
Sleep Fosters Odor Recognition in Children with Attention Deficit Hyperactivity Disorder but Not in Typically Developing Children

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design and Procedures
2.3. Statistical Analysis
3. Results
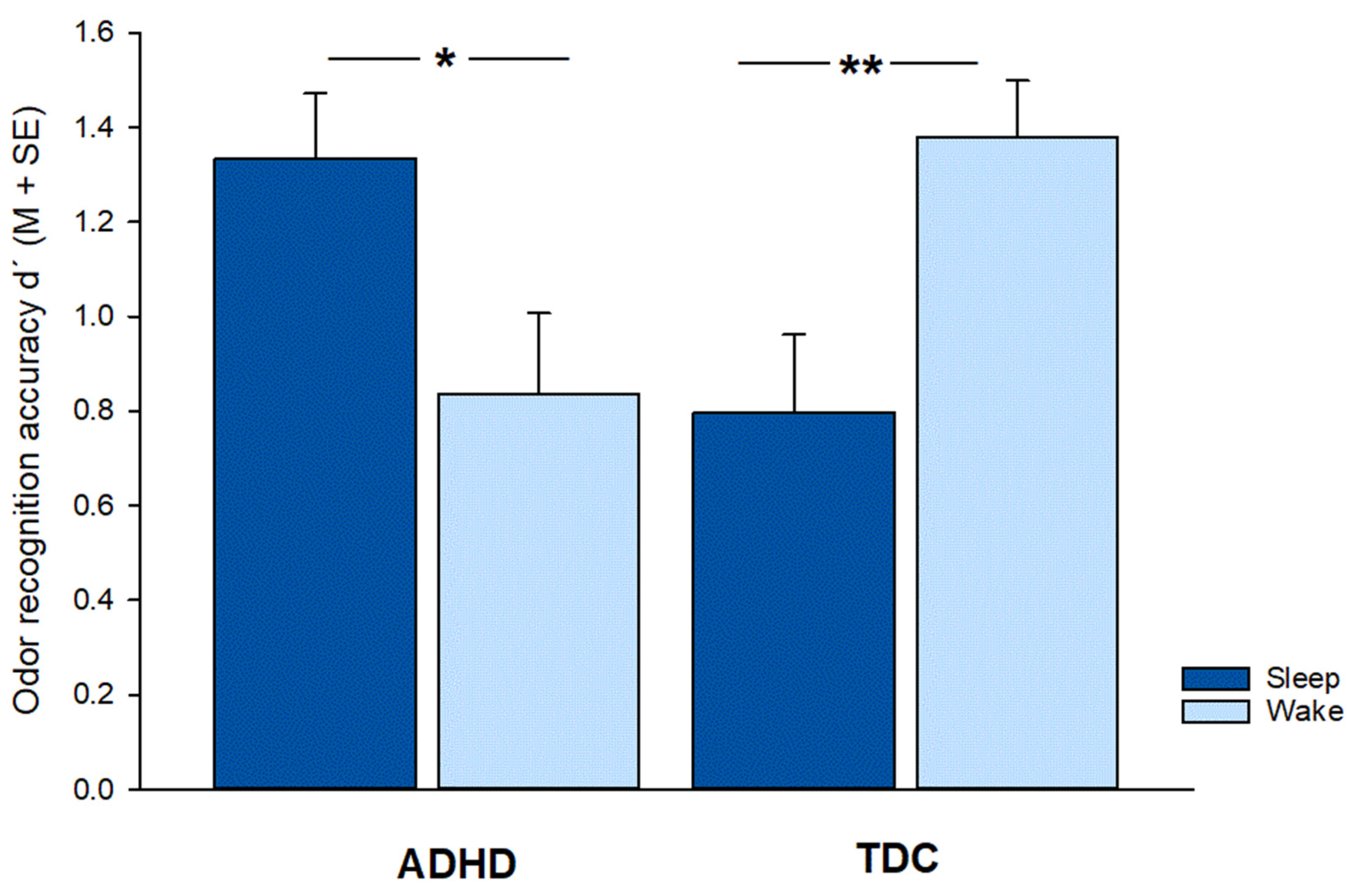
3.1. Memory Performance
3.2. Odor Rating
3.2.1. Encoding
3.2.2. Retrieval
3.3. Manipulation Check
3.4. Control Variables
4. Discussion
4.1. Superior Consolidation of Odor Memory in ADHD Because of Abundant Pre-Experience?
4.2. Lowered Threshold for Olfaction a Useful Biomarker in ADHD?
4.3. Strengths and Limitations
4.4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rasch, B.; Born, J. About Sleep’s Role in Memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef] [PubMed]
- Rasch, B.; Büchel, C.; Gais, S.; Born, J. Odor Cues During Slow-Wave Sleep Prompt Declarative Memory Consolidation. Science 2007, 315, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, I.; Diekelmann, S.; Born, J. Sleep in children improves memory performance on declarative but not procedural tasks. Learn. Mem. 2008, 15, 373–377. [Google Scholar] [CrossRef]
- Maski, K.; Holbrook, H.; Manoach, D.; Hanson, E.; Kapur, K.; Stickgold, R. Sleep Dependent Memory Consolidation in Children with Autism Spectrum Disorder. Sleep 2015, 38, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, I.; Prehn-Kristensen, A.; Born, J. Sleep-dependent memory consolidation–what can be learnt from children? Neurosci. Biobehav. Rev. 2012, 36, 1718. [Google Scholar] [CrossRef]
- Larsson, M.; Willander, J. Autobiographical odor memory. Ann. N. Y. Acad. Sci. 2009, 1170, 318–323. [Google Scholar] [CrossRef]
- Herz, R.S. Odor memory and the special role of associative learning. In Olfactory Cognition: From Perception and Memory to Environmental Odours and Neuroscience; Zucco, G.M., Herz, R.S., Schaal, B., Eds.; John Benjamins Publishing Company: Amsterdam, The Netherlands, 2012; pp. 95–114. [Google Scholar]
- Nguyen, L.A.; Ober, B.A.; Shenaut, G.K. Odor recognition memory: Two encoding trials are better than one. Chem. Senses 2012, 37, 745–754. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Lotzkat, K.; Bauhofer, E.; Wiesner, C.D.; Baving, L. Sleep Supports Memory of Odors in Adults but Not in Children. PLoS ONE 2015, 10, e0139069. [Google Scholar] [CrossRef]
- Thomas, R.; Sanders, S.; Doust, J.; Beller, E.; Glasziou, P. Prevalence of attention-deficit/hyperactivity disorder: A systematic review and meta-analysis. Pediatrics 2015, 135, e994–e1001. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Faraone, S.V.; Larsson, H. Genetics of attention deficit hyperactivity disorder. Mol. Psychiatry 2019, 24, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Saccaro, L.F.; Schilliger, Z.; Perroud, N.; Piguet, C. Inflammation, Anxiety, and Stress in Attention-Deficit/Hyperactivity Disorder. Biomedicines 2021, 9, 1313. [Google Scholar] [CrossRef]
- Shaw, P.; Eckstrand, K.; Sharp, W.; Blumenthal, J.; Lerch, J.P.; Greenstein, D.; Clasen, L.; Evans, A.; Giedd, J.; Rapoport, J.L. Attention-deficit/hyperactivity disorder is characterized by a delay in cortical maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 19649–19654. [Google Scholar] [CrossRef] [PubMed]
- Rubia, K. Neuro-anatomic evidence for the maturational delay hypothesis of ADHD. Proc. Natl. Acad. Sci. USA 2007, 104, 19663–19664. [Google Scholar] [CrossRef]
- Rubia, K.; Alegria, A.; Brinson, H. Imaging the ADHD brain: Disorder-specificity, medication effects and clinical translation. Expert Rev. Neurother. 2014, 14, 519–538. [Google Scholar] [CrossRef]
- Rubia, K. Cognitive Neuroscience of Attention Deficit Hyperactivity Disorder (ADHD) and Its Clinical Translation. Front. Hum. Neurosci. 2018, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Gorgoni, M.; D’Atri, A.; Scarpelli, S.; Reda, F.; De Gennaro, L. Sleep electroencephalography and brain maturation: Developmental trajectories and the relation with cognitive functioning. Sleep Med. 2020, 66, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, K.; Labrosse, M.; Gingras, M.A.; Godbout, R. Sleep Instability Correlates with Attentional Impairment in Boys with Attention Deficit Hyperactivity Disorder. Brain Sci. 2021, 11, 1425. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Göder, R.; Fischer, J.; Wilhelm, I.; Seeck-Hirschner, M.; Aldenhoff, J.; Baving, L. Reduced sleep-associated consolidation of declarative memory in attention-deficit/hyperactivity disorder. Sleep Med. 2011, 12, 672–679. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Munz, M.; Molzow, I.; Wilhelm, I.; Wiesner, C.D.; Baving, L. Sleep promotes consolidation of emotional memory in healthy children but not in children with attention-deficit hyperactivity disorder. PLoS ONE 2013, 29, e65098. [Google Scholar] [CrossRef]
- Cremone, A.; Lugo-Candelas, C.I.; Harvey, E.A.; McDermott, J.M.; Spencer, R.M. REM theta activity enhances inhibitory control in typically developing children but not children with ADHD symptoms. Exp. Brain. Res. 2017, 235, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Sonuga-Barke, E.; Bitsakou, P.; Thompson, M. Beyond the dual pathway model: Evidence for the dissociation of timing, inhibitory, and delay-related impairments in attention-deficit/hyperactivity disorder. J. Am. Acad. Child. Adolesc. Psychiatry. 2010, 49, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, R.; Brem, S.; Brandeis, D.; Grünblatt, E.; Berger, G.; Walitza, S. ADHD: Current Concepts and Treatments in Children and Adolescents. Neuropediatrics 2020, 51, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Wilens, T.E. Effects of methylphenidate on the catecholaminergic system in attention-deficit/hyperactivity disorder. J. Clin. Psychopharmacol. 2008, 28, 46–53. [Google Scholar] [CrossRef]
- Rubia, K.; Alegria, A.A.; Cubillo, A.I.; Smith, A.B.; Brammer, M.J.; Radua, J. Effects of stimulants on brain function in attention-deficit/hyperactivity disorder: A systematic review and meta-analysis. Biol. Psychiatry 2014, 76, 616–628. [Google Scholar] [CrossRef]
- Del Campo, N.; Chamberlain, S.R.; Sahakian, B.J.; Robbins, T.W. The roles of dopamine and noradrenaline in the pathophysiology and treatment of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2011, 69, e145–e157. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobio. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef]
- Schecklmann, M.; Schaldecker, M.; Aucktor, S.; Brast, J.; Kirchgassner, K.; Muhlberger, A.; Warnke, A.; Gerlach, M.; Fallgatter, A.J.; Romanos, M. Effects of methylphenidate on olfaction and frontal and temporal brain oxygenation in children with ADHD. J. Psychiatr. Res. 2011, 45, 1463–1470. [Google Scholar] [CrossRef]
- Lorenzen, A.; Scholz-Hehn, D.; Wiesner, C.D.; Wolff, S.; Bergmann, T.O.; van Eimeren, T.; Lentfer, L.; Baving, L.; Prehn-Kristensen, A. Chemosensory processing in children with attention-deficit/hyperactivity disorder. J. Psychiatr. Res. 2016, 76, 121–127. [Google Scholar] [CrossRef]
- Romanos, M.; Renner, T.J.; Schecklmann, M.; Hummel, B.; Roos, M.; von Mering, C.; Pauli, P.; Reichmann, H.; Warnke, A.; Gerlach, M. Improved odor sensitivity in attention-deficit/hyperactivity disorder. Biol. Psychiatry 2008, 64, 938–940. [Google Scholar] [CrossRef]
- Barkley, R.A. Behavioral inhibition, sustained attention, and executive functions: Constructing a unifying theory of ADHD. Psychol. Bull. 1997, 121, 65–94. [Google Scholar] [CrossRef] [PubMed]
- Willcutt, E.G.; Doyle, A.E.; Nigg, J.T.; Faraone, S.V.; Pennington, B.F. Validity of the executive function theory of attention-deficit/hyperactivity disorder: A meta-analytic review. Biol. Psychiatry. 2005, 57, 1336–1346. [Google Scholar] [CrossRef] [PubMed]
- Prehn-Kristensen, A.; Munz, M.; Göder, R.; Wilhelm, I.; Korr, K.; Vahl, W.; Wiesner, C.D.; Baving, L. Transcranial oscillatory direct current stimulation during sleep improves declarative memory consolidation in children with attention-deficit/hyperactivity disorder to a level comparable to healthy controls. Brain Stim. 2014, 7, 793–799. [Google Scholar] [CrossRef]
- Munz, M.; Prehn-Kristensen, A.; Thielking, F.; Mölle, M.; Göder, R.; Baving, L. Slow oscillating transcranial direct current stimulation during non-rapid eye movement sleep improves behavioral inhibition in attention-deficit/hyperactivity disorder. Front. Cell. Neurosci. 2015, 9, 307. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Ngo, H.V.; Lentfer, L.; Berghäuser, J.; Brandes, L.; Schulze, L.; Göder, R.; Mölle, M.; Baving, L. Acoustic closed-loop stimulation during sleep improves consolidation of reward-related memory information in healthy children but not in children with attention-deficit hyperactivity disorder. Sleep. 2020, 43, zsaa017. [Google Scholar] [CrossRef]
- Munz, M.; Baving, L.; Prehn-Kristensen, A. Transient Destabilization of Declarative Memory-Opposing Impact of Physical Exercise or Rest after Encoding in Typically Developing Children and Children with Attention Deficit Hyperactivity Disorder but No Difference after Subsequent Sleep. Brain Sci. 2022, 12, 322. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Molzow, I.; Munz, M.; Wilhelm, I.; Müller, K.; Freytag, D.; Wiesner, C.D.; Baving, L. Sleep restores daytime deficits in procedural memory in children with attention-deficit/hyperactivity disorder. Res. Dev. Disabil. 2011, 32, 2480–2488. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV-TR; American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- Delmo, C.; Weiffenbach, O.; Gabriel, M.; Bölte, S.; Marchio, E.; Poustka, F. Kiddie-SADS-Present and Lifetime Version (K-SADS-PL), 3rd ed.; Clinic of Child and Adolescent Psychiatry: Frankfurt, Germany, 2000. [Google Scholar]
- Kaufman, J.; Birmaher, B.; Brent, D.; Rao, U.; Flynn, C.; Moreci, P.; Williamson, D.; Ryan, N. Schedule for Affective Disorders and Schizophrenia for School-Age Children-Present and Lifetime Version (K-SADS-PL): Initial reliability and validity data. J. Am. Acad. Child. Adolesc. Psychiatry 1997, 36, 980–988. [Google Scholar] [CrossRef]
- Achenbach, T.M. Manual for the Child Behavior Checklist/4-18 and 1991 Profile; Department of Psychiatry, University of Vermont: Burlington, VT, USA, 1991. [Google Scholar]
- Weiß, R.; Albinus, B.; Arzt, D. Grundintelligenztest Skala 2-Revision (CFT 20-R); Hogrefe: Göttingen, Germany, 2006. [Google Scholar]
- Petersen, A.C.; Crockett, L.; Richards, M.; Boxer, A. A self-report measure of pubertal status: Reliability, validity, and initial norms. J. Youth Adolesc. 1988, 17, 117–133. [Google Scholar] [CrossRef]
- Doty, R.L.; Frye, R.E.; Agrawal, U. Internal consistency reliability of the fractionated and whole University of Pennsylvania Smell Identification Test. Percept. Psychophys. 1989, 45, 381–384. [Google Scholar] [CrossRef]
- Hoffmann, R.M.; Müller, T.; Hajak, G.; Cassel, W. Abend-Morgenprotokolle in Schlafforschung und Schlafmedizin—Ein Instrument für den deutschsprachigen Raum. Somnologie 1997, 1, 103–109. [Google Scholar] [CrossRef]
- Snodgrass, J.G.; Corwin, J. Pragmatics of measuring recognition memory: Applications to dementia and amnesia. J. Exp. Psychol. Gen. 1988, 117, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley and Sons: New York, NY, USA, 2000; pp. 160–164. [Google Scholar]
- World Health Organization. ICD-11 for Mortality and Morbidity Statistics. Available online: https://icd.who.int/browse11/l-m/en (accessed on 7 July 2022).
- Benhalima, K.; Mathieu, C. Gestational diabetes: Update of screening strategy and diagnostic criteria. Curr Opin Obstet Gynecol. 2013, 25, 462–467. [Google Scholar] [CrossRef]
- Bied, A.; Biederman, J.; Faraone, S. Parent-based diagnosis of ADHD is as accurate as a teacher-based diagnosis of ADHD. Postgrad. Med. 2017, 129, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Bruchmüller, K.; Margraf, J.; Schneider, S. Is ADHD diagnosed in accord with diagnostic criteria? Overdiagnosis and influence of client gender on diagnosis. J. Consult. Clin. Psychol. 2012, 80, 128–138. [Google Scholar] [CrossRef]
- Schecklmann, M.; Schwenck, C.; Taurines, R.; Freitag, C.; Warnke, A.; Gerlach, M.; Romanos, M. A systematic review on olfaction in child and adolescent psychiatric disorders. J. Neural. Transm. 2013, 120, 121–130. [Google Scholar] [CrossRef]
- Hutchison, I.C.; Rathore, S. The role of REM sleep theta activity in emotional memory. Front. Psychol. 2015, 6, 1439. [Google Scholar] [CrossRef]
- Takehara-Nishiuchi, K. Entorhinal cortex and consolidated memory. Neurosci Res. 2014, 84, 27–33. [Google Scholar] [CrossRef]
- Battaglia, S.; Thayer, J.F. Functional interplay between central and autonomic nervous systems in human fear conditioning. Trends Neurosci. 2022, 45, 504–506. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ADHD | TDC | ADHD vs. TDC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sleep Group n =14 M(SE) | Wake Group n =14 M(SE) | Sleep vs. Wake | Total n = 28 M(SE) | Sleep Group n = 15 M(SE) | Wake Group n = 15 M(SE) | Sleep vs. Wake | Total n = 30 M(SE) | t(56) | p | |||
| t(26) | p | t(28) | p | |||||||||
| Age | 10.5 (0.19) | 10.4 (0.30) | 0.4 | 0.704 | 10.5 (0.17) | 10.7 (0.40) | 10.5 (0.32) | 0.4 | 0.696 | 10.6 (0.25) | 0.4 | 0.720 |
| PDS | 3.4 (0.17) | 3.1 (0.29) | 1.1 | 0.295 | 3.3 (0.18) | 3.3 (0.15) | 3.1 (0.07) | 1.2 | 0.242 | 3.2 (0.08) | 0.4 | 0.652 |
| IQ | 101.9 (2.54) | 106.7 (5.17) | 0.9 | 0.390 | 104.1 (2,72) | 108.4 (5.00) | 111.2 (2.67) | 0.5 | 0.599 | 110.1 (2.50) | 1.6 | 0.111 |
| CSHQ score | 45.4 (1.15) | 44.8 (1.70) | 0.3 | 0.767 | 45.1 (1.01) | 38.5 (0.70) | 38.9 (0.72) | 0.4 | 0.694 | 38.7 (0.50) | 5.9 | <0.001 |
| ASEBA (T-values) | ||||||||||||
| Withdrawn/Depr. | 62.1 (1.97) | 62.4 (2.50) | 0.5 | 0.912 | 62.3 (1.56) | 53.5 (1.36) | 52.1 (0.98) | 0.8 | 0.410 | 52.8 (0.83) | 5.5 | <0.001 |
| Somatic Complaints | 59.9 (2.82) | 59.2 (2.57) | 0.4 | 0.868 | 59.5 (1.88) | 53.1 (1.07) | 52.2 (1.02) | 0.6 | 0.533 | 52.7 (0.73) | 3.5 | <0.001 |
| Anxious/Depressed | 62.6 (1.62) | 61.5 (2.70) | 0.4 | 0.737 | 62.0 (1.55) | 52.5 (1.10) | 52.9 (1.13) | 0.2 | 0.802 | 52.7 (0.78) | 5.5 | <0.001 |
| Social Problems | 62.1 (2.05) | 63.1 (2.72) | 0.4 | 0.771 | 62.6 (1.67) | 52.2 (1.07) | 52.3 (1.08) | 0.0 | 0.956 | 52.2 (0.75) | 5.8 | <0.001 |
| Thought Problems | 57.6 (2.39) | 56.4 (2.55) | 0.4 | 0.731 | 57.0 (1.72) | 51.2 (0.82) | 50.6 (0.60) | 0.6 | 0.559 | 50.9 (0.50) | 3.5 | <0.001 |
| Attention Problems | 66.9 (2.22) | 68.3 (1.89) | 0.3 | 0.629 | 67.6 (1.44) | 50.8 (0.61) | 50.9 (0.61) | 0.1 | 0.939 | 50.9 (0.42) | 11.4 | <0.001 |
| Rule-Breaking Beh. | 64.2 (2.31) | 61.9 (2.34) | 0.2 | 0.494 | 63.1 (1.63) | 51.3 (0.61) | 50.4 (0.21) | 1.3 | 0.188 | 50.8 (0.33) | 7.6 | <0.001 |
| Aggressive Beh. | 64.6 (2.45) | 63.6 (2.53) | 0.4 | 0.794 | 64.1 (1.73) | 50.9 (0.50) | 50.6 (0.45) | 0.4 | 0.697 | 50.8 (0.33) | 7.8 | <0.001 |
| Internal | 62.8 (2.44) | 62.3 (2.93) | 0.4 | 0.897 | 62.5 (1.87) | 48.4 (2.18) | 47.1 (2.21) | 0.4 | 0.686 | 47.8 (1.52) | 6.1 | <0.001 |
| External | 64.4 (2.79) | 63.4 (2.52) | 0.4 | 0.792 | 63.9 (1.85) | 43.3 (1.82) | 42.7 (1.70) | 0.2 | 0.812 | 43.0 (1.23) | 9.5 | <0.001 |
| Total | 66.4 (2.46) | 66.3 (2.31) | 0.5 | 0.967 | 66.4 (1.66) | 44.9 (2.23) | 43.2 (1.94) | 0.6 | 0.562 | 44.1 (1.46) | 10.1 | <0.001 |
| B | SE | Wald | df | p | |
|---|---|---|---|---|---|
| Intensity | 1.309 | 0.340 | 14.810 | 1 | <0.001 |
| Constant | −7.637 | 1.999 | 14.598 | 1 | <0.001 |
| Predicted | ||||
|---|---|---|---|---|
| TDC | ADHD | % Correct | ||
| observed | TDC | 22 | 8 | 73.3 |
| ADHD | 7 | 21 | 75.0 | |
| Total | 74.1 | |||
| Classification Measure | Value |
|---|---|
| accuracy [95% CI] | 0.741 [0.610; 0.847] |
| no information rate | 0.517 |
| p-value of ACC > NIR | <0.001 |
| sensitivity | 0.750 |
| specificity | 0.733 |
| positive prediction value | 0.7241 |
| negative prediction value | 0.7586 |
| balanced Accuracy | 0.741 |
| ADHD | TDC | ADHD vs. TDC | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sleep Group | Wake Group | Total | Sleep Group | Wake Group | Total | ||||||
| M(SE) | M(SE) | M(SE) | M(SE) | M(SE) | M(SE) | F(56) | p | η2 | |||
| Encoding | Targets | Intensity | 6.59 (0.27) | 6.51 (0.25) | 6.55 (0.18) | 4.88 (0.23) | 5.27 (0.28) | 5.07 (0.18) | 32.2 | <0.001 | 0.374 |
| Pleasantness | 5.41 (0.39) | 4.76 (0.38) | 5.09 (0.28) | 4.64 (0.34) | 4.79 (0.28) | 4.72 (0.22) | 1.1 | 0.290 | 0.021 | ||
| Unpleasantness | 2.42 (0.38) | 2.52 (0.44) | 2.47 (0.28) | 2.15 (0.31) | 1.91 (0.29) | 2.03 (0.21) | 1.5 | 0.220 | 0.028 | ||
| Familiarity | 5.66 (0.42) | 5.45 (0.46) | 5.55 (0.31) | 4.17 (0.32) | 4.58 (0.34) | 4.38 (0.23) | 9.4 | 0.003 | 0.149 | ||
| Retrieval | Targets | Intensity | 6.79 (0.29) | 6.48 (0.30) | 6.63 (0.21) | 4.98 (0.26) | 5.49 (0.30) | 5.24 (0.20) | 23.3 | <0.001 | 0.302 |
| Pleasantness | 4.94 (0.41) | 4.69 (0.47) | 4.81 (0.31) | 4.65 (0.37) | 5.01 (0.25) | 4.83 (0.22) | 0 | 0.967 | 0 | ||
| Unpleasantness | 2.99 (0.37) | 3.03 (0.41) | 3.01 (0.27) | 2.51 (0.34) | 2.17 (0.28) | 2.34 (0.22) | 3.7 | 0.060 | 0.064 | ||
| Familiarity | 5.24 (0.58) | 5.36 (0.39) | 5.30 (0.34) | 4.43 (0.47) | 5.12 (0.35) | 4.78 (0.29) | 1.3 | 0.254 | 0.024 | ||
| Confidence | 6.56 (0.29) | 6.29 (0.30) | 6.42 (0.21) | 5.23 (0.36) | 5.33 (0.51) | 5.28 (0.31) | 9.3 | 0.004 | 0.143 | ||
| Distractors | Intensity | 6.88 (0.24) | 6.46 (0.24) | 6.67 (0.17) | 5.03 (0.26) | 5.37 (0.30) | 5.20 (0.20) | 31.3 | <0.001 | 0.367 | |
| Pleasantness | 4.46 (0.40) | 4.38 (0.42) | 4.42 (0.28) | 4.45 (0.42) | 4.57 (0.32) | 4.51 (0.26) | 0.1 | 0.807 | 0.010 | ||
| Unpleasantness | 4.01 (0.81) | 3.59 (0.41) | 3.75 (0.45) | 2.41 (0.31) | 2.50 (0.31) | 2.45 (0.22) | 6.9 | 0.011 | 0.113 | ||
| Familiarity | 4.42 (0.54) | 4.51 (0.41) | 4.46 (0.33) | 4.03 (0.39) | 4.15 (0.45) | 4.09 (0.29) | 0.7 | 0.404 | 0.013 | ||
| Confidence | 6.39 (0.29) | 5.89 (0.31) | 6.13 (0.22) | 4.84 (0.39) | 5.43 (0.41) | 5.13 (0.28) | 7.8 | 0.007 | 0.126 | ||
| ADHD | TDC | ||||
|---|---|---|---|---|---|
| Sleep Group (M(SE) | Wake Group M(SE) | Sleep Group M(SE) | Wake Group M(SE) | ||
| Encoding | Mood | 5.5 (0.17) | 5.5 (0.20) | 4.0 (0.24) | 4.5 (0.17) |
| Tiredness | 4.3 (0.34) | 3.7 (0.44) | 3.1 (0.25) | 3.2 (0.39) | |
| Arousal | 4.6 (0.40) | 3.6 (0.44) | 3.7 (0.32) | 2.5 (0.22) | |
| Retrieval | Mood | 5.4 (0.17) | 5.6 (0.20) | 4.1 (0.24) | 4.2 (0.22) |
| Tiredness | 3.8 (0.45) | 4.4 (0.41) | 2.8 (0.24) | 3.5 (0.32) | |
| Arousal | 5.0 (0.21) | 4.7 (0.29) | 2.5 (0.34) | 3.7 (0.23) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munz, M.; Wiesner, C.D.; Vollersen-Krekiehn, M.; Baving, L.; Prehn-Kristensen, A. Sleep Fosters Odor Recognition in Children with Attention Deficit Hyperactivity Disorder but Not in Typically Developing Children. Brain Sci. 2022, 12, 1182. https://doi.org/10.3390/brainsci12091182
Munz M, Wiesner CD, Vollersen-Krekiehn M, Baving L, Prehn-Kristensen A. Sleep Fosters Odor Recognition in Children with Attention Deficit Hyperactivity Disorder but Not in Typically Developing Children. Brain Sciences. 2022; 12(9):1182. https://doi.org/10.3390/brainsci12091182
Chicago/Turabian StyleMunz, Manuel, Christian Dirk Wiesner, Meike Vollersen-Krekiehn, Lioba Baving, and Alexander Prehn-Kristensen. 2022. "Sleep Fosters Odor Recognition in Children with Attention Deficit Hyperactivity Disorder but Not in Typically Developing Children" Brain Sciences 12, no. 9: 1182. https://doi.org/10.3390/brainsci12091182
APA StyleMunz, M., Wiesner, C. D., Vollersen-Krekiehn, M., Baving, L., & Prehn-Kristensen, A. (2022). Sleep Fosters Odor Recognition in Children with Attention Deficit Hyperactivity Disorder but Not in Typically Developing Children. Brain Sciences, 12(9), 1182. https://doi.org/10.3390/brainsci12091182