
Micro-RNAs from Plasma-Derived Small Extracellular Vesicles as Potential Biomarkers for Tic Disorders Diagnosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants’ Clinical Characteristics
2.2. Blood Collection and Isolation of Small Extracellular Vesicles (sEV)
2.3. Nanoparticle Tracking Analysis (NTA)
2.4. Transmission Electron Microscopy (TEM)
2.5. Western Blot Assay
2.6. Library Preparation and Sequencing
2.7. Bioinformatics Analysis of miRNA
2.8. Gene Ontology (GO) Annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
2.9. Statistical Analysis
3. Results
3.1. Clinical Features of Participants
3.2. Isolation and Characterization of Human Plasma-Derived sEVs
3.3. Identification of Differentially Expressed miRNAs in Plasma sEVs from the Tic Disorder Group and HCs
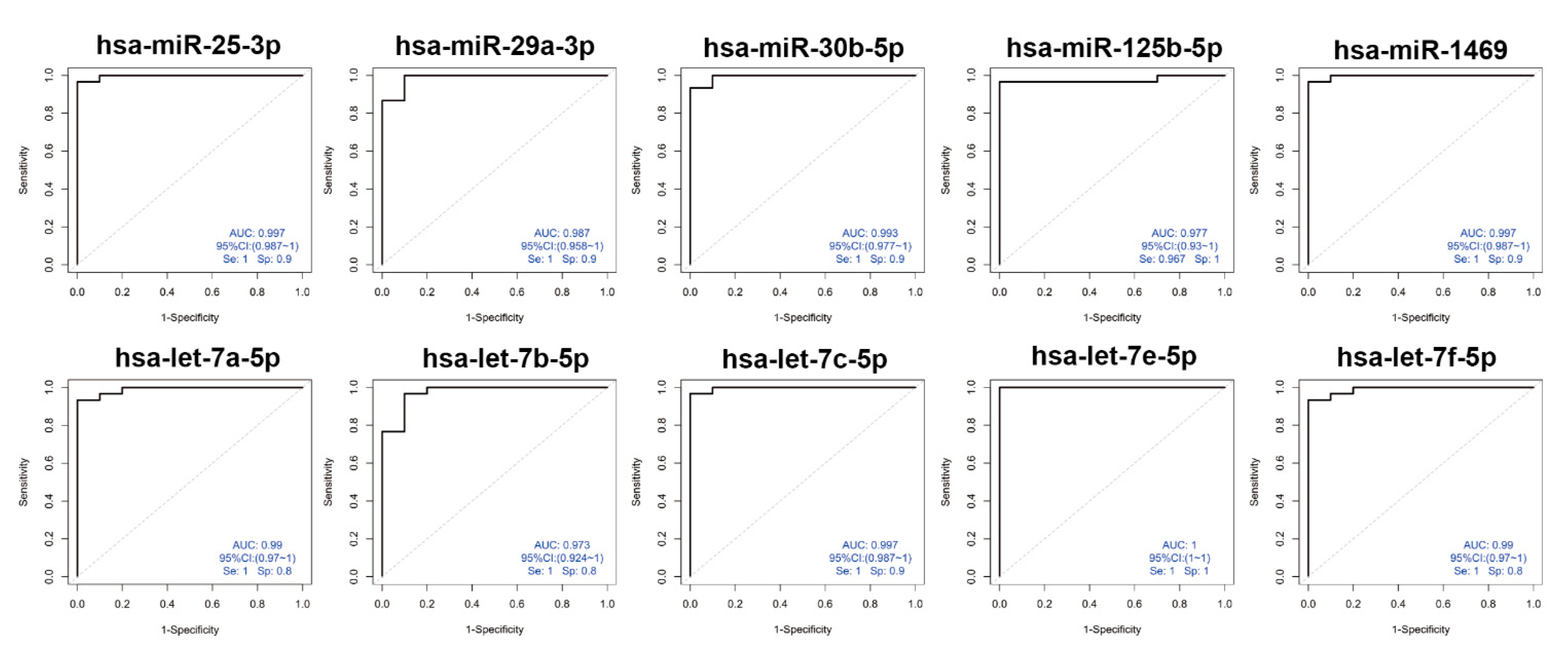
3.4. Diagnostic Potential for sEVs-Derived miRNAs by ROC Curve Analysis
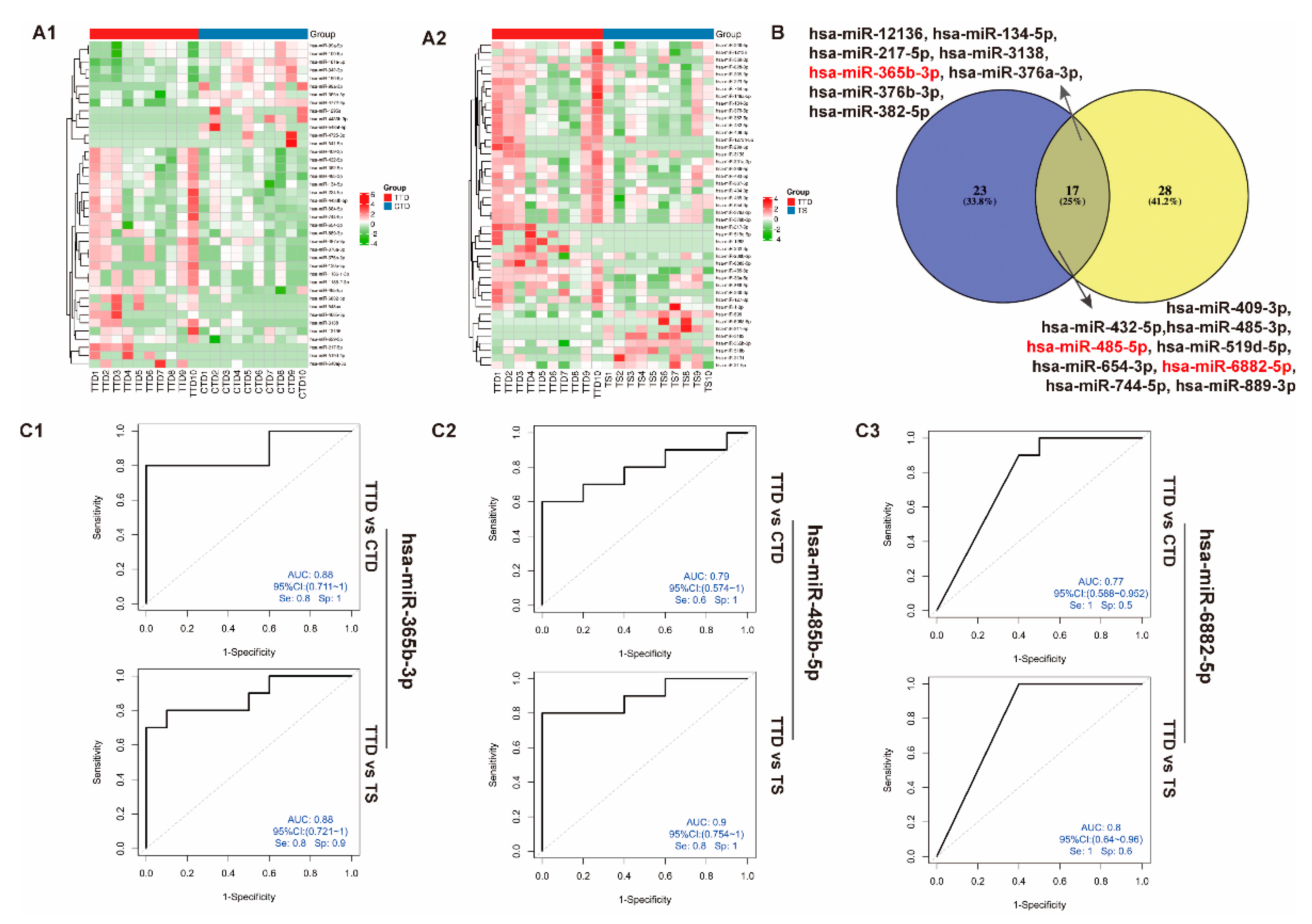
3.5. Identification of Differentially Expressed miRNAs in Plasma sEVs from the TTD, CTD, and TS Groups
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ganos, C. Tics and Tourette’s: Update on pathophysiology and tic control. Curr. Opin. Neurol. 2016, 29, 513–518. [Google Scholar] [CrossRef] [PubMed]
- First, M.B. Diagnostic and statistical manual of mental disorders, 5th edition, and clinical utility. J. Nerv. Ment. Dis. 2013, 201, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Delgar, B.; Servera, M.; Coffey, B.J.; Lazaro, L.; Openneer, T.; Benaroya-Milshtein, N.; Steinberg, T.; Hoekstra, P.J.; Dietrich, A.; Morer, A.; et al. Tic disorders in children and adolescents: Does the clinical presentation differ in males and females? A report by the EMTICS group. Eur. Child Adolesc. Psychiatry 2021, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cui, Y.; Li, Y.; Guo, L.; Ke, X.; Liu, J.; Luo, X.; Zheng, Y.; Leckman, J.F. Prevalence of mental disorders in school children and adolescents in China: Diagnostic data from detailed clinical assessments of 17,524 individuals. J. Child Psychol. Psychiatry 2022, 63, 34–46. [Google Scholar] [CrossRef]
- Billnitzer, A.; Jankovic, J. Current Management of Tics and Tourette Syndrome: Behavioral, Pharmacologic, and Surgical Treatments. Neurotherapeutics 2020, 17, 1681–1693. [Google Scholar] [CrossRef]
- Cavanna, A.E.; David, K.; Orth, M.; Robertson, M.M. Predictors during childhood of future health-related quality of life in adults with Gilles de la Tourette Syndrome. Eur. J. Paediatr. Neurol. 2012, 16, 605–612. [Google Scholar] [CrossRef]
- Evans, J.; Seri, S.; Cavanna, A.E. The effects of Gilles de la Tourette Syndrome and other chronic tic disorders on quality of life across the lifespan: A systematic review. Eur. Child Adolesc. Psychiatry 2016, 25, 939–948. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, R.; Ragusa, M.; Barbagallo, C.; Sammito, M.; Gulisano, M.; Cali, P.V.; Pappalardo, C.; Barchitta, M.; Granata, M.; Condorelli, A.G.; et al. Circulating miRNAs profiles in Tourette Syndrome: Molecular data and clinical implications. Mol. Brain 2015, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Wilens, T.E.; Spencer, T.J. Understanding attention-deficit/hyperactivity disorder from childhood to adulthood. Postgrad Med. 2010, 122, 97–109. [Google Scholar] [CrossRef]
- Randall, M.; Egberts, K.J.; Samtani, A.; Scholten, R.J.; Hooft, L.; Livingstone, N.; Sterling-Levis, K.; Woolfenden, S.; Williams, K. Diagnostic tests for autism spectrum disorder (ASD) in preschool children. Cochrane Database Syst. Rev. 2018, 7, CD009044. [Google Scholar] [CrossRef] [Green Version]
- Novotny, M.; Valis, M.; Klimova, B. Tourette Syndrome: A Mini-Review. Front. Neurol. 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Reyes, P.R.; Garza, B.S.; Sharma, A. MicroRNAs and Child Neuropsychiatric Disorders: A Brief Review. Neurochem. Res. 2020, 45, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Calin, G.A.; Liu, C.G.; Sevignani, C.; Ferracin, M.; Felli, N.; Dumitru, C.D.; Shimizu, M.; Cimmino, A.; Zupo, S.; Dono, M.; et al. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc. Natl. Acad. Sci. USA 2004, 101, 11755–11760. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Visone, R.; Di Leva, G.; Donati, V.; Petrocca, F.; Casalini, P.; Taccioli, C.; Volinia, S.; Liu, C.G.; Alder, H.; et al. MicroRNA signatures in human ovarian cancer. Cancer Res. 2007, 67, 8699–8707. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Yeo, G.; Muotri, A.R.; Kuwabara, T.; Gage, F.H. Noncoding RNAs in the mammalian central nervous system. Annu. Rev. Neurosci. 2006, 29, 77–103. [Google Scholar] [CrossRef] [Green Version]
- Bavamian, S.; Mellios, N.; Lalonde, J.; Fass, D.M.; Wang, J.; Sheridan, S.D.; Madison, J.M.; Zhou, F.; Rueckert, E.H.; Barker, D.; et al. Dysregulation of miR-34a links neuronal development to genetic risk factors for bipolar disorder. Mol. Psychiatry 2015, 20, 573–584. [Google Scholar] [CrossRef]
- Mehler, M.F.; Mattick, J.S. Noncoding RNAs and RNA editing in brain development, functional diversification, and neurological disease. Physiol. Rev. 2007, 87, 799–823. [Google Scholar] [CrossRef] [Green Version]
- Femminella, G.D.; Ferrara, N.; Rengo, G. The emerging role of microRNAs in Alzheimer’s disease. Front. Physiol. 2015, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margis, R.; Margis, R.; Rieder, C.R. Identification of blood microRNAs associated to Parkinsonis disease. J. Biotechnol. 2011, 152, 96–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.M.; Li, X.M.; Zheng, X.P.; Yu, J.T.; Tan, L. MicroRNAs dysregulation in epilepsy. Brain Res. 2014, 1584, 94–104. [Google Scholar] [CrossRef]
- Wang, Y.L.; Wang, Y.P.; Chen, Y.; Hua, Y.; Xu, L.; Zhu, M.Y.; Zhao, C.Y.; Zhang, W.R.; Sheng, G.X.; Liu, L.; et al. Circulating MicroRNAs From Plasma Small Extracellular Vesicles as Potential Diagnostic Biomarkers in Pediatric Epilepsy and Drug-Resistant Epilepsy. Front. Mol. Neurosci. 2022, 15, 823802. [Google Scholar] [CrossRef]
- Abelson, J.F.; Kwan, K.Y.; O’Roak, B.J.; Baek, D.Y.; Stillman, A.A.; Morgan, T.M.; Mathews, C.A.; Pauls, D.L.; Rasin, M.R.; Gunel, M.; et al. Sequence variants in SLITRK1 are associated with Tourette’s syndrome. Science 2005, 310, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating Extracellular Vesicles in Human Disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 147. [Google Scholar] [CrossRef]
- Gamez-Valero, A.; Monguio-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.; Beyer, K.; Borras, F.E. Size-Exclusion Chromatography-based isolation minimally alters Extracellular Vesicles’ characteristics compared to precipitating agents. Sci. Rep. 2016, 6, 33641. [Google Scholar] [CrossRef] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Monguio-Tortajada, M.; Galvez-Monton, C.; Bayes-Genis, A.; Roura, S.; Borras, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef] [PubMed]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Coleman, B.M.; Hanssen, E.; Lawson, V.A.; Hill, A.F. Prion-infected cells regulate the release of exosomes with distinct ultrastructural features. FASEB J. 2012, 26, 4160–4173. [Google Scholar] [CrossRef]
- Coumans, F.A.W.; Gool, E.L.; Nieuwland, R. Bulk immunoassays for analysis of extracellular vesicles. Platelets 2017, 28, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, A.; Fredsoe, J.; Sorensen, K.D.; Borre, M.; Evander, M.; Laurell, T.; Lilja, H.; Ceder, Y. High-Throughput and Automated Acoustic Trapping of Extracellular Vesicles to Identify microRNAs With Diagnostic Potential for Prostate Cancer. Front. Oncol. 2021, 11, 631021. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.S.; Cui, Y.H.; Sun, D.; Lu, Q.; Jiang, Y.W.; Jiang, L.; Wang, J.Q.; Luo, R.; Fang, F.; Zhou, S.Z.; et al. Current Status, Diagnosis, and Treatment Recommendation for Tic Disorders in China. Front. Psychiatry 2020, 11, 774. [Google Scholar] [CrossRef]
- Wei, Z.; Batagov, A.O.; Schinelli, S.; Wang, J.; Wang, Y.; El Fatimy, R.; Rabinovsky, R.; Balaj, L.; Chen, C.C.; Hochberg, F.; et al. Coding and noncoding landscape of extracellular RNA released by human glioma stem cells. Nat. Commun. 2017, 8, 1145. [Google Scholar] [CrossRef] [Green Version]
- Min, L.; Zhu, S.; Chen, L.; Liu, X.; Wei, R.; Zhao, L.; Yang, Y.; Zhang, Z.; Kong, G.; Li, P.; et al. Evaluation of circulating small extracellular vesicles derived miRNAs as biomarkers of early colon cancer: A comparison with plasma total miRNAs. J. Extracell Vesicles 2019, 8, 1643670. [Google Scholar] [CrossRef] [Green Version]
- Qing, L.; Chen, H.; Tang, J.; Jia, X. Exosomes and Their MicroRNA Cargo: New Players in Peripheral Nerve Regeneration. Neurorehabil. Neural. Repair. 2018, 32, 765–776. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, M.M.J.; Krauskopf, J.; Ramaekers, J.G.; Kleinjans, J.C.S.; Prickaerts, J.; Briede, J.J. Circulating microRNAs as potential biomarkers for psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2020, 185, 101732. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Baixauli, F.; Gutierrez-Vazquez, C.; Sanchez-Madrid, F.; Mittelbrunn, M. Sorting it out: Regulation of exosome loading. Semin Cancer Biol. 2014, 28, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.P.; Lin, Z.X.; Jiang, X.Y.; Yu, X.Y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharmacol. Sin. 2018, 39, 542–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thery, C. Cancer: Diagnosis by extracellular vesicles. Nature 2015, 523, 161–162. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Gao, K.P.; Wang, Y.X.; Liu, Z.C.; Tian, L.; Yang, X.Z.; Ding, J.Y.; Wu, W.T.; Yang, W.H.; Li, Y.L.; et al. RNA sequencing identified specific circulating miRNA biomarkers for early detection of diabetes retinopathy. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E374–E385. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Lin, Z.; Peng, W.; Hu, Z. Identification of potential biomarkers of lung adenocarcinoma brain metastases via microarray analysis of cDNA expression profiles. Oncol. Lett. 2019, 17, 2228–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Higgins, H.; Guo, J.; Harrison, K.; Schultz, E.N.; Hales, B.J.; Moses, E.K.; Goldblatt, J.; Pachter, N.; Zhang, G.C. Clinical significance of circulating microRNAs as markers in detecting and predicting congenital heart defects in children. J. Transl. Med. 2018, 16, 42. [Google Scholar] [CrossRef]
- Mahmoudian, M.; Razmara, E.; Hussen, B.M.; Simiyari, M.; Lotfizadeh, N.; Motaghed, H.; Monfared, A.K.; Montazeri, M.; Babashah, S. Identification of a six-microRNA signature as a potential diagnostic biomarker in breast cancer tissues. J. Clin. Lab. Anal. 2021, 35, e24010. [Google Scholar] [CrossRef]
- Cao, D.P.; Jiang, B.F.; Zhang, Y.G.; Pang, M.G. microRNA-125b-5p is a promising novel plasma biomarker for alveolar echinococcosis in patients from the southern province of Qinghai. BMC Infect. Dis. 2021, 21, 246. [Google Scholar] [CrossRef]
- Sun, N.; Nasello, C.; Deng, L.; Wang, N.; Zhang, Y.; Xu, Z.; Song, Z.; Kwan, K.; King, R.A.; Pang, Z.P.; et al. The PNKD gene is associated with Tourette Disorder or Tic disorder in a multiplex family. Mol. Psychiatry 2018, 23, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.Y.; Wu, M. mRNA expression of dopamine receptor D2 and dopamine transporter in peripheral blood lymphocytes before and after treatment in children with tic disorder. Zhongguo Dang Dai Er Ke Za Zhi 2016, 18, 297–300. [Google Scholar]
- Moya, P.R.; Wendland, J.R.; Rubenstein, L.M.; Timpano, K.R.; Heiman, G.A.; Tischfield, J.A.; King, R.A.; Andrews, A.M.; Ramamoorthy, S.; McMahon, F.J.; et al. Common and rare alleles of the serotonin transporter gene, SLC6A4, associated with Tourette’s disorder. Mov. Disord. 2013, 28, 1263–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos-Veneman, N.G.; Minderaa, R.B.; Hoekstra, P.J. The DRD4 gene and severity of tics and comorbid symptoms: Main effects and interactions with delivery complications. Mov. Disord. 2010, 25, 1470–1476. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery Set | ||||
|---|---|---|---|---|
| TD | HCs | |||
| TS | TTD | CTD | ||
| NO | 10 | 10 | 10 | 10 |
| Female: Male | 2:8 | 3:7 | 0:10 | 3:7 |
| Age, Mean ± SD | 9.01 ± 2.84 | 6.76 ± 1.61 | 9.42 ± 1.78 | 9.52 ± 2.86 |
| Age at first attack, Mean ± SD | 6.06 ± 1.57 | 6.50 ± 1.41 | 7.71 ± 1.98 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xu, X.; Chen, H.; Zhu, M.; Guo, X.; Gao, F. Micro-RNAs from Plasma-Derived Small Extracellular Vesicles as Potential Biomarkers for Tic Disorders Diagnosis. Brain Sci. 2022, 12, 829. https://doi.org/10.3390/brainsci12070829
Wang Y, Xu X, Chen H, Zhu M, Guo X, Gao F. Micro-RNAs from Plasma-Derived Small Extracellular Vesicles as Potential Biomarkers for Tic Disorders Diagnosis. Brain Sciences. 2022; 12(7):829. https://doi.org/10.3390/brainsci12070829
Chicago/Turabian StyleWang, Yilong, Xuebin Xu, Haihua Chen, Mengying Zhu, Xiaotong Guo, and Feng Gao. 2022. "Micro-RNAs from Plasma-Derived Small Extracellular Vesicles as Potential Biomarkers for Tic Disorders Diagnosis" Brain Sciences 12, no. 7: 829. https://doi.org/10.3390/brainsci12070829
APA StyleWang, Y., Xu, X., Chen, H., Zhu, M., Guo, X., & Gao, F. (2022). Micro-RNAs from Plasma-Derived Small Extracellular Vesicles as Potential Biomarkers for Tic Disorders Diagnosis. Brain Sciences, 12(7), 829. https://doi.org/10.3390/brainsci12070829