
Optimization of One-Step In Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cultivation Conditions
2.3. One-Step In Situ Transesterification
2.4. Quantitative Analysis
2.5. Two-Step Method for Lipid Extraction and Transesterification
3. Results
3.1. Analysis of FAME Profiles and Contents
3.2. Optimization of In Situ Transesterification Reaction
3.2.1. Effect of Methanol-to-Biomass Ratio on Reaction Efficiency
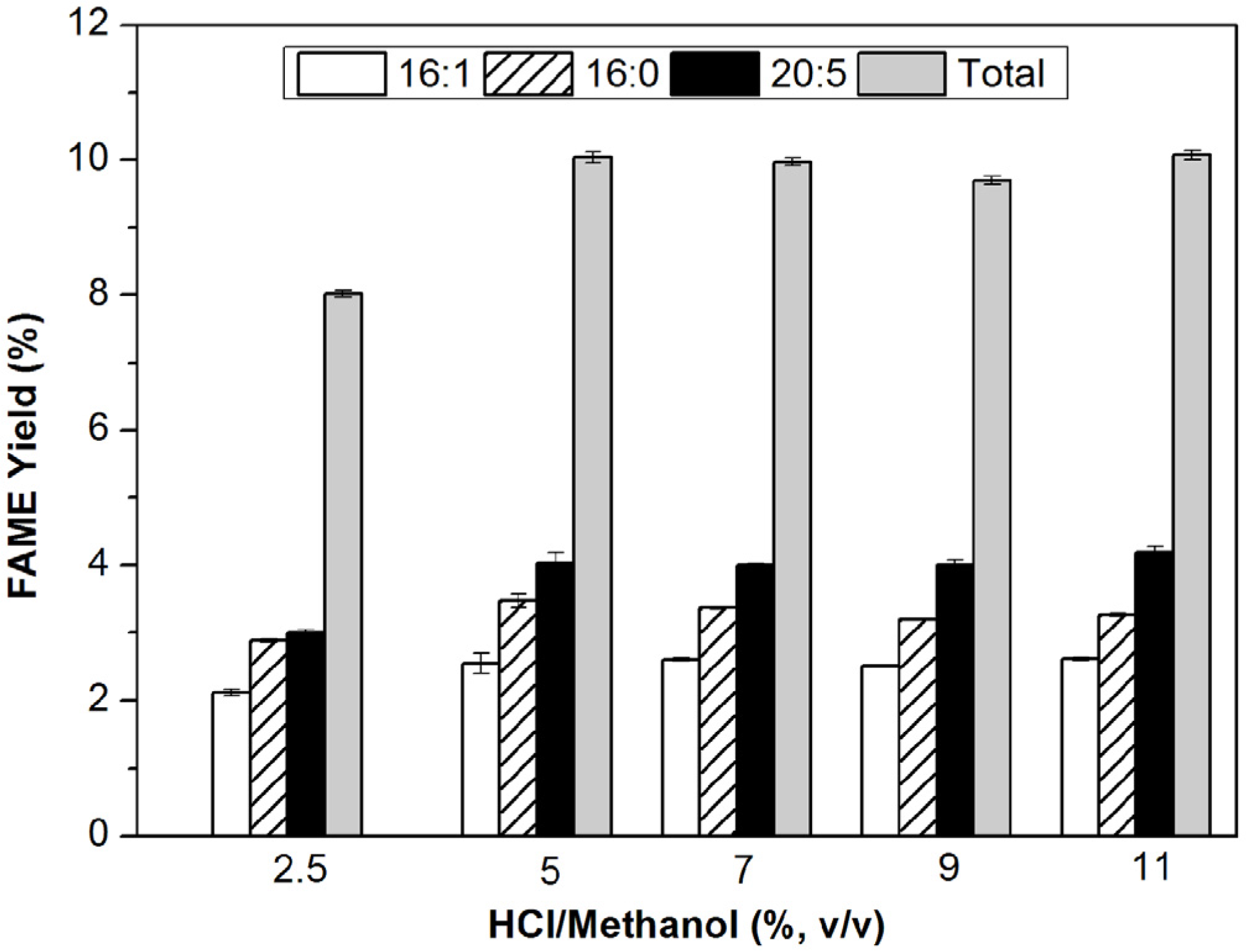
3.2.2. Effect of Acid Concentration on Reaction Efficiency
3.2.3. Effect of Reaction Time on Reaction Efficiency
3.2.4. Effect of Reaction Temperature on Reaction Efficiency
3.2.5. Effect of Extraction Solvent on Gravimetric Lipid Content and Total Fatty Acid Yields
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ahmad, A.L.; Yasin, N.H.M.; Derek, C.J.C.; Lim, J.K. Microalgae as a sustainable energy source for biodiesel production: A review. Renew. Sustain. Energy Rev. 2011, 15, 584–593. [Google Scholar] [CrossRef]
- Pires, J.C.M.; Alvim-Ferraz, M.C.M.; Martins, F.G.; Simões, M. Wastewater treatment to enhance the economic viability of microalgae culture. Environ. Sci. Pollut. Res. 2013, 20, 5096–5105. [Google Scholar] [CrossRef] [PubMed]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Goiris, K.; Muylaert, K.; Foubert, I. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, C.H.S.; Calder, P.C.; Reed, S.C.; Simpson, M.J.A. The impact of long-chain n-3 polyunsaturated fatty acids on human health. Nutr. Res. Rev. 2005, 18, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.L.; Lynch, B.; Barfield, W.; Bull, A.; Ryan, A.S.; Astwood, J.D. Genetic and acute toxicological evaluation of an algal oil containing eicosapentaenoic acid (epa) and palmitoleic acid. Food Chem. Toxicol. 2014, 72, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Chauton, M.S.; Reitan, K.I.; Norsker, N.H.; Tveterås, R.; Kleivdal, H.T. A techno-economic analysis of industrial production of marine microalgae as a source of epa and dha-rich raw material for aquafeed: Research challenges and possibilities. Aquaculture 2015, 436, 95–103. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, Z.; Yu, C.; Yin, Y.; Zhou, G. Evaluation of the potential of 9 nannochloropsis strains for biodiesel production. Bioresour. Technol. 2014, 167, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Campos, H.; Boeing, W.J.; Dungan, B.N.; Schaub, T. Cultivating the marine microalga nannochloropsis salina under various nitrogen sources: Effect on biovolume yields, lipid content and composition, and invasive organisms. Biomass Bioenergy 2014, 66, 301–307. [Google Scholar] [CrossRef]
- Wan, C.; Bai, F.-W.; Zhao, X.-Q. Effects of nitrogen concentration and media replacement on cell growth and lipid production of oleaginous marine microalga nannochloropsis oceanica dut01. Biochem. Eng. J. 2013, 78, 32–38. [Google Scholar] [CrossRef]
- Mayers, J.J.; Flynn, K.J.; Shields, R.J. Influence of the n:P supply ratio on biomass productivity and time-resolved changes in elemental and bulk biochemical composition of nannochloropsis sp. Bioresour. Technol. 2014, 169, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, H.; Abdu Rahman, N.; Banerjee, S.; Harun, N.; Suleiman, S.S.; Zakaria, N.H.; Lananan, F.; Abdul Hamid, S.H.; Endut, A. Effects of different salinities and ph on the growth and proximate composition of nannochloropsis sp. And tetraselmis sp. Isolated from south china sea cultured under control and natural condition. Int. Biodeterior. Biodegrad. 2014, 95 Part A, 11–18. [Google Scholar] [CrossRef]
- Simionato, D.; Sforza, E.; Corteggiani Carpinelli, E.; Bertucco, A.; Giacometti, G.M.; Morosinotto, T. Acclimation of nannochloropsis gaditana to different illumination regimes: Effects on lipids accumulation. Bioresour. Technol. 2011, 102, 6026–6032. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ning, K.; Li, J.; Hu, J.; Han, D.; Wang, H.; Zeng, X.; Jing, X.; Zhou, Q.; Su, X.; et al. Nannochloropsis genomes reveal evolution of microalgal oleaginous traits. PLoS Genet. 2014, 10, e1004094. [Google Scholar] [CrossRef] [PubMed]
- Mercer, P.; Armenta, R.E. Developments in oil extraction from microalgae. Eur. J. Lipid Sci. Technol. 2011, 113, 539–547. [Google Scholar] [CrossRef]
- McNichol, J.; MacDougall, K.; Melanson, J.; McGinn, P. Suitability of soxhlet extraction to quantify microalgal fatty acids as determined by comparison with in situ transesterification. Lipids 2012, 47, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Carrapiso, A.; García, C. Development in lipid analysis: Some new extraction techniques and in situ transesterification. Lipids 2000, 35, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Laurens, L.L.; Quinn, M.; Van Wychen, S.; Templeton, D.; Wolfrum, E. Accurate and reliable quantification of total microalgal fuel potential as fatty acid methyl esters by in situ transesterification. Anal. Bioanal. Chem. 2012, 403, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L.; Jenkins, T.C. Challenges with fats and fatty acid methods. J. Anim. Sci. 2003, 81, 3250–3254. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-L.; Huang, C.-C.; Ho, K.-C.; Hsiao, P.-X.; Wu, M.-S.; Chang, J.-S. Biodiesel production from wet microalgae feedstock using sequential wet extraction/transesterification and direct transesterification processes. Bioresour. Technol. 2015, 194, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.N.; Kobayashi, N.; Barnes, A.; Noel, E.A.; Betenbaugh, M.J.; Oyler, G.A. Comparative analyses of three chlorella species in response to light and sugar reveal distinctive lipid accumulation patterns in the microalga c. Sorokiniana. PLoS ONE 2014, 9, e92460. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.-Y.; Kwon, J.-H.; Jo, G.-J. Removal of algae and turbidity by floating-media and sand filtration. Desalin. Water Treat. 2013, 52, 1007–1013. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Volkman, J.K.; Barrett, S.M.; Dunstan, G.A.; Jeffrey, S.W. C30-c32 alkyl diols and unsaturated alcohols in microalgae of the class eustigmatophyceae. Organ. Geochem. 1992, 18, 131–138. [Google Scholar] [CrossRef]
- Shiu, P.-J.; Gunawan, S.; Hsieh, W.-H.; Kasim, N.S.; Ju, Y.-H. Biodiesel production from rice bran by a two-step in-situ process. Bioresour. Technol. 2010, 101, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.N.; Oyler, G.A.; Wilkinson, L.; Betenbaugh, M.J. A green light for engineered algae: Redirecting metabolism to fuel a biotechnology revolution. Curr. Opin. Biotechnol. 2008, 19, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Pieber, S.; Schober, S.; Mittelbach, M. Pressurized fluid extraction of polyunsaturated fatty acids from the microalga nannochloropsis oculata. Biomass Bioenergy 2012, 47, 474–482. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bermúdez, S.; Termote-Verhalle, R.; Bruneel, C.; Muylaert, K.; Parra-Saldivar, R.; Foubert, I. Influence of extraction solvent system on the extractability of lipid components from the biomass of nannochloropsis gaditana. J. Appl. Phycol. 2014, 26, 1501–1510. [Google Scholar] [CrossRef]
- Camacho-Rodríguez, J.; Cerón-García, M.C.; Fernández-Sevilla, J.M.; Molina-Grima, E. The influence of culture conditions on biomass and high value product generation by nannochloropsis gaditana in aquaculture. Algal Res. 2015, 11, 63–73. [Google Scholar] [CrossRef]
- Chen, Y.; Vaidyanathan, S. Simultaneous assay of pigments, carbohydrates, proteins and lipids in microalgae. Anal. Chim. Acta 2013, 776, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-Y.; Chen, F. Heterotrophic production of eicosapentaenoic acid by microalgae. Biotechnol. Adv. 2003, 21, 273–294. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Mitra, M.; Patidar, S.K.; George, B.; Shah, F.; Mishra, S. A euryhaline nannochloropsis gaditana with potential for nutraceutical (epa) and biodiesel production. Algal Res. 2015, 8, 161–167. [Google Scholar] [CrossRef]
- Kim, C.W.; Sung, M.-G.; Nam, K.; Moon, M.; Kwon, J.-H.; Yang, J.-W. Effect of monochromatic illumination on lipid accumulation of nannochloropsis gaditana under continuous cultivation. Bioresour. Technol. 2014, 159, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Noel, E.; Barnes, A.; Rosenberg, J.; DiRusso, C.; Black, P.; Oyler, G. Rapid detection and quantification of triacylglycerol by hplc–elsd in chlamydomonas reinhardtii and chlorella strains. Lipids 2013, 48, 1035–1049. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of FAME | Amount (Peak Area %) |
|---|---|
| 14:0 | 3.11 ± 0.06 |
| 16:1 | 18.34 ± 0.17 |
| 16:0 | 26.60 ± 0.20 |
| 18:0 | 6.78 ± 0.04 |
| 18:1 & 18:2 | 8.95 ± 0.04 |
| 20:5 | 34.46 ± 0.43 |
| Others | 1.75 ± 0.03 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Zhang, Y.; Rosenberg, J.N.; Betenbaugh, M.J.; Wang, F. Optimization of One-Step In Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana. Appl. Sci. 2016, 6, 343. https://doi.org/10.3390/app6110343
Tang Y, Zhang Y, Rosenberg JN, Betenbaugh MJ, Wang F. Optimization of One-Step In Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana. Applied Sciences. 2016; 6(11):343. https://doi.org/10.3390/app6110343
Chicago/Turabian StyleTang, Yuting, Yue Zhang, Julian N. Rosenberg, Michael J. Betenbaugh, and Fei Wang. 2016. "Optimization of One-Step In Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana" Applied Sciences 6, no. 11: 343. https://doi.org/10.3390/app6110343
APA StyleTang, Y., Zhang, Y., Rosenberg, J. N., Betenbaugh, M. J., & Wang, F. (2016). Optimization of One-Step In Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana. Applied Sciences, 6(11), 343. https://doi.org/10.3390/app6110343