Contribution of Dopamine Transporter Gene Methylation Status to Cannabis Dependency

 , , , , ,
, , , , ,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
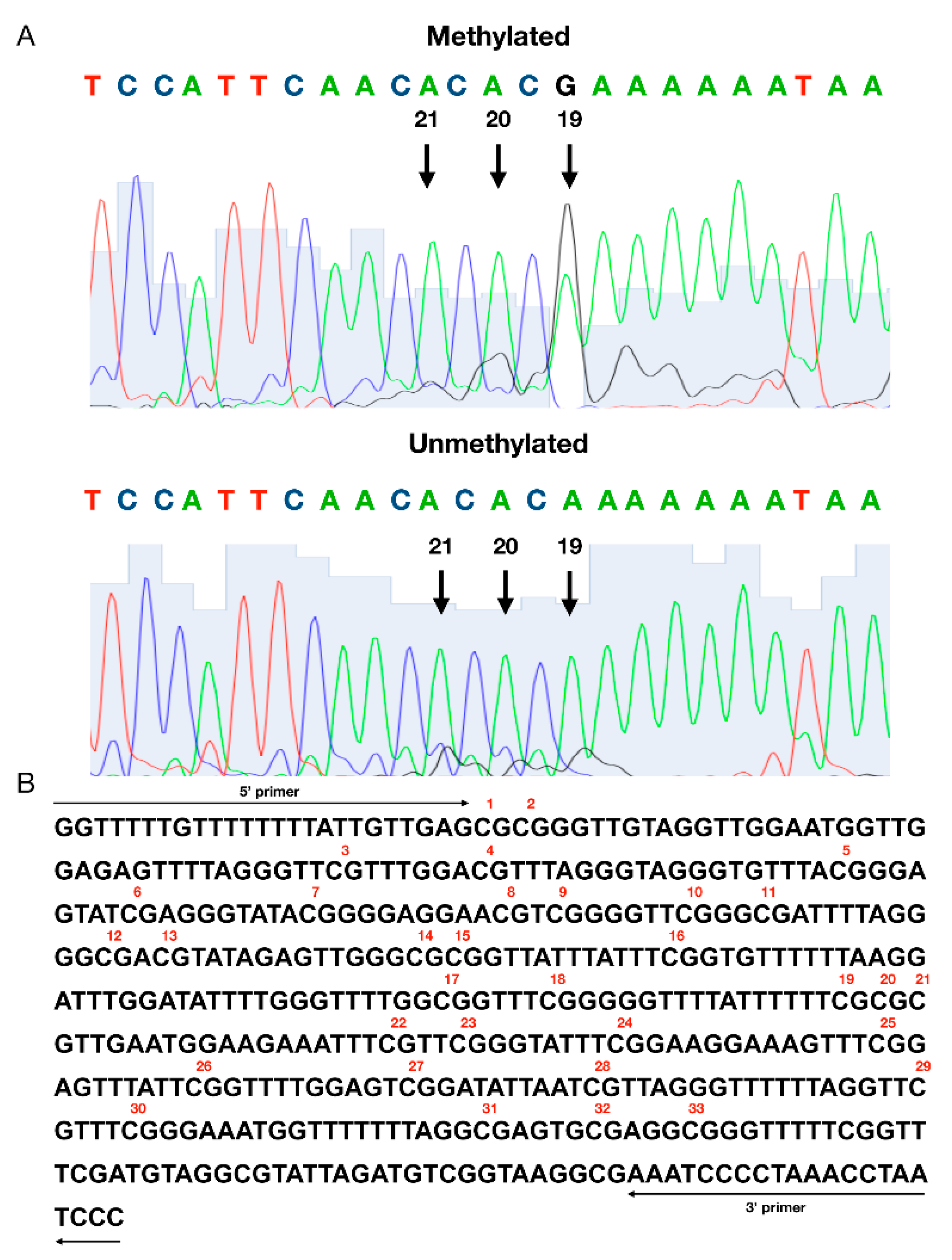
2.2. Methylation Status Assessment
2.3. Assessment of the Ability to Bind Transcription Factors
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef]
- Van Speybroeck, L. From epigenesis to epigenetics. Ann. N. Y. Acad. Sci. 2002, 981, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Nagre, N.N.; Subbanna, S.; Shivakumar, M.; Psychoyos, D.; Basavarajappa, B.S. CB1-receptor knockout neonatal mice are protected against ethanolinduced impairments of DNMT1, DNMT3A, and DNA methylation. J. Neurochem. 2015, 132, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Renthal, W.; Nestler, E.J. Epigenetic mechanisms in drug addiction. Trends Mol. Med. 2008, 14, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Maze, I.; Nestler, E.J. The epigenetic landscape of addiction. Ann. N. Y. Acad. Sci. 2011, 1216, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Baedke, J. The epigenetic landscape in the course of time: Conrad Hal Waddington’s methodological impact on the life sciences. Stud. Hist. Philos. Sci. Part C 2013, 44, 756–773. [Google Scholar] [CrossRef]
- Szutorisz, H.; Hurd, Y.L. Epigenetic effects of cannabis exposure. Biol. Psychiatry 2016, 79, 586–594. [Google Scholar] [CrossRef]
- Blankman, J.L.; Simon, G.M.; Cravatt, B.F. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem. Biol. 2007, 14, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Gao, S.; Qi, W.; Shi, C.; Qiu, M.; Yang, F.; Bai, S.; Li, H.; Wang, Z.; Sun, Z.; et al. 5-Hydroxymethylcytosine as a potential epigenetic biomarker in papillary thyroid carcinoma. Oncol. Lett. 2019, 18, 2304–2309. [Google Scholar] [CrossRef]
- Wilson, M.E.; Sengoku, T. Developmental regulation of neuronal genes by DNA methylation: Environmental influences. Int. J. Dev. Neurosci. 2013, 31, 448–451. [Google Scholar] [CrossRef]
- Watson, C.T.; Szutorisz, H.; Garg, P.; Martin, Q.; Landry, J.A.; Sharp, A.J.; Hurd, Y.L. Genome-wide DNA methylation profiling reveals epigenetic changes in the rat nucleus accumbens associated with crossgenerational effects of adolescent THC exposure. Neuropsychopharmacology 2015, 40, 2993–3005. [Google Scholar] [CrossRef] [PubMed]
- Spiers, H.; Hannon, E.; Schalkwyk, L.C.; Smith, R.; Wong, C.C.Y.; O’Donovan, M.C.; Bray, N.J.; Mill, J. Methylomic trajectories across human fetal brain development. Genome Res. 2015, 25, 338–352. [Google Scholar] [CrossRef]
- Schmitt, A.; Malchow, B.; Hasan, A.; Falkai, P. The impact of environmental factors in severe psychiatric disorders. Front. Neurosci. 2014, 8, 19. [Google Scholar] [CrossRef]
- Tuesta, L.M.; Zhang, Y. Mechanisms of epigenetic memory and addiction. EMBO J. 2014, 33, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Nestler, E.J. Epigenetic mechanisms of drug addiction. Curr. Opin. Neurobiol. 2013, 23, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C. The epidemiology of cannabis dependence. In Cannabis Dependence: Its Nature, Consequences and Treatment; Roffman, R., Stephens, R.S., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 58–105. [Google Scholar]
- Gunn, J.K.; Rosales, C.B.; Center, K.E.; Nunez, A.; Gibson, S.J.; Christ, C.; Ehiri, J.E. Prenatal exposure to cannabis and maternal and child health outcomes: A systematic review and meta-analysis. BMJ Open 2016, 6, e009986. [Google Scholar] [CrossRef]
- Yang, X.; Hegde, V.L.; Rao, R.; Zhang, J.; Nagarkatti, P.S.; Nagarkatti, M. Histone modifications are associated with D9-tetrahydrocannabinol-mediated alterations in antigen-specific t cell responses. J. Biol. Chem. 2014, 289, 18707–18718. [Google Scholar] [CrossRef]
- Szutorisz, H.; DiNieri, J.A.; Sweet, E.; Egervari, G.; Michaelides, M.; Carter, J.M.; Ren, Y.; Miller, M.L.; Blitzer, R.D.; Hurd, Y.L. Parental THC exposure leads to compulsive heroin-seeking and altered striatal synaptic plasticity in the subsequent generation. Neuropsychopharmacology 2014, 39, 1315–1323. [Google Scholar] [CrossRef]
- Gerra, M.C.; Jayanthi, S.; Manfredini, M.; Walther, D.; Schroeder, J.; Philips, K.A.; Cadet, J.L.; Donnini, C. Gene variants and educational attainment in cannabis use: Mediating role of DNA methylation. Transl. Psychiatry 2018, 8, 23. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Jones, S.R.; Fumagalli, F.; Wightman, R.M.; Caron, M.G. Re-evaluation of the role of the dopamine transporter in dopamine system homeostasis. Brain Res. Rev. 1998, 26, 148–153. [Google Scholar] [CrossRef]
- Jasiewicz, A.; Rubiś, B.; Samochowiec, J.; Małecka, I.; Suchanecka, A.; Jabłoński, M.; Grzywacz, A. DAT1 methylation changes in alcohol-dependent individuals vs. controls. J. Psych. Res. 2015, 64, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Hillemacher, T.; Frieling, H.; Hartl, T.; Wilhelm, J.; Kornhuber, J.; Bleich, S. Promoter specific methylation of the dopamine trans-porter gene is altered in alcohol dependence and associated with craving. J. Psychiatr. Res. 2009, 43, 388–392. [Google Scholar] [CrossRef]
- Muench, C.; Wiers, C.E.; Cortes, C.R.; Momenan, R.; Lohoff, F.W. Dopamine Transporter Gene Methylation is Associated with Nucleus Accumbens Activation During Reward Processing in Healthy but not Alcohol-Dependent Individuals. Alcohol. Clin. Exp. Res. 2018, 42, 21–31. [Google Scholar] [CrossRef]
- Wiers, C.; Lohoff, F.W.; Lee, J.; Muench, C.; Freeman, C.; Zehra, A.; Marenco, S.; Lipska, B.K.; Auluck, P.K.; Feng, N.; et al. Methylation of the dopamine transporter gene in blood is associated with striatal dopamine transporter availability in ADHD: A preliminary study. Eur. J. Neurosci. 2018, 48, 1884–1895. [Google Scholar] [CrossRef] [PubMed]
- Zoratto, F.; Romano, E.; Pascale, E.; Pucci, M.; Falconi, A.; Dell’Osso, B.; Maccarrone, M.; Laviola, G.; D’Addario, C.; Adriani, W. Down-regulation of serotonin and dopamine transporter genes in individual rats expressing a gambling-prone profile: A possible role for epigenetic mechanisms. Neuroscience 2017, 340, 101–116. [Google Scholar] [CrossRef]
- Rajala, A.Z.; Zaitoun, I.; Henriques, J.B.; Converse, A.K.; Murali, D.; Epstein, M.L.; Populin, L.C. Dopamine teransporter gene susceptibility to methylation is associated with impulsivity in nonhuman primates. J. Neurophysiol. 2014, 112, 2138–2146. [Google Scholar] [CrossRef]
- Terry, M.B.; Delgado-Cruzata, L.; Vin-Raviv, N.; Wu, H.C.; Santella, R.M. DNA methylation in white blood cells: Association with risk factors in epidemiologic studies. Epigenetics 2011, 6, 828–837. [Google Scholar] [CrossRef]
- Li, L.; Zheng, H.; Huang, Y.; Huang, C.; Zhang, S.; Tian, J.; Li, P.; Sood, A.K.; Zhang, W.; Chen, K. DNA methylation signatures and coagulation factors in the peripheral blood leucocytes of epithelial ovarian cancer. Carcinogenesis 2017, 38, 797–805. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, G.; Qian, J. Transcription factors as readers and effectors of DNA methylation. Nat. Rev. Genet. 2016, 17, 551–565. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=5079 (accessed on 15 May 2020).
- Adams, B.; Dörfler, P.; Aguzzi, A.; Kozmik, Z.; Urbánek, P.; Maurer-Fogy, I.; Busslinger, M. Pax-5 encodes the transcription factor BSAP and is expressed in B lymphocytes, the developing CNS, and adult testis. Genes Dev. 1992, 6, 1589–1607. [Google Scholar] [CrossRef]
- Goldfarb, E.V.; Sinha, R. Drug-induced glucocorticoids and memory for substance use. Trends Neurosci. 2018, 41, 853–868. [Google Scholar] [CrossRef] [PubMed]
- Deroche-Gamonet, V.; Sillaber, I.; Aouizerate, B.; Izawa, R.; Jaber, M.; Ghozland, S.; Kellendonk, C.; Le Moal, M.; Spanagel, R.; Schütz, G.; et al. The glucocorticoid receptor as a potential target to reduce cocaine abuse. J. Neurosci. 2003, 23, 4785–4790. [Google Scholar] [CrossRef] [PubMed]
- Madalena, K.M.; Lerch, J.K. The Effect of Glucocorticoid and Glucocorticoid Receptor Interactions on Brain, Spinal Cord, and Glial Cell Plasticity. Neural Plast. 2017, 2017, 8640970. [Google Scholar] [CrossRef] [PubMed]
- Vassoler, F.M.; White, S.L.; Schmidt, H.D.; Sadri-Vakili, G.; Pierce, R.C. Epigenetic inheritance of a cocaine resistance phenotype. Nat. Neurosci. 2013, 16, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Landthaler, M.; Schlussman, S.D.; Yuferov, V.; Ho, A.; Tuschl, T.; Kreek, M.J. Mu opioid receptor knockdown in the substantia nigra/ventral tegmental area by synthetic small interfering RNA blocks the rewarding and locomotor effects of heroin. Neuroscience 2009, 158, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Lasek, A.W.; Janak, P.H.; He, L.; Whistler, J.L.; Heberlein, U. Downregulation of mu opioid receptor by RNA interference in the ventral tegmental area reduces ethanol consumption in mice. Genes Brain Behav. 2007, 6, 728–735. [Google Scholar] [CrossRef]
- Sun, Y.V.; Turner, S.T.; Smith, J.A.; Hammond, P.I.; Lazarus, A.; van de Rostyne, J.L.; Cunningham, J.M.; Kardia, S.L. Comparison of the DNA methylation profiles of human peripheral blood cells and transformed B lymphocytes. Hum. Genet. 2010, 127, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Prini, P.; Penna, F.; Sciuccati, E.; Alberio, T.; Rubino, T. Chronic Δ⁸-THC Exposure Differently Affects Histone Modifications in the Adolescent and Adult Rat Brain. Int. J. Mol. Sci. 2017, 18, 2094. [Google Scholar] [CrossRef]
- Prini, P.; Rusconi, F.; Zamberletti, E.; Gabaglio, M.; Penna, F.; Fasano, M.; Battaglioli, E.; Parolaro, D.; Rubino, T. Adolescent THC exposure in female rats leads to cognitive deficits through a mechanism involving chromatin modifications in the prefrontal cortex. J. Psychiatry Neurosci. 2017, 42, 170082. [Google Scholar] [CrossRef] [PubMed]
- Kordi-Tamandani, D.M.; Tajoddini, S.; Salimi, F. Promoter Methylation and BDNF and DAT1 Gene Expression Profiles in Patients with Drug Addiction. Pathobiology 2015, 82, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.; Dempster, E.L.; Illott, N.; Chabrawi, S.; Maior, R.S.; Tomaz, C.; Silva, M.A.; Huston, J.P.; Mill, J.; Müller, C.P. Decreased methylation of the NK3 receptor coding gene (TACR3) after cocaine-induced place preference in marmoset monkeys. Addict. Biol. 2013, 18, 452–454. [Google Scholar] [CrossRef]
- Lohoff, F.W.; Bloch, P.J.; Hodge, R.; Nall, A.H.; Ferraro, T.N.; Kampman, K.M.; Dackis, C.A.; O’Brien, C.P.; Pettinati, H.M.; Oslin, D.W. Association analysis between polymorphisms in the dopamine D2 receptor (DRD2) and dopamine transporter (DAT1) genes with cocaine dependence. Neurosci. Lett. 2010, 473, 87–91. [Google Scholar] [CrossRef]
- Wetherill, R.R.; Jagannathan, K.; Lohoff, F.W.; Ehrman, R.; O’Brien, C.P.; Childress, A.R.; Franklin, T.R. Neural correlates of attentional bias for smoking cues: Modulation by variance in the dopamine transporter gene. Addict. Biol. 2014, 19, 294–304. [Google Scholar] [CrossRef]


{kind=link}
| CpG Site | Studied Group | Methylation Status (%) | χ2(p) | OR | 95% CI (−95%, +95%) | Spearman’s R (p) |
|---|---|---|---|---|---|---|
| 1 * | dependent N (201) | 62% | 11.689 (0.001) | 0.528 | (0.365, 0.763) | −0.155 (0.001) |
| control N (285) | 46% | |||||
| 2 | dependent N (201) | 78% | 1.992 (0.158) | 0.739 | (0.485, 1.125) | −0.064 (0.159) |
| control N (285) | 72% | |||||
| 3 | dependent N (201) | 86% | 4.986 (0.026) | 1.921 | (1.075, 3.431) | 0.101 (0.026) |
| control N (285) | 92% | |||||
| 4 | dependent N (201) | 25% | 0.003 (0.952) | 1.013 | (0.669, 1.533) | 0.003 (0.952) |
| control N (285) | 26% | |||||
| 5 | dependent N (201) | 26% | 5.357 (0.021) | 1.597 | (1.072, 2.377) | 0.105 (0.021) |
| control N (285) | 36% | |||||
| 6 * | dependent N (201) | 18% | 16.044 (0.0001) | 0.309 | (0.170, 0.562) | −0.182 (0.0001) |
| control N (285) | 6% | |||||
| 7 | dependent N (201) | 15% | 0.289 (0.591) | 0.869 | (0.522, 1.448) | −0.024 (0.592) |
| control N (285) | 14% | |||||
| 8 | dependent N (201) | 3% | 1.499 (0.221) | 1.812 | (0.691, 4.754) | 0.056 (0.222) |
| control N (285) | 5% | |||||
| 9 | dependent N (201) | 36% | 0.217 (0.641) | 1.093 | (0.751, 1.590) | 0.021 (0.642) |
| control N (285) | 38% | |||||
| 10 | dependent N (201) | 37% | 0.00001 (0.998) | 1.000 | (0.689, 1.453) | 0.0001 (0.998) |
| control N (285) | 37% | |||||
| 11 | dependent N (201) | 3% | 2.360 (0.124) | 1.979 | (0.816, 4.802) | 0.069 (0.125) |
| control N (285) | 7% | |||||
| 12 | dependent N (201) | 30% | 0.105 (0.746) | 1.067 | (0.721, 1.580) | 0.015 (0.746) |
| control N (285) | 31% | |||||
| 13 | dependent N (201) | 2% | 8.037 (0.005) | 3.769 | (1.417, 10.022) | 0.128 (0.005) |
| control N (285) | 9% | |||||
| 14 | dependent N (201) | 85% | 0.046 (0.832) | 0.947 | (0.577, 1.556) | −0.010 (0.831) |
| control N (285) | 84% | |||||
| 15 | dependent N (201) | 83% | 0.019 (0.891) | 0.967 | (0.602, 1.554) | −0.006 (0.891) |
| control N (285) | 82% | |||||
| 16 | dependent N (201) | 67% | 2.594 (0.107) | 0.733 | (0.502, 1.070) | −0.073 (0.108) |
| control N (285) | 60% | |||||
| 17 | dependent N (201) | 28% | 0.266 (0.606) | 1.110 | (0.746, 1.651) | 0.023 (0.607) |
| control N (285) | 31% | |||||
| 18 | dependent N (201) | 7% | 0.002 (0.969) | 0.986 | (0.495, 1.964) | −0.002 (0.969) |
| control N (285) | 7% | |||||
| 19 | dependent N (201) | 92% | 7.920 (0.005) | 3.435 | (1.386, 8.511) | 0.128 (0.005) |
| control N (285) | 98% | |||||
| 20 | dependent N (201) | 45% | 2.558 (0.110) | 0.741 | (0.513, 1.070) | −0.072 (0.110) |
| control N (285) | 38% | |||||
| 21 | dependent N (201) | 72% | 2.441 (0.118) | 0.732 | (0.495, 1.083) | −0.071 (0.119) |
| control N (285) | 65% | |||||
| 22 | dependent N (201) | 87% | 10.045 (0.002) | 2.876 | (1.461, 5.659) | 0.144 (0.001) |
| control N (285) | 95% | |||||
| 23 | dependent N (201) | 19% | 0.080 (0.777) | 0.935 | (0.587, 1.489) | −0.013 (0.777) |
| control N (285) | 18% | |||||
| 24 | dependent N (201) | 70% | 0.284 (0.594) | 0.899 | (0.609, 1.328) | −0.024 (0.595) |
| control N (285) | 67% | |||||
| 25 | dependent N (201) | 25% | 5.761 (0.016) | 1.632 | (1.092, 2.440) | 0.109 (0.016) |
| control N (285) | 35% | |||||
| 26 | dependent N (201) | 37% | 2.549 (0.110) | 1.350 | (0.933, 1.953) | 0.072 (0.111) |
| control N (285) | 45% | |||||
| 27 | dependent N (201) | 17% | 0.143 (0.705) | 1.096 | (0.681, 1.764) | 0.017 (0.706) |
| control N (285) | 18% | |||||
| 28 | dependent N (201) | 73% | 6.067 (0.014) | 0.611 | (0.412, 0.906) | −0.112 (0.014) |
| control N (285) | 62% | |||||
| 29 | dependent N (201) | 25% | 1.888 (0.169) | 0.738 | (0.479, 1.139) | −0.062 (0.170) |
| control N (285) | 20% | |||||
| 30 | dependent N (201) | 10% | 0.074 (0.786) | 1.084 | (0.605, 1.941) | 0.012 (0.786) |
| control N (285) | 11% | |||||
| 31 | dependent N (201) | 5% | 0.290 (0.590) | 1.234 | (0.574, 2.653) | 0.024 (0.591) |
| control N (285) | 7% | |||||
| 32 | dependent N (201) | 68% | 0.004 (0.951) | 1.012 | (0.686, 1.492) | 0.003 (0.951) |
| control N (285) | 68% | |||||
| 33 | dependent N (201) | 73% | 2.599 (0.107) | 1.413 | (0.927, 2.152) | 0.073 (0.107) |
| control N (285) | 79% |
| Matrix Similarity Rate | CpG Position | Transcription Factor |
|---|---|---|
| (a) 100% | 3 | PAX5 |
| 33 | PAX5 | |
| (b) 95% | 1 | GCF |
| 3 | PAX5 | |
| 11 | RXR-alpha | |
| 19 | c-Ets-2 | |
| 20 | c-Ets-2 | |
| 22 | PAX5, p53, Sp1 | |
| 25 | AP-2alphaA | |
| 28 | NFI/CTF | |
| 33 | PAX5 | |
| (c) 85% | 1 | GCF, E2F1 |
| 3 | PAX5, p53 | |
| 5 | AhR | |
| 6 | GR alpha | |
| 11 | TFII-I, STAT4, RXR-alpha | |
| 13 | PAX5, p53 | |
| 19 | GR alpha, c-Ets-2, E2F1, GCF | |
| 20 | c-Ets-2, E2F1, GCF | |
| 22 | PAX5, p53, E2F1, Sp1 | |
| 25 | GR alpha, AP-2alphaA, NF-AT2 | |
| 26 | PAX5, p53 | |
| 28 | ENKTF1, EBF, E2F1, NFI/CTF | |
| 33 | PAX5, p53, E2F1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzywacz, A.; Barczak, W.; Chmielowiec, J.; Chmielowiec, K.; Suchanecka, A.; Trybek, G.; Masiak, J.; Jagielski, P.; Grocholewicz, K.; Rubiś, B. Contribution of Dopamine Transporter Gene Methylation Status to Cannabis Dependency. Brain Sci. 2020, 10, 400. https://doi.org/10.3390/brainsci10060400
Grzywacz A, Barczak W, Chmielowiec J, Chmielowiec K, Suchanecka A, Trybek G, Masiak J, Jagielski P, Grocholewicz K, Rubiś B. Contribution of Dopamine Transporter Gene Methylation Status to Cannabis Dependency. Brain Sciences. 2020; 10(6):400. https://doi.org/10.3390/brainsci10060400
Chicago/Turabian StyleGrzywacz, Anna, Wojciech Barczak, Jolanta Chmielowiec, Krzysztof Chmielowiec, Aleksandra Suchanecka, Grzegorz Trybek, Jolanta Masiak, Paweł Jagielski, Katarzyna Grocholewicz, and Blazej Rubiś. 2020. "Contribution of Dopamine Transporter Gene Methylation Status to Cannabis Dependency" Brain Sciences 10, no. 6: 400. https://doi.org/10.3390/brainsci10060400
APA StyleGrzywacz, A., Barczak, W., Chmielowiec, J., Chmielowiec, K., Suchanecka, A., Trybek, G., Masiak, J., Jagielski, P., Grocholewicz, K., & Rubiś, B. (2020). Contribution of Dopamine Transporter Gene Methylation Status to Cannabis Dependency. Brain Sciences, 10(6), 400. https://doi.org/10.3390/brainsci10060400