

Exploring the Potential Protective Effect of Probiotics in Obesity-Induced Colorectal Cancer: What Insights Can In Vitro Models Provide?

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
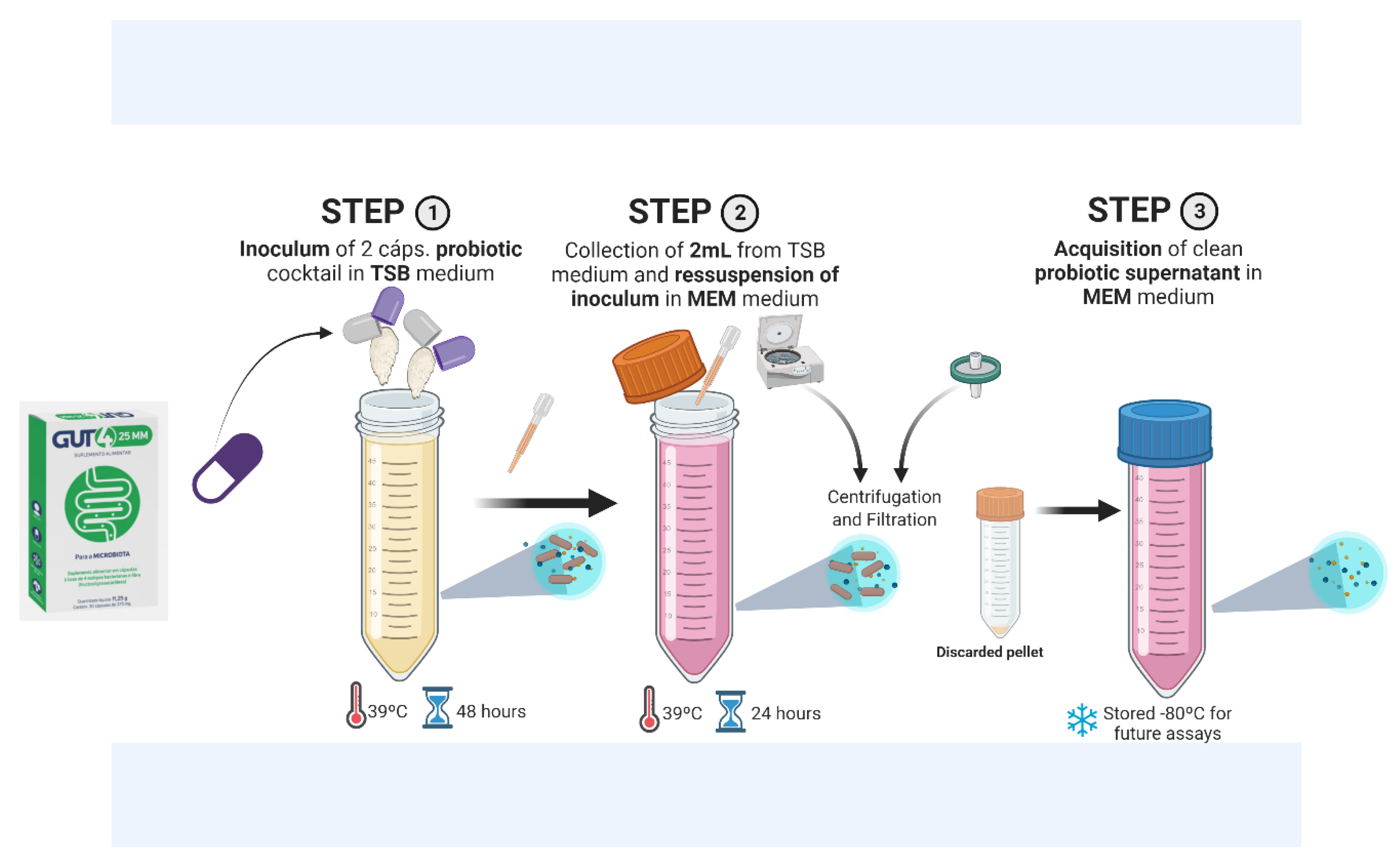
2.1. Growth of Probiotic Bacteria and Collection of the Supernatant
2.2. Adipocyte Differentiation and Secretome Collect
2.3. Viability Assay in Caco2
2.4. Injury Assay in Caco2
2.5. Antioxidant Assay in Caco2
2.6. Inflammatory Genes Quantified by RT-qPCR
2.7. Statistical Analyses
3. Results
3.1. Gene Expression
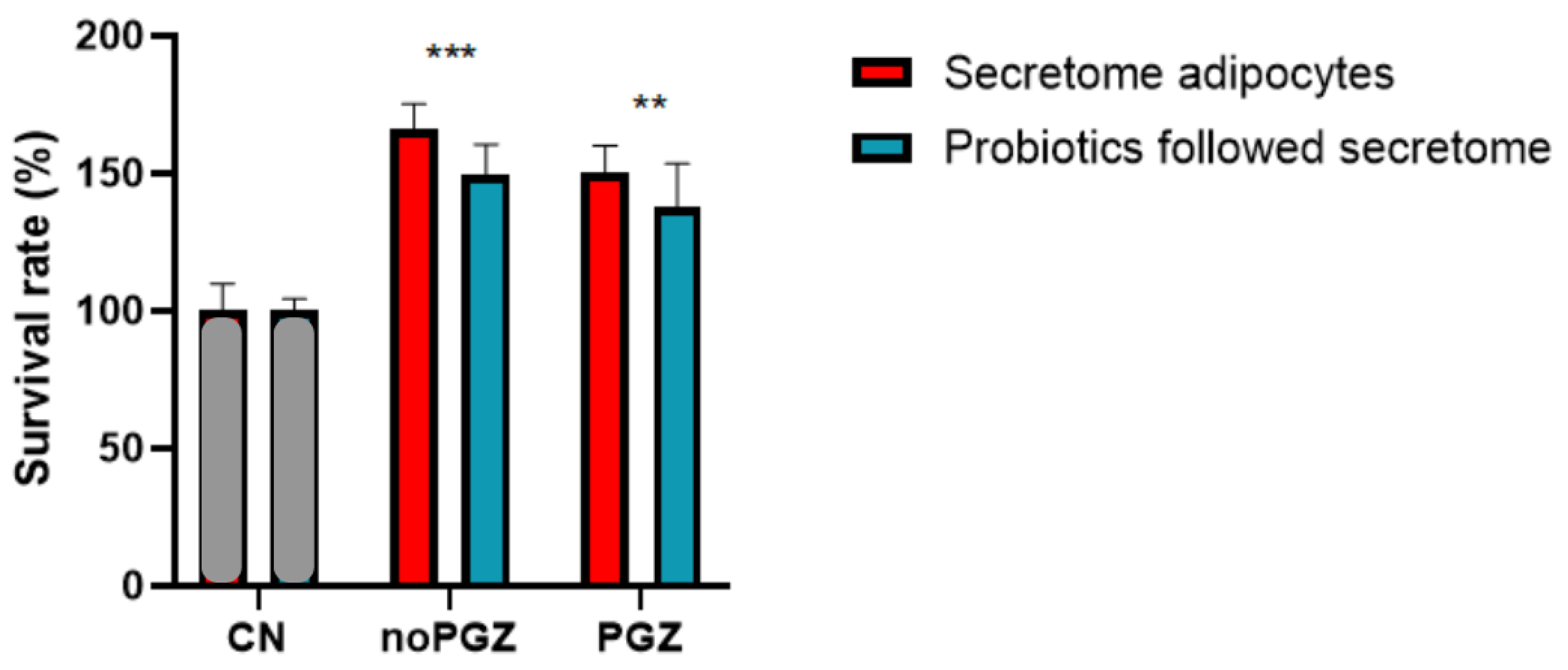
3.2. Adiposity Increases Colon Tumor
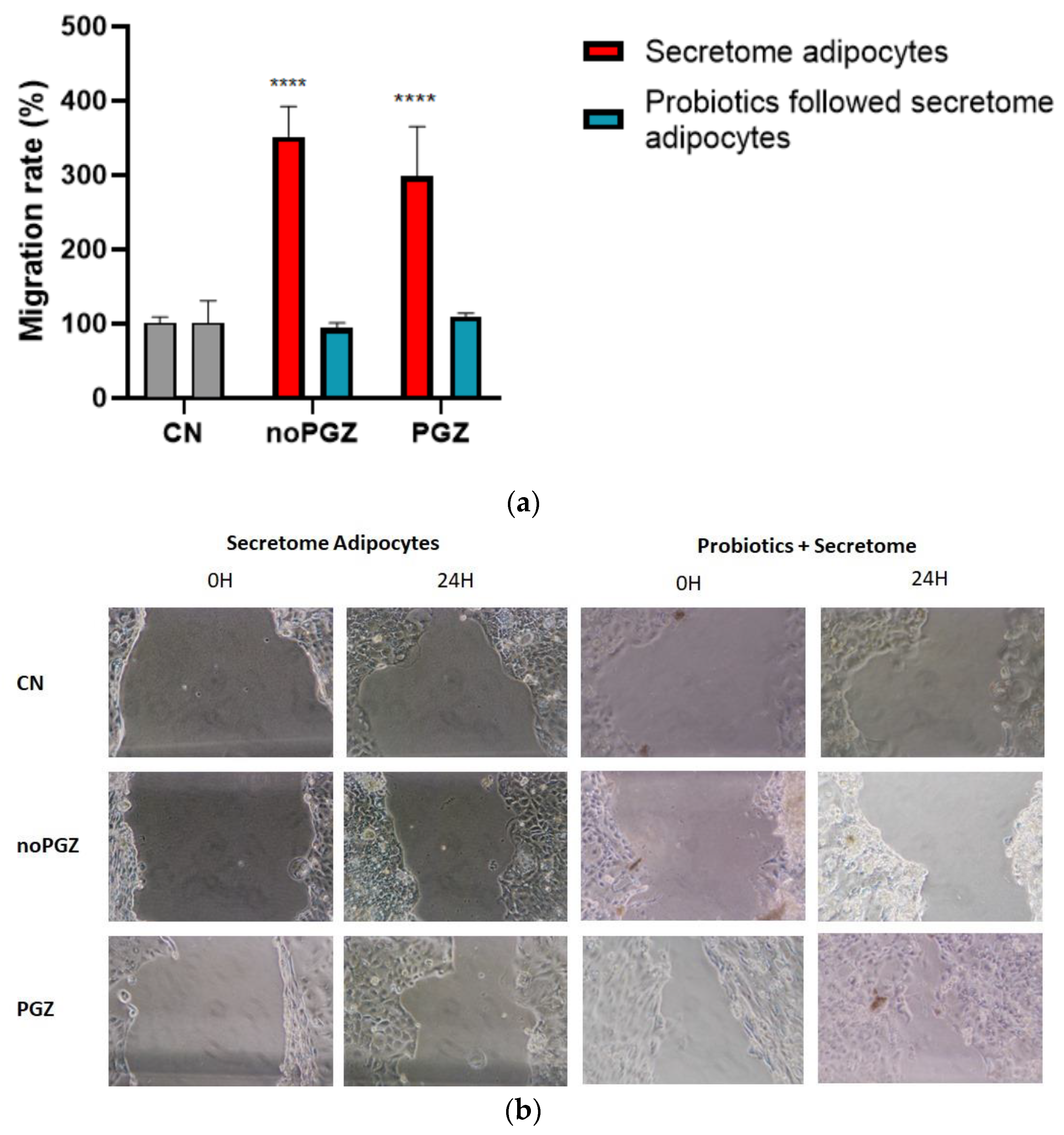
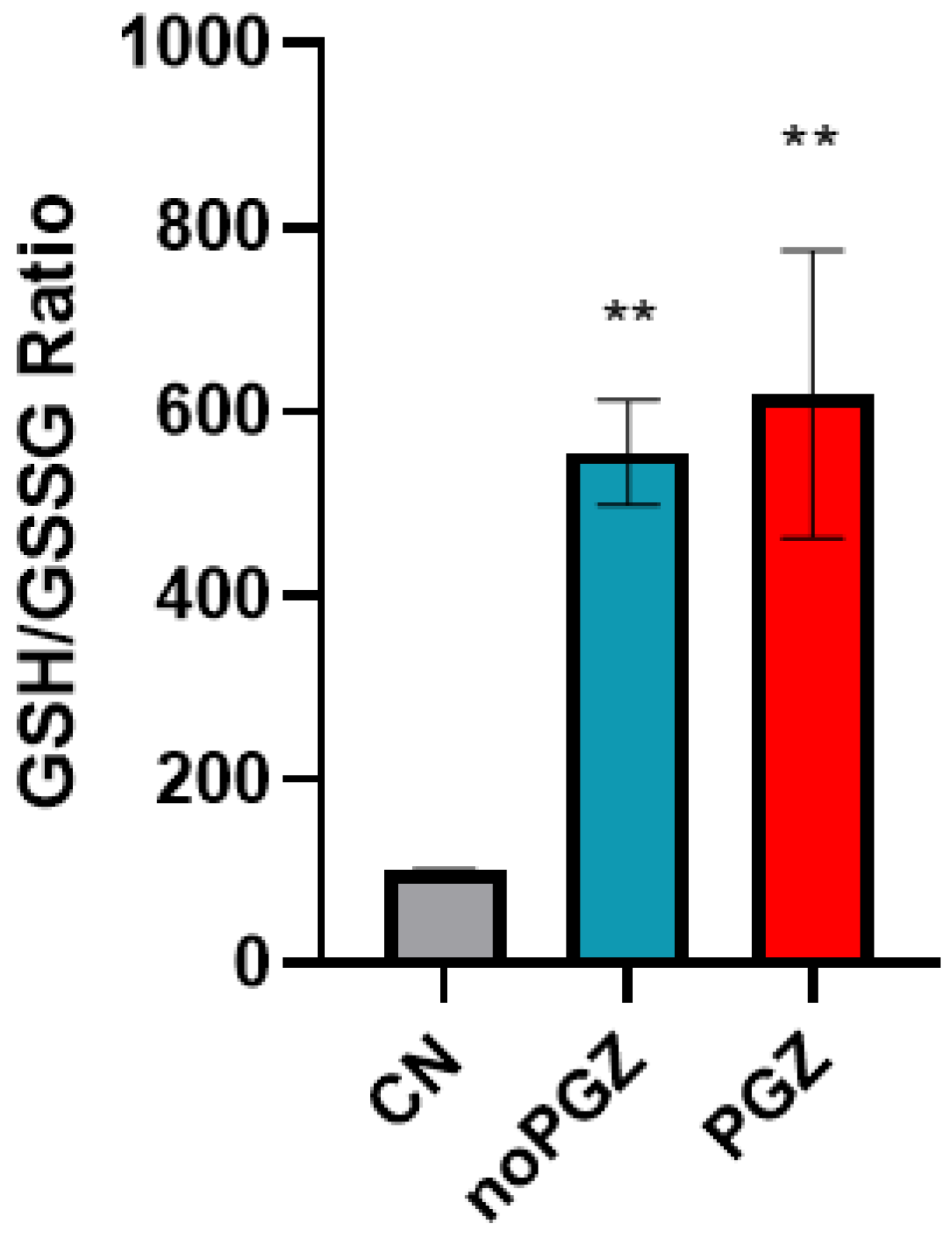
3.3. Invasion Activity and Antioxidant
3.4. Inflammatory Genes Responsive to Adiposity
4. Discussion
5. Conclusions
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 12 July 2023).
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Regional Office for Europe. WHO European Regional Obesity Report. 2022. Available online: https://iris.who.int/handle/10665/353747 (accessed on 3 October 2023).
- Kamal, N.; Ilowefah, M.A.; Hilles, A.R.; Anua, N.A.; Awin, T.; Alshwyeh, H.A.; Aldosary, S.K.; Jambocus, N.G.S.; Alosaimi, A.A.; Rahman, A.; et al. Genesis and Mechanism of Some Cancer Types and an Overview on the Role of Diet and Nutrition in Cancer Prevention. Molecules 2022, 27, 1794. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Xi, Y.; Huang, Z.; Xu, P. Linking Obesity with Colorectal Cancer: Epidemiology and Mechanistic Insights. Cancers 2020, 12, 1408. [Google Scholar] [CrossRef] [PubMed]
- Nieman, K.M.; Kenny, H.A.; Penicka, C.V.; Ladanyi, A.; Buell-Gutbrod, R.; Zillhardt, M.R.; Romero, I.L.; Carey, M.S.; Mills, G.B.; Hotamisligil, G.S.; et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat. Med. 2011, 17, 1498–1503. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, P.; Sarma, D.K.; Kumawat, M.; Tiwari, R.; Verma, V.; Nagpal, R.; Kumar, M. Implication of Obesity and Gut Microbiome Dysbiosis in the Etiology of Colorectal Cancer. Cancers 2023, 15, 1913. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Machado, S.A.; Pasquarelli-do-Nascimento, G.; da Silva, D.; Farias, G.R.; de Oliveira Santos, I.; Baptista, L.B. Browning of the white adipose tissue regulation: New insights into nutritional and metabolic relevance in health and diseases. Nutr. Metab. 2022, 19, 61. [Google Scholar] [CrossRef]
- Endalifer, M.L.; Diress, G. Epidemiology, Predisposing Factors, Biomarkers, and Prevention Mechanism of Obesity: A Systematic Review. J. Obes. 2020, 2020, 6134362. [Google Scholar] [CrossRef]
- Palavicini, J.P.; Chavez-Velazquez, A.; Fourcaudot, M.; Tripathy, D.; Pan, M.; Norton, L.; DeFronzo, R.A.; Shannon, C.E. The Insulin-Sensitizer Pioglitazone Remodels Adipose Tissue Phospholipids in Humans. Front. Physiol. 2021, 12, 784391. [Google Scholar] [CrossRef]
- Cancello, R.; Tordjman, J.; Poitou, C.; Guilhem, G.; Bouillot, J.L.; Hugol, D.; Coussieu, C.; Basdevant, A.; Bar Hen, A.; Bedossa, P.; et al. Increased infiltration of macrophages in omental adipose tissue is associated with marked hepatic lesions in morbid human obesity. Diabetes 2006, 55, 1554–1561. [Google Scholar] [CrossRef]
- Divella, R.; De Luca, R.; Abbate, I.; Naglieri, E.; Daniele, A. Obesity and cancer: The role of adipose tissue and adipo-cytokines-induced chronic inflammation. J. Cancer 2016, 7, 2346–2359. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef]
- Cӑtoi, A.F.; Vodnar, D.C.; Corina, A.; Nikolic, D.; Citarrella, R.; Pérez-Martínez, P.; Rizzo, M. Gut Microbiota, Obesity and Bariatric Surgery: Current Knowledge and Future Perspectives. Curr. Pharm. Des. 2019, 25, 2038–2050. [Google Scholar] [CrossRef]
- Teixeira, C.; Sousa, A.P.; Santos, I.; Rocha, A.C.; Alencastre, I.; Pereira, A.C.; Martins-Mendes, D.; Barata, P.; Baylina, P.; Fernandes, R. Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ). Biology 2022, 11, 806. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.; Almeida, J.; Prudêncio, C.; Fernandes, R.; Soares, R. Effect of Adipocyte Secretome in Melanoma Progression and Vasculogenic Mimicry. J. Cell Biochem. 2016, 117, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, R.S.; Pinto, J.R.F.E.; Santos, L.V.; Góis, M.C.; Silva, R.P.; Silva, H.M. Rectal cancer survival in a Brazilian Cancer Reference Unit. J. Coloproctol. 2015, 36, 203–207. [Google Scholar] [CrossRef]
- Sánchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordóñez, R.; Medina, J.A.; Gómez-Millán, J.; Queipo-Ortuño, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers 2020, 12, 1406. [Google Scholar] [CrossRef]
- Machado, F.F.; Lazzaretti, R.K.; Poziomyck, A.K. Uso de prebióticos, probióticos e simbióticos nos pré e pós- operatórios do câncer colorretal: Uma revisão. Rev. Bras. Cancerol. 2014, 60, 363–370. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. Effect of probiotics in the treatment of colorretal cancer. Arch. Health Investig. 2019, 8, 394–399. [Google Scholar] [CrossRef]
- Drago, L. Probiotics and Colon Cancer. Microorganisms 2019, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, M.S.; Santos, C.I.D.; Araújo, M.C.; Girón, J.A.; Fernandes, E.S.; Monteiro-Neto, V. Probiotics, mechanisms of action, and clinical perspectives for diarrhea management in children. Food Funct. 2018, 9, 5074–5095. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, A.; Rodriguez, R.M.; Segura-Sampedro, J.J.; Ochogavía-Seguí, A.; Romaguera, D.; Barceló-Coblijn, G. Insights behind the Relationship between Colorectal Cancer and Obesity: Is Visceral Adipose Tissue the Missing Link? Int. J. Mol. Sci. 2022, 23, 13128. [Google Scholar] [CrossRef] [PubMed]
- Safaei, M.; Sundararajan, E.A.; Driss, M.; Boulila, W.; Shapi’i, A. A systematic literature review on obesity: Understanding the causes & consequences of obesity and reviewing various machine learning approaches used to predict obesity. Comput. Biol. Med. 2021, 136, 104754. [Google Scholar] [CrossRef]
- Misra, S.; Pandey, P.; Mishra, H.N. Novel approaches for co-encapsulation of probiotic bacteria with bioactive compounds, their health benefits and functional food product development: A review. Trends Food Sci. Technol. 2021, 109, 340–351. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Koklesova, L.; Kubatka, P.; Büsselberg, D. Immunomodulation by Gut Microbiome on Gastrointestinal Cancers: Focusing on Colorectal Cancer. Cancers 2022, 14, 2140. [Google Scholar] [CrossRef] [PubMed]
- Tajik, N.; Frech, M.; Schulz, O.; Schälter, F.; Lucas, S.; Azizov, V.; Dürholz, K.; Steffen, F.; Omata, Y.; Rings, A.; et al. Targeting zonulin and intestinal epithelial barrier function to prevent onset of arthritis. Nat. Commun. 2020, 11, 1995. [Google Scholar] [CrossRef]
- Cheng, Y.; Ling, Z.; Li, L. The Intestinal Microbiota and Colorectal Cancer. Front. Immunol. 2020, 11, 615056. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Frantz, A.L.; Rogier, E.W.; Weber, C.R.; Shen, L.; Cohen, D.A.; Fenton, L.A.; Bruno, M.E.; Kaetzel, C.S. Targeted deletion of MyD88 in intestinal epithelial cells results in compromised antibacterial immunity associated with downregulation of polymeric immunoglobulin receptor, mucin-2, and antibacterial peptides. Mucosal Immunol. 2012, 5, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Rébé, C.; Ghiringhelli, F. Interleukin-1β and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef] [PubMed]
- Nagaraju, G.P.; Aliya, S.; Alese, O.B. Role of adiponectin in obesity related gastrointestinal carcinogenesis. Cytokine Growth Factor Rev. 2015, 26, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Hirschenson, J.; Moore, A.; Mailloux, R.J. Conditions Conducive to the Glutathionylation of Complex I Subunit NDUFS1 Augment ROS Production following the Oxidation of Ubiquinone Linked Substrates, Glycerol-3-Phosphate and Proline. Antioxidants 2022, 11, 2043. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jin, P.P.; Sun, X.C.; Hu, T.T. Thiazolidinediones and risk of colorectal cancer in patients with diabetes mellitus: A meta-analysis. Saudi J. Gastroenterol. 2018, 24, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, I.; Yamazaki, K.; Oyama, K.; Hayashi, H.; Tajima, H.; Kitagawa, H.; Fushida, S.; Fujimura, T.; Ohta, T. Pioglitazone inhibits the proliferation and metastasis of human pancreatic cancer cells. Oncol. Lett. 2014, 8, 2709–2714. [Google Scholar] [CrossRef]
- Belfiore, A.; Genua, M.; Malaguarnera, R. PPAR-γ agonists and their effects on IGF-I receptor signaling: Implications for cancer. PPAR Res. 2009, 2009, 830501. [Google Scholar] [CrossRef]
- Elnemr, A.; Ohta, T.; Iwata, K.; Ninomia, I.; Fushida, S.; Nishimura, G.; Kitagawa, H.; Kayahara, M.; Yamamoto, M.; Terada, T.; et al. PPARgamma ligand (thiazolidinedione) induces growth arrest and differentiation markers of human pancreatic cancer cells. Int. J. Oncol. 2000, 17, 1157–1164. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Species | Primer Forward (5′-3′) | Primer Reverse (3′-5′) | Size (bp—Base Pairs) | Annealing Temperature (°C) |
|---|---|---|---|---|
| Lactobacillus ssp. | AGCAGTAGGGAATCTTCCA | CATTYCACCGCTACACATG | 346 | 60 |
| Total Bacteria | CCTACGGGAGGCAGCAG | ATTACCGCGGCTGCTGG | 169 | 60 |
| Gene | Primer Forward (5′-3′) | Primer Reverse (3′-5′) |
|---|---|---|
| IL 1-β | ACCTAGCTGTCAACGTGTGG | TCAAAGCAATGTGCTGGTGC |
| IL-10 | TGAAAACAAGAGCAAGGCCG | ATAGAGTCGCCACCCTGATG |
| Number of Cycles | Temperature (°C) | Time |
|---|---|---|
| 1 | 95 | 02:00 |
| 40 | 95 | 00:05 |
| 60 | 00:20 | |
| 1 | 4 | ∞ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viana, R.; Rocha, A.C.; Sousa, A.P.; Ferreira, D.; Fernandes, R.; Almeida, C.; Pais, P.J.; Baylina, P.; Pereira, A.C. Exploring the Potential Protective Effect of Probiotics in Obesity-Induced Colorectal Cancer: What Insights Can In Vitro Models Provide? Appl. Sci. 2024, 14, 951. https://doi.org/10.3390/app14020951
Viana R, Rocha AC, Sousa AP, Ferreira D, Fernandes R, Almeida C, Pais PJ, Baylina P, Pereira AC. Exploring the Potential Protective Effect of Probiotics in Obesity-Induced Colorectal Cancer: What Insights Can In Vitro Models Provide? Applied Sciences. 2024; 14(2):951. https://doi.org/10.3390/app14020951
Chicago/Turabian StyleViana, Rejane, Ana C. Rocha, André P. Sousa, Diogo Ferreira, Rúben Fernandes, Cátia Almeida, Patrick J. Pais, Pilar Baylina, and Ana Cláudia Pereira. 2024. "Exploring the Potential Protective Effect of Probiotics in Obesity-Induced Colorectal Cancer: What Insights Can In Vitro Models Provide?" Applied Sciences 14, no. 2: 951. https://doi.org/10.3390/app14020951