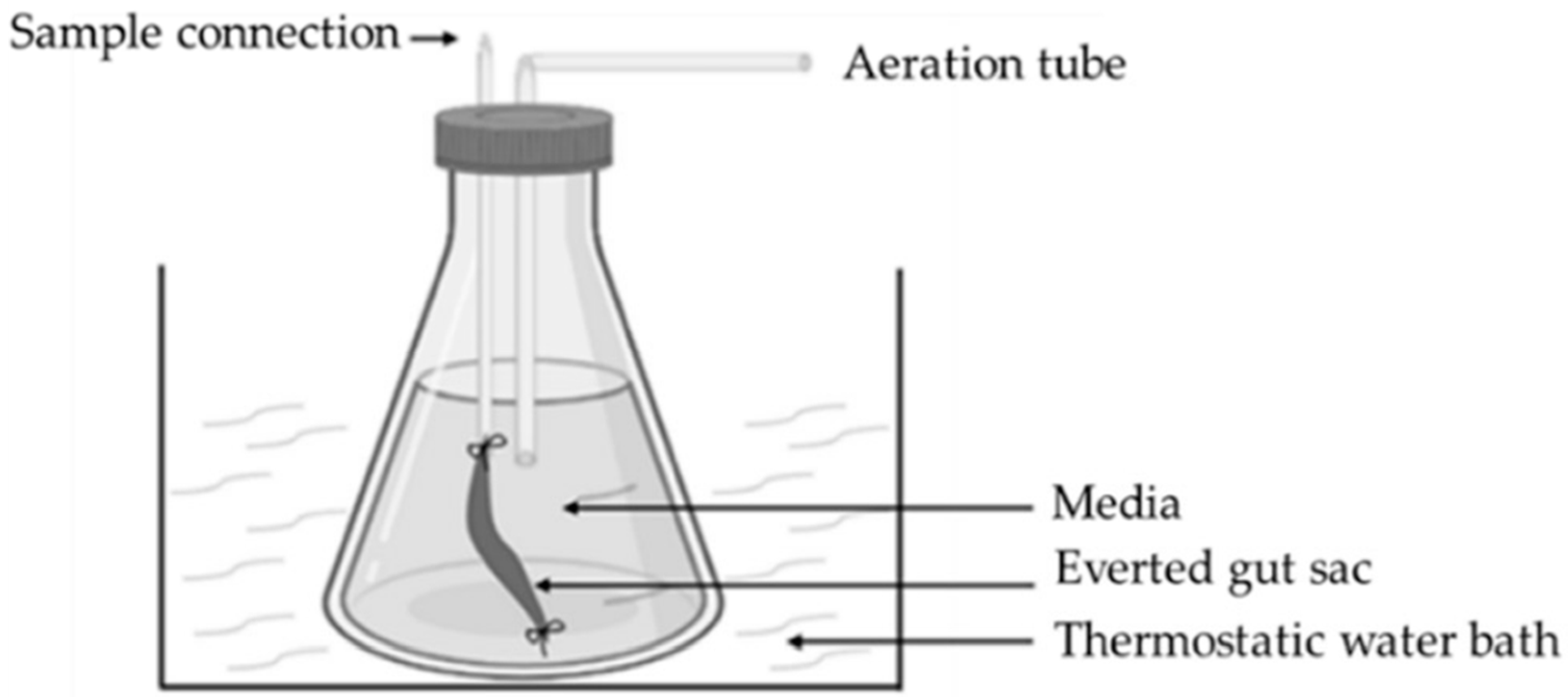
- Hamilton, K.L.; Butt, A.G. Glucose transport into everted sacs of the small intestine of mice. Adv. Physiol. Educ. 2013, 374, 415–426. [Google Scholar] [CrossRef]
- van Krimpen, M.M.; Hulst, M.M.; van der Meulen, J.; Schokker, D.; Savelkoul, H.F.J.; Tijhaar, E.J.; Rutten, V.P.M.G. Nutritional Intervention in Animals: Benchmarking of Strategies, Monitoring Biomarkers and Immune Competence. Available online: https://edepot.wur.nl/320444 (accessed on 27 December 2022).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Advantage | Disadvantage |
|---|---|---|
| Centrifugation | Easy to use; cheap; reproducible; high-throughput screening. | It simplifies the in vivo conditions too much; active transport is not simulated |
| Filtration | ||
| Dialysis | ||
| Biomimetic membranes | Easy to use; cheap; reproducible; high-throughput screening [22] | Only passive transcellular transport is measured [22] |
| Brush border membrane vesicles | All components of the cellular membrane are present (transporters, enzymes) | It contains only one side of the intestinal epithelial cell (apical or basolateral part); the isolation of completely pure vesicles is not possible; transporters, enyzmes associated with the membrane can be damaged during isolation; small sample volume for analytical analysis [22] |
| Basolateral membrane vesicles | ||
| Primary cells | Phenotype similar to in vivo conditions | Laborious; low reproducibility |
| Cell lines | Reproducible | Usually, they have the phenotype of cancerous cells, which is different than the in vivo status; In cases of monocultures (e.g., Caco-2 cells), it may lack the mucus layer that influences absorption |
| Organoids | Imitates the 3D organization of intestinal epithelial cells | Laborious; the apical side is not accessible (with the exception of fragmented organoids and organoids with reversed polarity) |
| Transwell systems® | Standard method for studying transport processes | Shear stress and intestinal motility is not reproduced |
| Utilization of scaffolds | Imitates the 3D organization of intestinal epithelial cells | Time-consuming; the inter- and intra-laboratory variability is higher [22]; the reproducibility is further diminished if scaffolds of animal origin are utilized |
| Microfluidic systems | Imitates the 3D organization of intestinal epithelial cells; shear stress and/or intestinal motility is reproduced; several microfluidic systems also reproduce the microbiota, which can be important in case of numerous research questions | Expensive [22]; laborious; complex laboratory equipment is necessary |
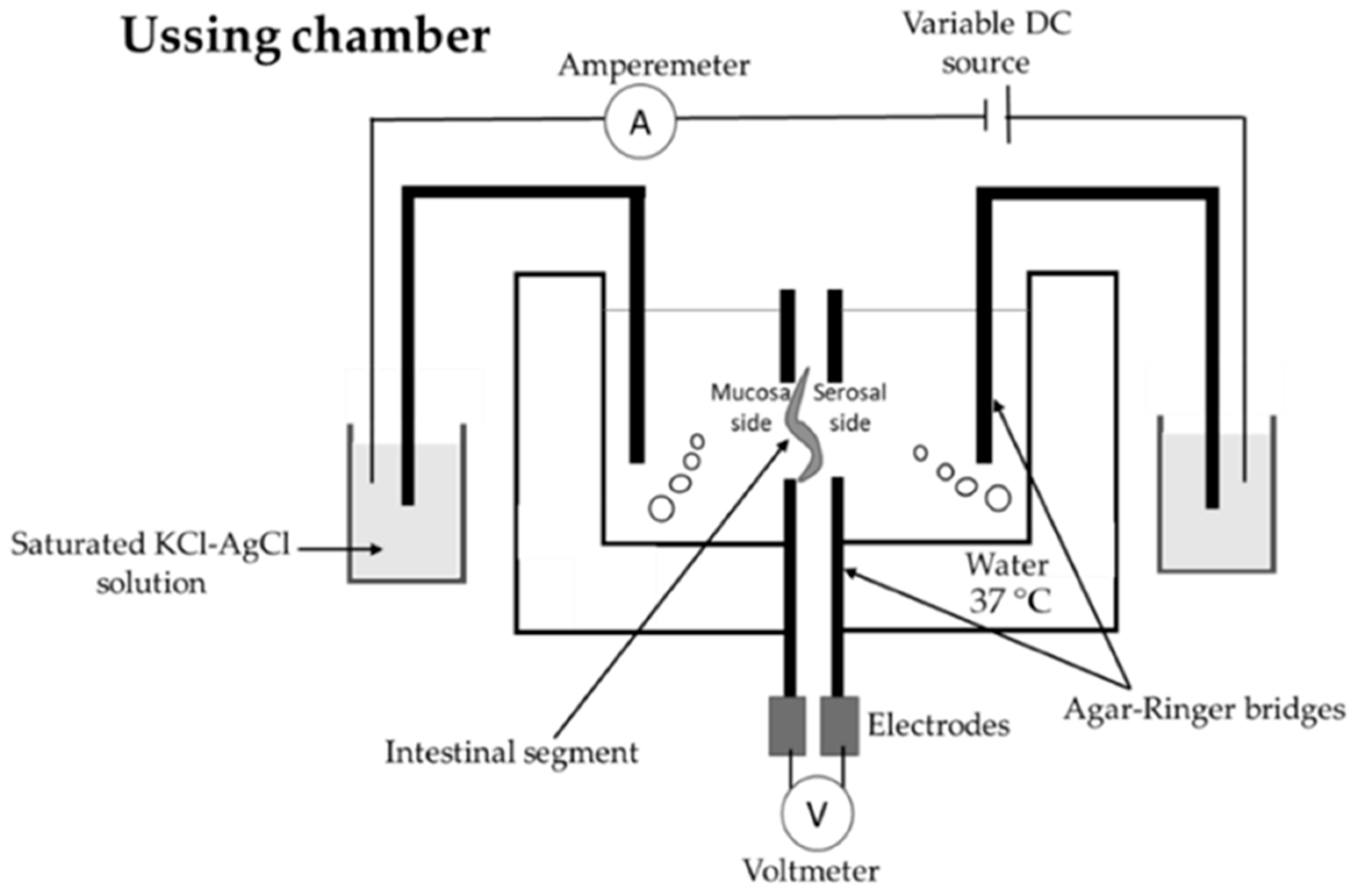
| Ussing chamber | The tissue viability can be continuously monitored; better stirring conditions than in cases of everted intestinal sac or Intestine® system from TNO [22] | The integrity of the tissue is maintained for 2–3 h; the equipment is expensive [22] |
| Franz cell | Better stirring conditions than in cases of everted intestinal sac or Intestine® system from TNO [22] | The integrity of the tissue is maintained for 2 h; special equipment is needed [22] |
| Everted intestinal sac | Cheap equipment | Tissue viability is less than 30 min; eversion can cause structural damage in the tissue; the sample is not properly stirred [22] |
| Gastrointestinal tissue robotic interface system | Histological integrity maintained for one week [78] | Complex, specialized equipment is necessary |
| Intestine® system from TNO | Simple; commercially available [22] | The sample is not properly stirred [22] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antal, O.; Dalmadi, I.; Takács, K. Upgrading In Vitro Digestion Protocols with Absorption Models. Appl. Sci. 2024, 14, 8320. https://doi.org/10.3390/app14188320
Antal O, Dalmadi I, Takács K. Upgrading In Vitro Digestion Protocols with Absorption Models. Applied Sciences. 2024; 14(18):8320. https://doi.org/10.3390/app14188320
Chicago/Turabian StyleAntal, Otilia, István Dalmadi, and Krisztina Takács. 2024. "Upgrading In Vitro Digestion Protocols with Absorption Models" Applied Sciences 14, no. 18: 8320. https://doi.org/10.3390/app14188320