
Predicting Math Ability Using Working Memory, Number Sense, and Neurophysiology in Children and Adults

 , ,
, ,
Abstract
:1. Introduction
1.1. Age Differences in Processes Underlying Mathematical Ability
1.2. Electrophysiological Correlates of Mathematical Ability
1.3. Analyzing Beta Oscillations
1.4. The Present Study
2. Materials and Methods
2.1. Participants and Ethics Statement
2.2. Overview of Experimental Paradigm and Stimuli
2.2.1. Resting-State Electroencephalography Recordings and Pre-Processing
2.2.2. Arithmetic Ability
2.2.3. Cognitive Measurements
Number Sense
Working Memory
2.3. EEG Analysis
Resting-State Beta Activity (Traditional and Parameterization)
2.4. Data Analysis
3. Results
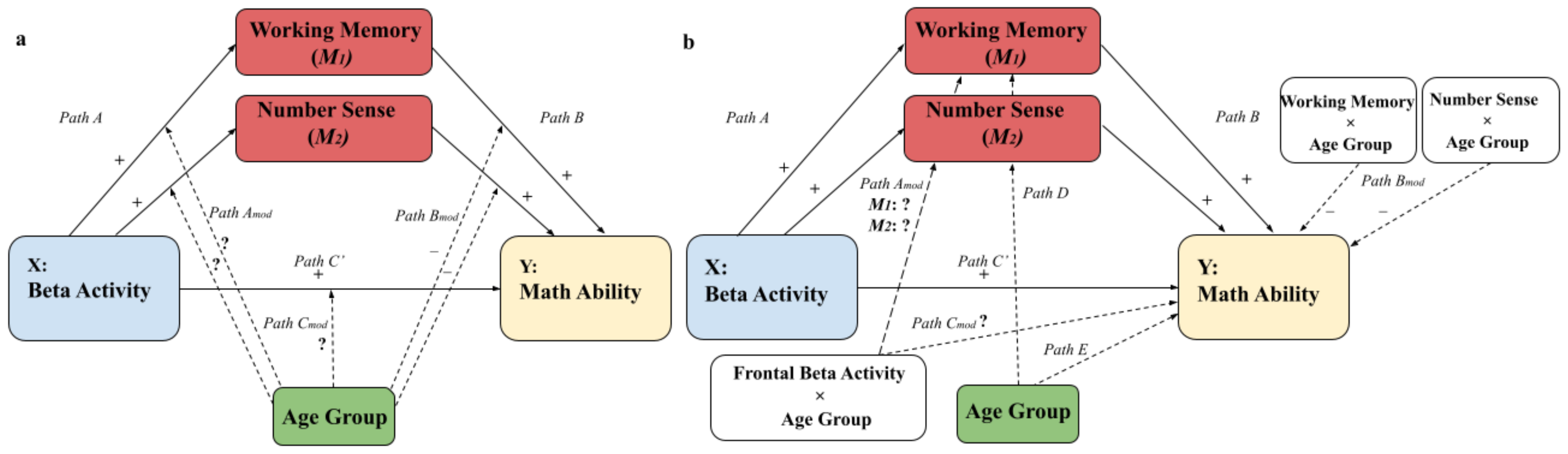
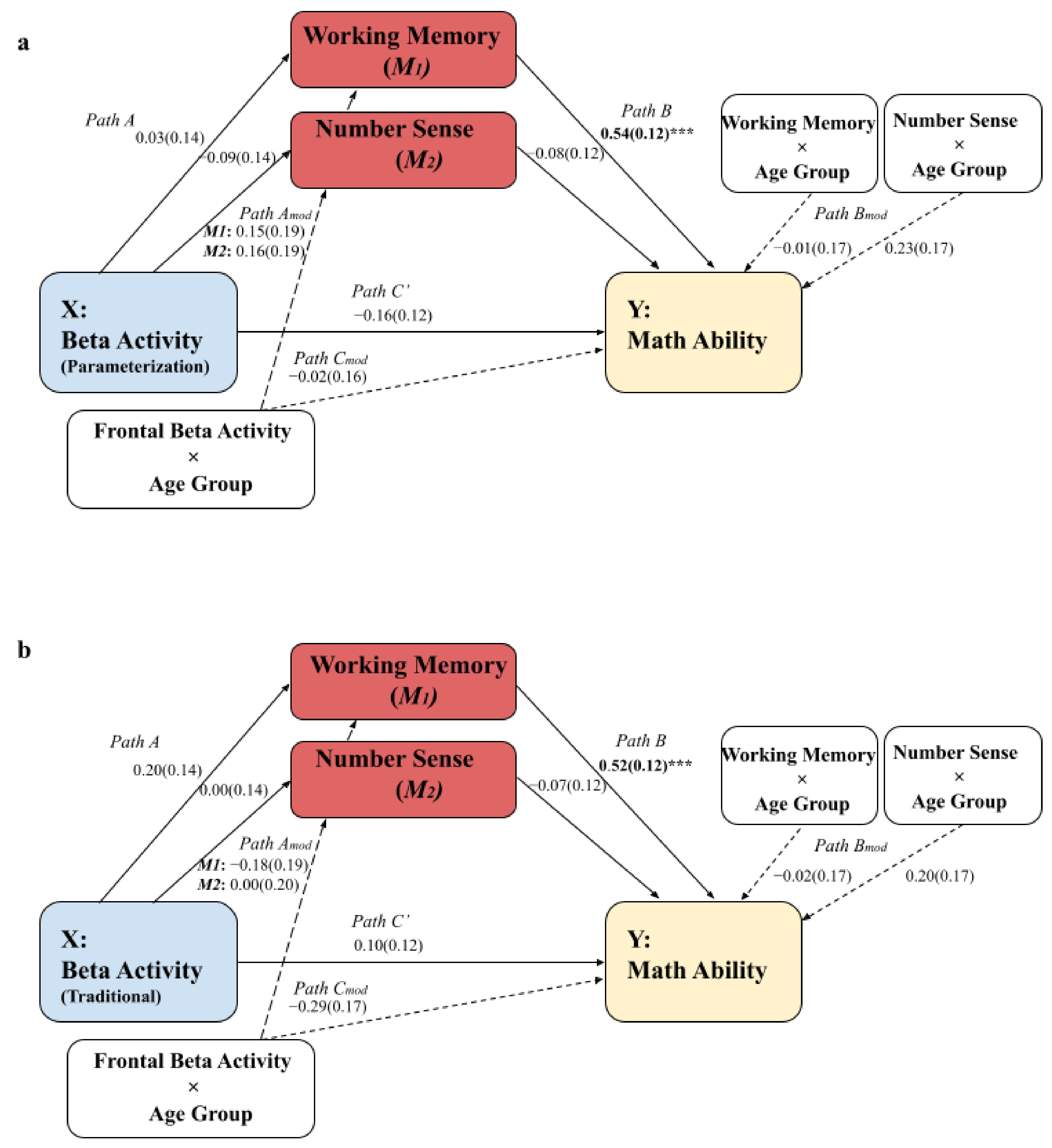
3.1. Moderated Mediation Models: The Parameterization Method and the Traditional Method
3.2. Comparisons of the Spectral Calculation Methods
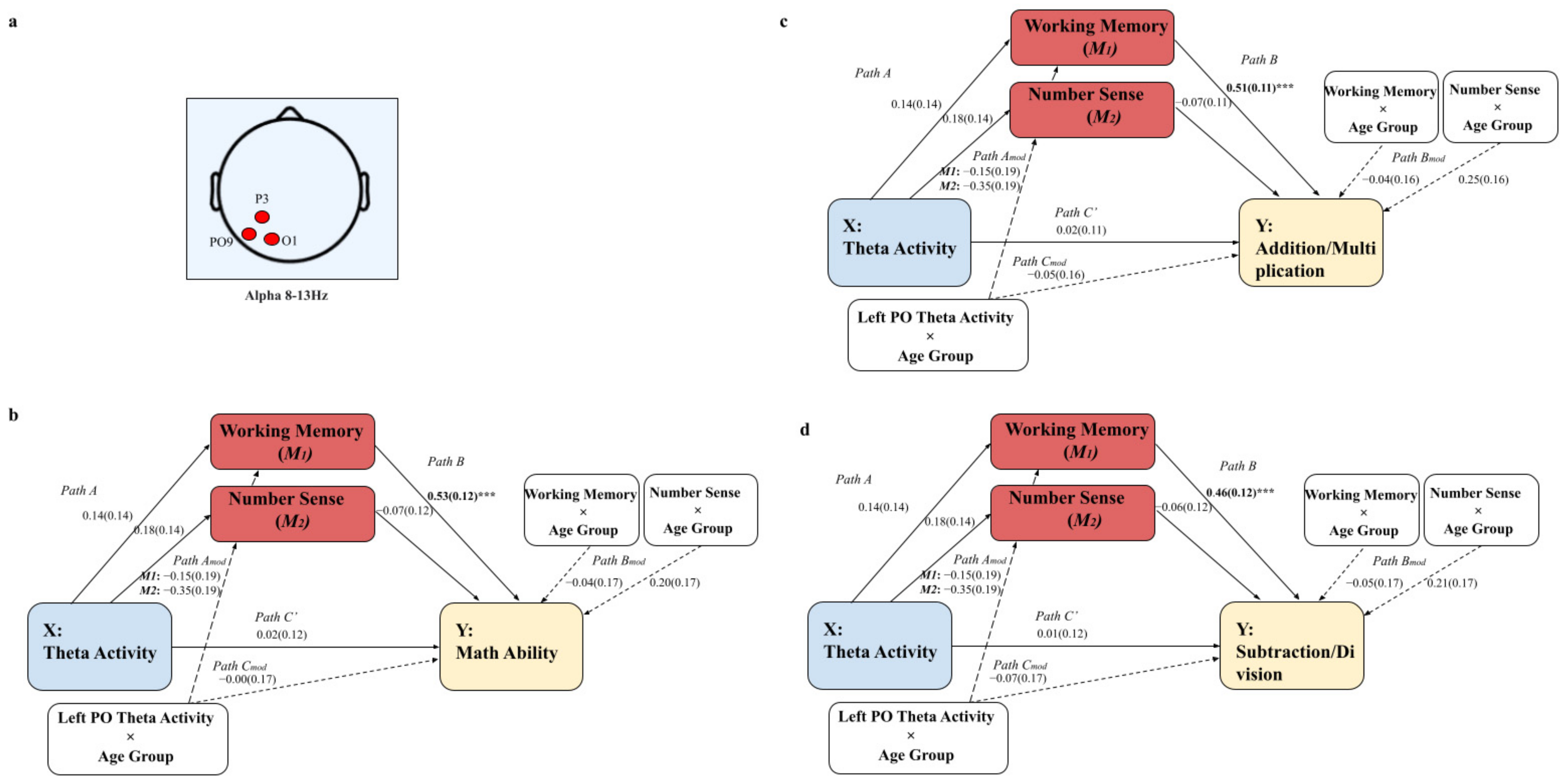
3.3. Exploratory Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Results of Resting-State Theta and Alpha Activity








References
- Butterworth, B.; Varma, S.; Laurillard, D. Dyscalculia: From brain to education. Science 2011, 332, 1049–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, S.; Bynner, B. Does Numeracy Matter More? National Research and Development Centre for Adult Literacy and Numeracy: London, UK, 2005. [Google Scholar]
- Cragg, L.; Gilmore, C. Skills underlying mathematics: The role of executive function in the development of mathematics proficiency. Trends Neurosci. Educ. 2014, 3, 63–68. [Google Scholar] [CrossRef]
- De Smedt, B.; Janssen, R.; Bouwens, K.; Verschaffel, L.; Boets, B.; Ghesquière, P. Working memory and individual differences in mathematics achievement: A longitudinal study from first grade to second grade. J. Exp. Child Psychol. 2009, 103, 186–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friso-Van Den Bos, I.; Van Der Ven, S.H.G.; Kroesbergen, E.H.; Van Luit, J.E.H. Working memory and mathematics in primary school children: A meta-analysis. Educ. Res. Rev. 2013, 10, 29–44. [Google Scholar] [CrossRef]
- Kroesbergen, E.H.; Van Luit, J.E.H.; Van Lieshout, E.C.D.M.; Van Loosbroek, E.; Van De Rijt, B.A.M. Individual differences in early numeracy: The role of executive functions and subitizing. J. Psychoeduc. Assess. 2009, 27, 226–236. [Google Scholar] [CrossRef]
- Kroesbergen, E.H.; van Dijk, M. Working memory and number sense as predictors of mathematical (Dis-)ability. Z. Psychol. 2015, 223, 102–109. [Google Scholar] [CrossRef]
- Raghubar, K.P.; Barnes, M.A.; Hecht, S.A. Working memory and mathematics: A review of developmental, individual difference, and cognitive approaches. Learn. Individ. Differ. 2010, 20, 110–122. [Google Scholar] [CrossRef]
- Sowinski, C.; LeFevre, J.-A.; Skwarchuk, S.-L.; Kamawar, D.; Bisanz, J.; Smith-Chant, B. Refining the quantitative pathway of the Pathways to Mathematics model. J. Exp. Child Psychol. 2015, 131, 73–93. [Google Scholar] [CrossRef]
- Geary, D.C.; vanMarle, K. Young children’s core symbolic and nonsymbolic quantitative knowledge in the prediction of later mathematics achievement. Dev. Psychol. 2016, 52, 2130–2144. [Google Scholar] [CrossRef]
- Sasanguie, D.; Göbel, S.M.; Moll, K.; Smets, K.; Reynvoet, B. Approximate number sense, symbolic number processing, or number–space mappings: What underlies mathematics achievement? J. Exp. Child Psychol. 2013, 114, 418–431. [Google Scholar] [CrossRef]
- Kail, R.; Hall, L.K. Distinguishing short-term memory from working memory. Mem. Cognit. 2001, 29, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mcintosh, A.; Reys, B.J.; Reys, R.E. A proposed framework for examining basic number sense. Learn. Math. 1992, 12, 2–44. [Google Scholar]
- Von Aster, M.G.; Shalev, R.S. Number development and developmental dyscalculia. Dev. Med. Child Neurol. 2007, 49, 868–873. [Google Scholar] [CrossRef]
- Best, J.R.; Miller, P.H.; Naglieri, J.A. Relations between executive function and academic achievement from ages 5 to 17 in a large, representative national sample. Learn. Individ. Differ. 2011, 21, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Friso-van den Bos, I.; Kroesbergen, E.H.; Van Luit, J.E.H. Number sense in kindergarten children: Factor structure and working memory predictors. Learn. Individ. Differ. 2014, 33, 23–29. [Google Scholar] [CrossRef]
- Kolkman, M.E.; Kroesbergen, E.H.; Leseman, P.P.M. Involvement of working memory in longitudinal development of number–magnitude skills. Infant Child Dev. 2014, 23, 36–50. [Google Scholar] [CrossRef]
- Nelwan, M.; Friso-van den Bos, I.; Vissers, C.; Kroesbergen, E. The relation between working memory, number sense, and mathematics throughout primary education in children with and without mathematical difficulties. Child Neuropsychol. 2021, 28, 143–170. [Google Scholar] [CrossRef]
- Rasmussen, C.; Bisanz, J. Representation and working memory in early arithmetic. J. Exp. Child Psychol. 2005, 91, 137–157. [Google Scholar] [CrossRef]
- Van de Weijer-Bergsma, E.; Kroesbergen, E.H.; Van Luit, J.E.H. Verbal and visual-spatial working memory and mathematical ability in different domains throughout primary school. Mem. Cognit. 2015, 43, 367. [Google Scholar] [CrossRef] [Green Version]
- Imbo, I.; Vandierendonck, A. The development of strategy use in elementary school children: Working memory and individual differences. J. Exp. Child Psychol. 2007, 96, 284–309. [Google Scholar] [CrossRef] [Green Version]
- Peters, L.; De Smedt, B. Arithmetic in the developing brain: A review of brain imaging studies. Dev. Cogn. Neurosci. 2018, 30, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Davis, N.; Cannistraci, C.J.; Rogers, B.P.; Gatenby, J.C.; Fuchs, L.S.; Anderson, A.W.; Gore, J.C. The neural correlates of calculation ability in children: An fMRI study. Magn. Reson. Imaging 2009, 27, 1187–1197. [Google Scholar] [CrossRef] [Green Version]
- Ischebeck, A.; Zamarian, L.; Egger, K.; Schocke, M.; Delazer, M. Imaging early practice effects in arithmetic. Neuroimage 2007, 36, 993–1003. [Google Scholar] [CrossRef]
- Ansari, D. Neurocognitive approaches to developmental disorders of numerical and mathematical cognition: The perils of neglecting the role of development. Learn. Individ. Differ. 2010, 20, 123–129. [Google Scholar] [CrossRef]
- Hinault, T.; Lemaire, P. What does EEG tell us about arithmetic strategies? A review. Int. J. Psychophysiol. 2016, 106, 115–126. [Google Scholar] [CrossRef]
- Artemenko, C.; Soltanlou, M.; Bieck, S.M.; Ehlis, A.-C.; Dresler, T.; Nuerk, H.-C. Individual differences in math ability determine neurocognitive processing of arithmetic complexity: A combined fNIRS-EEG study. Front. Hum. Neurosci. 2019, 13, 227. [Google Scholar] [CrossRef] [Green Version]
- De Smedt, B.; Grabner, R.H.; Studer, B. Oscillatory EEG correlates of arithmetic strategy use in addition and subtraction. Exp. Brain Res. 2009, 195, 635–642. [Google Scholar] [CrossRef]
- Grabner, R.H.; De Smedt, B. Neurophysiological evidence for the validity of verbal strategy reports in mental arithmetic. Biol. Psychol. 2011, 87, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Soltanlou, M.; Artemenko, C.; Dresler, T.; Fallgatter, A.J.; Nuerk, H.-C.; Ehlis, A.-C. Oscillatory EEG changes during arithmetic learning in children. Dev. Neuropsychol. 2019, 44, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Grabner, R.H.; De Smedt, B. Oscillatory EEG correlates of arithmetic strategies: A training study. Front. Psychol. 2012, 3, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, C.S.; Strüber, D.; Helfrich, R.F.; Engel, A.K. EEG oscillations: From correlation to causality. Int. J. Psychophysiol. 2016, 103, 12–21. [Google Scholar] [CrossRef]
- Ward, L.M. Synchronous neural oscillations and cognitive processes. Trends Cogn. Sci. 2003, 7, 553–559. [Google Scholar] [CrossRef]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Sauseng, P.; Griesmayr, B.; Freunberger, R. Control mechanisms in working memory: A possible function of EEG theta oscillations. Neurosci. Biobehav. Rev. 2010, 34, 1015–1022. [Google Scholar] [CrossRef]
- Van Bueren, N.E.R.; Reed, T.L.; Nguyen, V.; Sheffield, J.G.; van der Ven, S.H.G.; Osborne, M.A.; Kroesbergen, E.H.; Cohen Kadosh, R. Personalized brain stimulation for effective neurointervention across participants. PLoS Comput. Biol. 2021, 17, e1008886. [Google Scholar] [CrossRef]
- Donoghue, T.; Haller, M.; Peterson, E.J.; Varma, P.; Sebastian, P.; Gao, R.; Noto, T.; Lara, A.H.; Wallis, J.D.; Knight, R.T.; et al. Parameterizing neural power spectra into periodic and aperiodic components. Nat. Neurosci. 2020, 23, 1655–1665. [Google Scholar] [CrossRef]
- He, W.; Donoghue, T.; Sowman, P.F.; Seymour, R.A.; Brock, J.; Crain, S.; Voytek, B.; Hillebrand, A. Co-increasing neuronal noise and beta power in the developing brain. bioRxiv 2019, 839258. [Google Scholar] [CrossRef] [Green Version]
- Molina, J.L.; Voytek, B.; Thomas, M.L.; Joshi, Y.B.; Bhakta, S.G.; Talledo, J.A.; Swerdlow, N.R.; Light, G.A. Memantine effects on electroencephalographic measures of putative excitatory/inhibitory balance in schizophrenia. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 562–568. [Google Scholar] [CrossRef]
- Robertson, M.M.; Furlong, S.; Voytek, B.; Donoghue, T.; Boettiger, C.A.; Sheridan, M.A. EEG power spectral slope differs by ADHD status and stimulant medication exposure in early childhood. J. Neurophysiol. 2019, 122, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Voytek, B.; Kramer, M.A.; Case, J.; Lepage, K.Q.; Tempesta, Z.R.; Knight, R.T.; Gazzaley, A. Age-related changes in 1/f neural electrophysiological noise. J. Neurosci. 2015, 35, 13257–13265. [Google Scholar] [CrossRef]
- Miller, K.J.; Hermes, D.; Honey, C.J.; Hebb, A.O.; Ramsey, N.F.; Knight, R.T.; Ojemann, J.G.; Fetz, E.E. Human motor cortical activity is selectively phase-entrained on underlying rhythms. PLoS Comput. Biol. 2012, 8, e1002655. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Logothetis, N.; Singer, W. Scaling brain size, keeping timing: Evolutionary preservation of brain rhythms. Neuron 2013, 80, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Peterson, E.J.; Voytek, B. Inferring synaptic excitation/inhibition balance from field potentials. Neuroimage 2017, 158, 70–78. [Google Scholar] [CrossRef]
- Cellier, D.; Riddle, J.; Petersen, I.; Hwang, K. The development of theta and alpha neural oscillations from ages 3 to 24 years. Dev. Cogn. Neurosci. 2021, 50, 100969. [Google Scholar] [CrossRef]
- Halberda, J.; Feigenson, L. Developmental change in the acuity of the “number sense”: The approximate number system in 3-, 4-, 5-, and 6-year-olds and adults. Dev. Psychol. 2008, 44, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Cragg, L.; Keeble, S.; Richardson, S.; Roome, H.E.; Gilmore, C. Direct and indirect influences of executive functions on mathematics achievement. Cognition 2017, 162, 12–26. [Google Scholar] [CrossRef] [Green Version]
- De Vos, T. TTR. Tempotest Rekenen [Arithmetic Number Fact Test]; Swets & Zeitlinger: Lisse, The Netherlands, 1992. [Google Scholar]
- Friso-Van den Bos, I.; Schoevers, E.M.; Slot, E.; Kroesbergen, E.H. The Dutch Assessment of Number Sense (DANS): Analyses of the Conditions of the Number Line, Symbolic Comparison, and Non-Symbolic Comparison Task; Department of Education and Pedagogy, Utrecht University: Utrecht, The Netherlands, 2015. [Google Scholar]
- Huijsmans, M.D.E.; Kleemans, T.; Kroesbergen, E.H. How cognitive strengths compensate weaknesses related to specific learning difficulties in fourth-grade children. Front. Psychol. 2021, 12, 552458. [Google Scholar] [CrossRef]
- Alloway, T.P.; Gathercole, S.E.; Kirkwood, H.; Elliott, J. Evaluating the validity of the automated working memory assessment. Educ. Psychol. 2008, 28, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Zacharopoulos, G.; Sella, F.; Kadosh, K.C.; Hartwright, C.; Emir, U.; Cohen Kadosh, R. Predicting learning and achievement using GABA and glutamate concentrations in human development. PLoS Biol. 2021, 19, e3001325. [Google Scholar] [CrossRef]
- Roopun, A.K.; Middleton, S.J.; Cunningham, M.O.; LeBeau, F.E.H.; Bibbig, A.; Whittington, M.A.; Traub, R.D. A beta2-frequency (20–30 Hz) oscillation in nonsynaptic networks of somatosensory cortex. Proc. Natl. Acad. Sci. USA 2006, 103, 15646–15650. [Google Scholar] [CrossRef] [Green Version]
- Yamawaki, N.; Stanford, I.M.; Hall, S.D.; Woodhall, G.L. Pharmacologically induced and stimulus evoked rhythmic neuronal oscillatory activity in the primary motor cortex in vitro. Neuroscience 2008, 151, 386–395. [Google Scholar] [CrossRef]
- Frohlich, J.; Senturk, D.; Saravanapandian, V.; Golshani, P.; Reiter, L.T.; Sankar, R.; Thibert, R.L.; DiStefano, C.; Huberty, S.; Cook, E.H.; et al. A quantitative electrophysiological biomarker of duplication 15q11.2-q13.1 syndrome. PLoS ONE 2016, 11, e0167179. [Google Scholar] [CrossRef] [Green Version]
- Frohlich, J.; Miller, M.T.; Bird, L.M.; Garces, P.; Purtell, H.; Hoener, M.C.; Philpot, B.D.; Sidorov, M.S.; Tan, W.H.; Hernandez, M.C.; et al. Electrophysiological phenotype in angelman syndrome differs between genotypes. Biol. Psychiatry 2019, 85, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Van Bueren, N.E.R.; Van der Ven, H.G.; Kroesbergen, E.H.; Sella, F.; Cohen Kadosh, R. Explaining and predicting the effects of neurostimulation via neuronal excitation/inhibition in the brain. bioRxiv 2022. to be submitted. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Children | Adults | Mean Comparison | ||||
|---|---|---|---|---|---|---|
| Variables | Mean | SD | Mean | SD | t-Value | p |
| TTR Total | 100.84 | 28.84 | 147.83 | 25.33 | −8.86 | <0.001 |
| Error score NLE | 44.73 | 22.76 | 28.22 | 10.54 | 4.69 | <0.001 |
| RTs Symbolic | 1125.09 | 650.88 | 648.63 | 82.24 | 5.13 | <0.001 |
| Accuracy Non−Symbolic | 29.77 | 3.41 | 30.35 | 2.90 | −0.93 | 0.35 |
| Accuracy BDR | 10.78 | 3.33 | 14.76 | 4.09 | −5.52 | <0.001 |
| Accuracy OOO | 15.46 | 3.69 | 20.41 | 2.98 | −7.46 | <0.001 |
| Number Sense (combined) | 1617.72 | 219.22 | 1784.49 | 28.72 | −5.28 | <0.001 |
| Working Memory (combined) | 13.13 | 2.86 | 17.58 | 2.81 | −8.02 | <0.001 |
| Beta parameterization | 0.27 | 0.17 | 0.26 | 0.16 | −0.11 | 0.90 |
| Beta traditional | 0.39 | 0.17 | 0.46 | 0.15 | −2.13 | 0.035 |
| Aperiodic activity 1 (13–40 Hz) | 1.33 | 1.20 | 0.69 | 1.18 | 2.79 | p < 0.001 |
| Aperiodic activity 1 (1–40 Hz) | 0.90 | 0.29 | 0.47 | 0.28 | 7.64 | <0.001 |
| Offset | 0.37 | 0.29 | −0.23 | 0.30 | 10.67 | <0.001 |
| Exponent | 1.43 | 0.31 | 1.19 | 0.29 | 4.07 | <0.001 |
| Beta Parameterization | Beta Traditional | |
|---|---|---|
| Beta Parameterization | - | |
| Beta Traditional | 0.05 | - |
| Aperiodic Activity | 0.35 *** | −0.54 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Bueren, N.E.R.; van der Ven, S.H.G.; Roelofs, K.; Cohen Kadosh, R.; Kroesbergen, E.H. Predicting Math Ability Using Working Memory, Number Sense, and Neurophysiology in Children and Adults. Brain Sci. 2022, 12, 550. https://doi.org/10.3390/brainsci12050550
van Bueren NER, van der Ven SHG, Roelofs K, Cohen Kadosh R, Kroesbergen EH. Predicting Math Ability Using Working Memory, Number Sense, and Neurophysiology in Children and Adults. Brain Sciences. 2022; 12(5):550. https://doi.org/10.3390/brainsci12050550
Chicago/Turabian Stylevan Bueren, Nienke E. R., Sanne H. G. van der Ven, Karin Roelofs, Roi Cohen Kadosh, and Evelyn H. Kroesbergen. 2022. "Predicting Math Ability Using Working Memory, Number Sense, and Neurophysiology in Children and Adults" Brain Sciences 12, no. 5: 550. https://doi.org/10.3390/brainsci12050550
APA Stylevan Bueren, N. E. R., van der Ven, S. H. G., Roelofs, K., Cohen Kadosh, R., & Kroesbergen, E. H. (2022). Predicting Math Ability Using Working Memory, Number Sense, and Neurophysiology in Children and Adults. Brain Sciences, 12(5), 550. https://doi.org/10.3390/brainsci12050550