
Neural Correlates of Delay Discounting in the Light of Brain Imaging and Non-Invasive Brain Stimulation: What We Know and What Is Missed
 , ,
, ,
Abstract
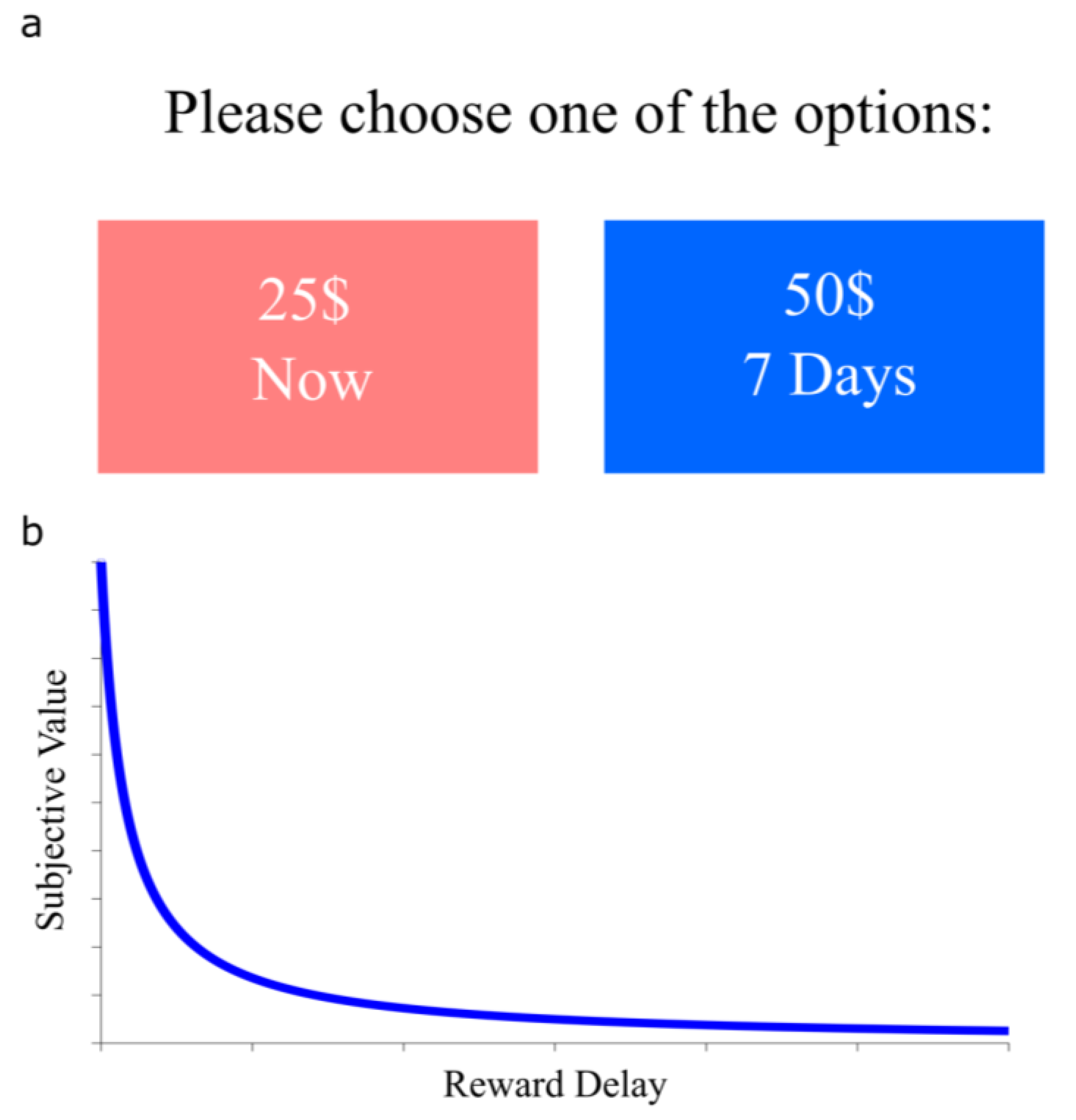
1. Introduction
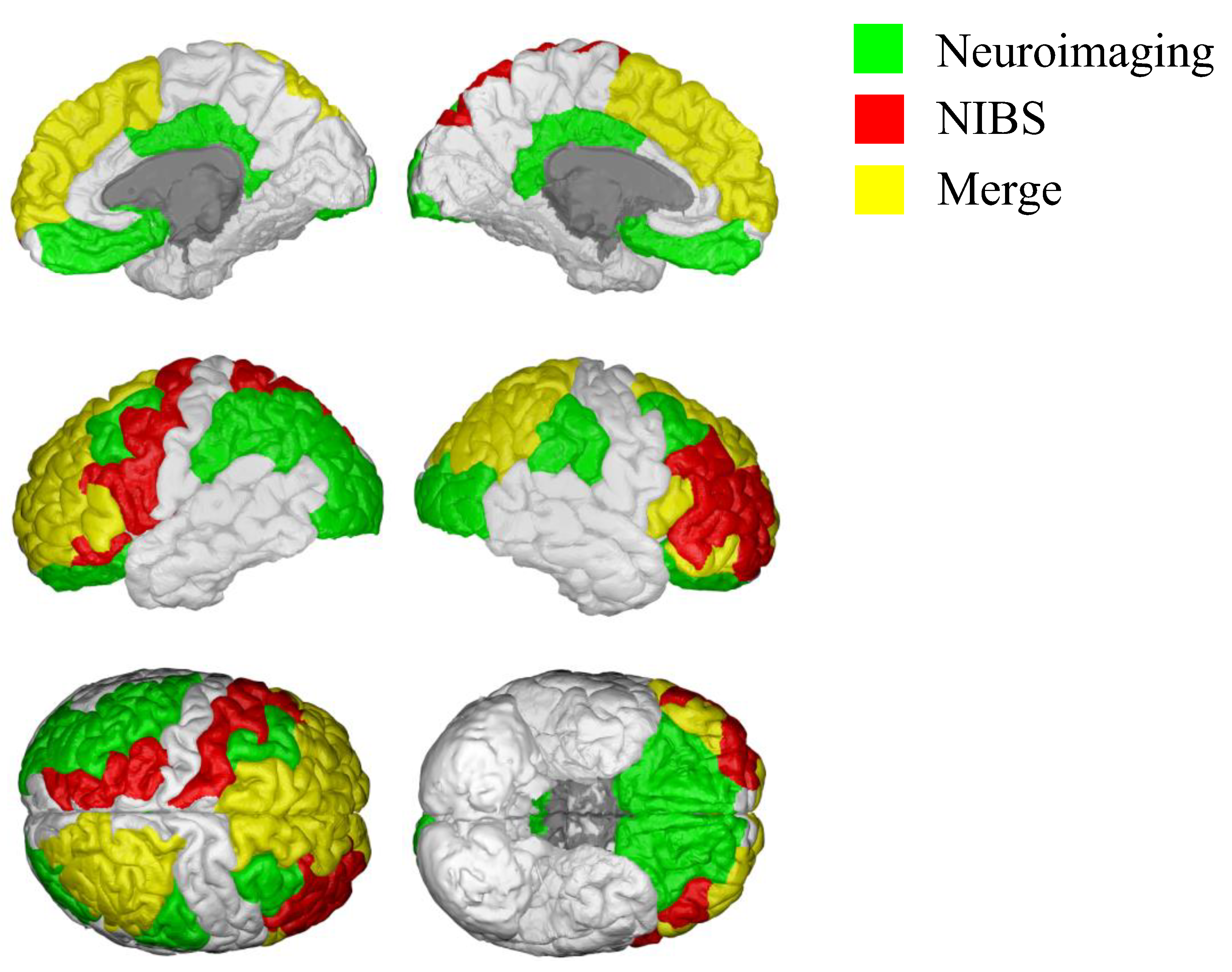
2. Neural Correlates of Delay Discounting Assessed by Neuroimaging Studies
3. Investigating the Causal Role of Neural Circuits in Delay Discounting
Effects of Transcranial Magnetic Stimulation (TMS) Neuromodulation on Delay Discounting

{kind=link}
{kind=link}
| Study | Brain Area | Protocol | Stimulation Intensity | n | Disease | Discounting Assessment | Effect |
|---|---|---|---|---|---|---|---|
| Cho et al., 2015 [51] | mPFC | 10 Hz-rTMS | 80% MT | 24 | - | ln (k) | Decreased delay discounting |
| Zack et al., 2016 [40] | mPFC | rTMS | 80% MT | 9 | PG | k | - |
| Zack et al., 2016 [40] | Right dlPFC | TBS | 80% MT | 9 | PG | k | - |
| Cho et al., 2010 [42] | Right dlPFC | cTBS | 80% MT | 7 | - | k | Decreased delay discounting |
| Figner et al., 2010 [12] | Right dlPFC | 1 Hz low-frequency rTMS | 54% MT | 19 | - | Immediate choices (%) | - |
| Cho et al., 2012 [46] | Right dlPFC | cTBS | 80% MT | 8 | - | ln (k) | Decreased delay discounting |
| Schluter et al., 2019 [43] | Right dlPFC | HF-rTMS | 110% MT | 40 | AUD | AUC | - |
| Ballard et al., 2018 [45] | Right dlPFC | 1 Hz low-frequency rTMS | 120% MT | 12 | - | log (k) | Increased delay discounting |
| Essex et al., 2012 [44] | Right dlPFC + right PPC | 1 Hz low-frequency rTMS | 54% MT | 16 | - | Immediate choices (%) | Decreased delay discounting |
| Teti Mayer et al., 2019 [39] | Left dlPFC | 10 Hz | 110% MT | 20 | MDD | k | - |
| Sheffer et al., 2013 [41] | Left dlPFC | HF rTMS | 110% MT | 47 | Smokers | k | Decreased delay discounting |
| Figner et al., 2010 [12] | Left dlPFC | 1 Hz low-frequency rTMS | 54% MT | 18 | - | Immediate choices (%) | Increased delay discounting |
| Ballard et al., 2018 [45] | Left dlPFC | 1 Hz low-frequency rTMS | 120% MT | 15 | - | log (k) | Increased delay discounting |
| Yang et al., 2018 [52] | Left dlPFC | iTBS | 80% MT | 23 | - | ln (k) | - |
| Essex et al., 2012 [44] | Left dlPFC + left PPC | 1 Hz low-frequency rTMS | 54% MT | 16 | - | Immediate choices (%) | Increased delay discounting |
| Soutschek et al., 2016 [49] | Right pTPJ | cTBS | 80% MT | 22 | - | log (k) | Increased delay discounting |
| Soutschek et al., 2016 [49] | Right pTPJ | cTBS | 80% MT | 20 | - | log (k) | Increased delay discounting |
| Soutschek et al., 2016 [49] | Left S1 | cTBS | 80% MT | 21 | - | log (k) | - |
4. Effects of Transcranial Direct Current Stimulation (tDCS) Neuromodulation on Delay Discounting
| Study | Anode Position | Cathode Position | Current Intensity | n | Disease | Discounting Assessment | Effect |
|---|---|---|---|---|---|---|---|
| Manuel at al., 2019 [63] | vmPFC | Vertex | 2.0 mA | 20 | - | log (k) | Decreased delay discounting |
| Nejati et al., 2021 [61] | Right vmPFC | Left dlPFC | 1.0 mA | 20 | ADHD children | k | Decreased delay discounting |
| Wang et al., 2021 [67] | FPC | Vertex | 1.5 mA | 90 | - | k | - |
| Soutschek et al., 2017 [68] | Left FPC | Vertex | 1.0 mA | 27 | - | Immediate choices (%) | - |
| To et al., 2018 [69] | Right IFG | Left IFG | 2.0 mA | 23 | Chocolate cravers | k | - |
| Nejati et al., 2018 [13] | Right OFC | Left dlPFC | 1.5 mA | 24 | - | k | Decreased delay discounting |
| He at al., 2016 [55] | Right dlPFC | - | 1.5 mA | 23 | - | k | - |
| Shen et al., 2016 [56] | Right dlPFC | Left dlPFC | 2.0 mA | 39 | - | k | - |
| Shen et al., 2016 [56] | Right dlPFC | - | 2.0 mA | 39 | - | k | - |
| Xiong et al., 2019 [58] | Right dlPFC | Left dlPFC | 1.5 mA | 20 | - | k | - |
| Kekic et al., 2017 [57] | Right dlPFC | Left dlPFC | 2.0 mA | 39 | BN | δ | Decreased delay discounting |
| Kekic et al., 2014 [70] | Right dlPFC | Left dlPFC | 2.0 mA | 17 | Food cravers | k | - |
| Hecht et al., 2013 [71] | Right dlPFC | Left dlPFC | 1.6 mA | 14 | - | Immediate choices (%) | Decreased delay discounting |
| Brunelin and Fecteau, 2021 [59] | Left dlPFC | Right dlPFC | 2.0 mA | 15 | Acutely stressed | Immediate choices (%) | Decreased delay discounting |
| He at al., 2016 [55] | Left dlPFC | - | 1.5 mA | 22 | - | k | Decreased delay discounting |
| Shen et al., 2016 [56] | Left dlPFC | Right dlPFC | 2.0 mA | 39 | - | k | - |
| Shen et al., 2016 [56] | Left dlPFC | - | 2.0 mA | 39 | - | k | Decreased delay discounting |
| Nejati et al., 2018 [13] | Left dlPFC | Right OFC | 1.5 mA | 24 | - | k | Decreased delay discounting |
| Terenzi et al., 2021 [62] | Left dlPFC | Right shoulder | 1.5 mA | 28 | PD | log (k) | - |
| Nejati et al., 2021 [61] | Left dlPFC | Right vmPFC | 1.0 mA | 20 | ADHD children | k | - |
| Xiong et al., 2019 [58] | Left dlPFC | Right dlPFC | 2.0 mA | 20 | - | k | Decreased delay discounting |
| Kekic et al., 2017 [57] | Left dlPFC | Right dlPFC | 2.0 mA | 39 | BN | δ | Decreased delay discounting |
| Gaudreault et al., 2021 [72] | Left dlPFC | Right dlPFC | 2.0 mA | 17 | CUD | k | Decreased delay discounting |
| Colombo et al., 2020 [60] | Left dlPFC | Right triceps | 1.5 mA | 13 | - | RT | - |
| Hecht et al., 2013 [71] | Left dlPFC | Right dlPFC | 1.6 mA | 14 | - | Immediate choices (%) | Decreased delay discounting |
| Manuel at al., 2019 [63] | Vertex | vmPFC | 2.0 mA | 20 | - | log (k) | - |
| Wang et al., 2021 [67] | Vertex | FPC | 1.5 mA | 90 | - | k | - |
| Soutschek et al., 2017 [68] | Vertex | Left FPC | 1.0 mA | 26 | - | Immediate choices (%) | - |
| Terenzi et al., 2021 [62] | Left M1 | Right shoulder | 1.5 mA | 28 | PD | log (k) | - |
| Wynn et al., 2019 [64] | Medial cerebellum | Right deltoid muscle | 2.0 mA | 26 | - | AUC | - |
| Colombo et al., 2020 [60] | Right triceps | Left dlPFC | 1.5 mA | 13 | - | RT | Increased delay discounting |
| Shen et al., 2016 [56] | - | Right dlPFC | 2.0 mA | 39 | - | k | - |
| Shen et al., 2016 [56] | - | Left dlPFC | 2.0 mA | 39 | - | k | Increased delay discounting |
5. Conclusions, Limitations and Future Developments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mischel, W.; Shoda, Y.; Rodriguez, M.L. Delay of Gratification in Children. Science 1989, 244, 933–938. [Google Scholar] [CrossRef]
- Mazur, J.E. An adjusting procedure for studying delayed reinforcement. In The Effect of Delay and of Intervening Events on Reinforcement Value; Lawrence Erlbaum Associates, Inc: Hillsdale, NJ, USA, 1987; pp. 55–73. [Google Scholar]
- Vanderveldt, A.; Oliveira, L.; Green, L. Delay discounting: Pigeon, rat, human—Does it matter? J. Exp. Psychol. Anim. Learn. Cogn. 2016, 42, 141–162. [Google Scholar] [CrossRef]
- Hernandez, C.M.; Orsini, C.A.; Labiste, C.C.; Wheeler, A.R.; Eyck, T.W.T.; Bruner, M.M.; Sahagian, T.J.; Harden, S.W.; Frazier, C.J.; Setlow, B.; et al. Optogenetic dissection of basolateral amygdala contributions to intertemporal choice in young and aged rats. Elife 2019, 8, e46174. [Google Scholar] [CrossRef]
- de Wit, H. Impulsivity as a determinant and consequence of drug use: A review of underlying processes. Addict. Biol. 2009, 14, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Castellanos-Ryan, N.; Struve, M.; Whelan, R.; Banaschewski, T.; Barker, G.J.; Bokde, A.L.W.; Bromberg, U.; Büchel, C.; Flor, H.; Fauth-Bühler, M.; et al. Neural and Cognitive Correlates of the Common and Specific Variance Across Externalizing Problems in Young Adolescence. Am. J. Psychiatry 2014, 171, 1310–1319. [Google Scholar] [CrossRef]
- Amlung, M.; Marsden, E.; Holshausen, K.; Morris, V.; Patel, H.; Vedelago, L.; Naish, K.R.; Reed, D.D.; McCabe, R.E. Delay Discounting as a Transdiagnostic Process in Psychiatric Disorders: A Meta-analysis. JAMA Psychiatry 2019, 76, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, J.; Sulpizio, S.; Ferro, M.; Martoni, R.; Abutalebi, J.; Malgaroli, A. Behavioral assessment of activity-based-anorexia: How cognition can become the drive wheel. Physiol. Behav. 2019, 202, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McClure, S.M.; Laibson, D.I.; Loewenstein, G.; Cohen, J.D. Separate neural systems value immediate and delayed monetary rewards. Science 2004, 306, 503–507. [Google Scholar] [CrossRef]
- Kable, J.W.; Glimcher, P.W. The neural correlates of subjective value during intertemporal choice. Nat. Neurosci. 2007, 10, 1625–1633. [Google Scholar] [CrossRef]
- Ballard, K.; Knutson, B. Dissociable neural representations of future reward magnitude and delay during temporal discounting. Neuroimage 2009, 45, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Figner, B.; Knoch, D.; Johnson, E.J.; Krosch, A.R.; Lisanby, S.H.; Fehr, E.; Weber, E.U. Lateral prefrontal cortex and self-control in intertemporal choice. Nat. Neurosci. 2010, 13, 538–539. [Google Scholar] [CrossRef]
- Nejati, V.; Salehinejad, M.A.; Nitsche, M.A. Interaction of the Left Dorsolateral Prefrontal Cortex (l-DLPFC) and Right Orbitofrontal Cortex (OFC) in Hot and Cold Executive Functions: Evidence from Transcranial Direct Current Stimulation (tDCS). Neuroscience 2018, 369, 109–123. [Google Scholar] [CrossRef]
- Scherbaum, S.; Frisch, S.; Dshemuchadse, M. Step by step: Harvesting the dynamics of delay discounting decisions. Q. J. Exp. Psychol. 2018, 71, 949–964. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.; McNaughton, N. The neural basis of delay discounting: A review and preliminary model. Neurosci. Biobehav. Rev. 2017, 79, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Heekeren, H.R.; Marrett, S.; Ungerleider, L.G. The neural systems that mediate human perceptual decision making. Nat. Rev. Neurosci. 2008, 9, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Yu, R. Regional white matter volumes correlate with delay discounting. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, B.C.; Smallwood, J.; Tusche, A.; Ruby, F.J.M.; Engen, H.G.; Steinbeis, N.; Singer, T. Medial prefrontal and anterior cingulate cortical thickness predicts shared individual differences in self-generated thought and temporal discounting. Neuroimage 2014, 90, 290–297. [Google Scholar] [CrossRef]
- Moschak, T.M.; Carelli, R.M. Impulsive rats exhibit blunted dopamine release dynamics during a delay discounting task independent of cocaine history. eNeuro 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Xu, S.; Das, G.; Hueske, E.; Tonegawa, S. Dorsal Raphe Serotonergic Neurons Control Intertemporal Choice under Trade-off. Curr. Biol. 2017, 27, 3111–3119.e3. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, H.; Lu, C.; Lu, T.; Froudist-Walsh, S.; Chen, M.; Wang, X.J.; Hu, J.; Sun, W. The neural basis of delayed gratification. Sci. Adv. 2021, 7, 111470. [Google Scholar] [CrossRef]
- McClure, S.M.; Ericson, K.M.; Laibson, D.I.; Loewenstein, G.; Cohen, J.D. Time discounting for primary rewards. J. Neurosci. 2007, 27, 5796–5804. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Büchel, C. Overlapping and distinct neural systems code for subjective value during intertemporal and risky decision making. J. Neurosci. 2009, 29, 15727–15734. [Google Scholar] [CrossRef]
- Casey, B.J.; Somerville, L.H.; Gotlib, I.H.; Ayduk, O.; Franklin, N.T.; Askren, M.K.; Jonides, J.; Berman, M.G.; Wilson, N.L.; Teslovich, T.; et al. Behavioral and neural correlates of delay of gratification 40 years later. Proc. Natl. Acad. Sci. USA 2011, 108, 14998–15003. [Google Scholar] [CrossRef]
- Hare, T.A.; Hakimi, S.; Rangel, A. Activity in dlPFC and its effective connectivity to vmPFC are associated with temporal discounting. Front. Neurosci. 2014, 8, 50. [Google Scholar] [CrossRef]
- van den Bos, W.; Rodriguez, C.A.; Schweitzer, J.B.; McClure, S.M. Connectivity strength of dissociable striatal tracts predict individual differences in temporal discounting. J. Neurosci. 2014, 34, 10298–10310. [Google Scholar] [CrossRef] [PubMed]
- Szuhany, K.L.; MacKenzie, D.; Otto, M.W. The impact of depressed mood, working memory capacity, and priming on delay discounting. J. Behav. Ther. Exp. Psychiatry 2018, 60, 37–41. [Google Scholar] [CrossRef]
- Peters, J.; Büchel, C. Episodic Future Thinking Reduces Reward Delay Discounting through an Enhancement of Prefrontal-Mediotemporal Interactions. Neuron 2010, 66, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Fellows, L.K.; Farah, M.J. Dissociable elements of human foresight: A role for the ventromedial frontal lobes in framing the future, but not in discounting future rewards. Neuropsychologia 2005, 43, 1214–1221. [Google Scholar] [CrossRef]
- Sellitto, M.; Ciaramelli, E.; Di Pellegrino, G. Myopic discounting of future rewards after medial orbitofrontal damage in humans. J. Neurosci. 2010, 30, 16429–16436. [Google Scholar] [CrossRef]
- Rochkind, S. Photobiomodulation in Neuroscience: A Summary of Personal Experience. Photomed. Laser Surg. 2017, 35, 604–615. [Google Scholar] [CrossRef]
- Chen, Z.; Tsytsarev, V.; Finfrock, Y.Z.; Antipova, O.A.; Cai, Z.; Arakawa, H.; Lischka, F.W.; Hooks, B.M.; Wilton, R.; Wang, D.; et al. Wireless Optogenetic Modulation of Cortical Neurons Enabled by Radioluminescent Nanoparticles. ACS Nano 2021, 15, 5201–5208. [Google Scholar] [CrossRef] [PubMed]
- Sahel, J.-A.; Boulanger-Scemama, E.; Pagot, C.; Arleo, A.; Galluppi, F.; Martel, J.N.; Esposti, S.D.; Delaux, A.; de Saint Aubert, J.-B.; de Montleau, C.; et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 2021, 27, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Darmani, G.; Bergmann, T.O.; Butts Pauly, K.; Caskey, C.F.; de Lecea, L.; Fomenko, A.; Fouragnan, E.; Legon, W.; Murphy, K.R.; Nandi, T.; et al. Non-invasive transcranial ultrasound stimulation for neuromodulation. Clin. Neurophysiol. 2022, 135, 51–73. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmöller, J.; Carpenter, L.I.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; et al. Safety and recommendations for TMS use in healthy subjects and patient populations, with updates on training, ethical and regulatory issues: Expert Guidelines. Clin. Neurophysiol. 2021, 132, 269–306. [Google Scholar] [CrossRef] [PubMed]
- Ferro, M.; Lamanna, J.; Spadini, S.; Nespoli, A.; Sulpizio, S.; Malgaroli, A. Synaptic plasticity mechanisms behind TMS efficacy: Insights from its application to animal models. J. Neural Transm. 2022, 129, 25–36. [Google Scholar] [CrossRef]
- Garcia-Sanz, S.; Ghotme, K.A.; Hedmont, D.; Arévalo-Jaimes, M.Y.; Cohen Kadosh, R.; Serra-Grabulosa, J.M.; Redolar-Ripoll, D. Use of transcranial magnetic stimulation for studying the neural basis of numerical cognition: A systematic review. J. Neurosci. Methods 2022, 369, 109485. [Google Scholar] [CrossRef]
- Bestmann, S. The physiological basis of transcranial magnetic stimulation. Trends Cogn. Sci. 2008, 12, 81–83. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Dileone, M.; Pilato, F.; Capone, F.; Musumeci, G.; Ranieri, F.; Ricci, V.; Bria, P.; Di Iorio, R.; de Waure, C.; et al. Modulation of motor cortex neuronal networks by rTMS: Comparison of local and remote effects of six different protocols of stimulation. J. Neurophysiol. 2011, 105, 2150–2156. [Google Scholar] [CrossRef]
- Teti Mayer, J.; Nicolier, M.; Tio, G.; Mouchabac, S.; Haffen, E.; Bennabi, D. Effects of High Frequency Repetitive Transcranial Magnetic Stimulation (HF-rTMS) on Delay Discounting in Major Depressive Disorder. Encephale 2019, 45, S68. [Google Scholar] [CrossRef]
- Zack, M.; Cho, S.S.; Parlee, J.; Jacobs, M.; Li, C.; Boileau, I.; Strafella, A. Effects of High Frequency Repeated Transcranial Magnetic Stimulation and Continuous Theta Burst Stimulation on Gambling Reinforcement, Delay Discounting, and Stroop Interference in Men with Pathological Gambling. Brain Stimul. 2016, 9, 867–875. [Google Scholar] [CrossRef]
- Sheffer, C.E.; Mennemeier, M.; Landes, R.D.; Bickel, W.K.; Brackman, S.; Dornhoffer, J.; Kimbrell, T.; Brown, G. Neuromodulation of delay discounting, the reflection effect, and cigarette consumption. J. Subst. Abuse Treat. 2013, 45, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.S.; Ko, J.H.; Pellecchia, G.; Van Eimeren, T.; Cilia, R.; Strafella, A.P. Continuous theta burst stimulation of right dorsolateral prefrontal cortex induces changes in impulsivity level. Brain Stimul. 2010, 3, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Schluter, R.S.; van Holst, R.J.; Goudriaan, A.E. Effects of Ten Sessions of High Frequency Repetitive Transcranial Magnetic Stimulation (HF-rTMS) Add-on Treatment on Impulsivity in Alcohol Use Disorder. Front. Neurosci. 2019, 13, 1257. [Google Scholar] [CrossRef]
- Essex, B.G.; Clinton, S.A.; Wonderley, L.R.; Zald, D.H. The impact of the posterior parietal and dorsolateral prefrontal cortices on the optimization of long-term versus immediate value. J. Neurosci. 2012, 32, 15403–15413. [Google Scholar] [CrossRef] [PubMed]
- Ballard, I.C.; Aydogan, G.; Kim, B.; McClure, S.M. Causal Evidence for the Dependence of the Magnitude Effect on Dorsolateral Prefrontal Cortex. Sci. Rep. 2018, 8, 16545. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.S.; Pellecchia, G.; Ko, J.H.; Ray, N.; Obeso, I.; Houle, S.; Strafella, A.P. Effect of continuous theta burst stimulation of the right dorsolateral prefrontal cortex on cerebral blood flow changes during decision making. Brain Stimul. 2012, 5, 116–123. [Google Scholar] [CrossRef]
- Yang, C.C.; Khalifa, N.; Lankappa, S.; Völlm, B. Effects of intermittent theta burst stimulation applied to the left dorsolateral prefrontal cortex on empathy and impulsivity in healthy adult males. Brain Cogn. 2018, 128, 37–45. [Google Scholar] [CrossRef]
- Cho, S.S.; Koshimori, Y.; Aminian, K.; Obeso, I.; Rusjan, P.; Lang, A.E.; Daskalakis, Z.J.; Houle, S.; Strafella, A.P. Investing in the future: Stimulation of the medial prefrontal cortex reduces discounting of delayed rewards. Neuropsychopharmacology 2015, 40, 546–553. [Google Scholar] [CrossRef]
- Soutschek, A.; Ruff, C.C.; Strombach, T.; Kalenscher, T.; Tobler, P.N. Brain stimulation reveals crucial role of overcoming self-centeredness in self-control. Sci. Adv. 2016, 2, 2–10. [Google Scholar] [CrossRef]
- Do, M.; Byrne, L.K.; Pearce, A.J. Examining the feasibility and tolerability of a clinically informed multisite, repetitive transcranial magnetic stimulation protocol. J. Neurosci. Methods 2016, 258, 24–27. [Google Scholar] [CrossRef]
- Das, S.; Holland, P.; Frens, M.A.; Donchin, O. Impact of Transcranial Direct Current Stimulation (tDCS) on Neuronal Functions. Front. Neurosci. 2016, 10, 550. [Google Scholar] [CrossRef] [PubMed]
- Brunyé, T.T. Non-invasive Brain Stimulation Effects on the Perceptual and Cognitive Processes Underlying Decision-making: A Mini Review. J. Cogn. Enhanc. 2021, 5, 233–244. [Google Scholar] [CrossRef]
- He, Q.; Chen, M.; Chen, C.; Xue, G.; Feng, T.; Bechara, A. Anodal stimulation of the left DLPFC increases IGT scores and decreases delay discounting rate in healthy males. Front. Psychol. 2016, 7, 1421. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Yin, Y.; Wang, J.; Zhou, X.; McClure, S.M.; Li, J. High-definition tDCS alters impulsivity in a baseline-dependent manner. Neuroimage 2016, 143, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Kekic, M.; McClelland, J.; Bartholdy, S.; Boysen, E.; Musiat, P.; Dalton, B.; Tiza, M.; David, A.S.; Campbell, I.C.; Schmidt, U. Single-session transcranial direct current stimulation temporarily improves symptoms, mood, and self-regulatory control in bulimia nervosa: A randomised controlled trial. PLoS ONE 2017, 12, e0167606. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Li, X.; Dong, Z.; Cai, S.; Huang, J.; Li, Q. Modulating activity in the prefrontal cortex changes intertemporal choice for loss: A transcranial direct current stimulation study. Front. Hum. Neurosci. 2019, 13, 167. [Google Scholar] [CrossRef]
- Brunelin, J.; Fecteau, S. Impact of bifrontal transcranial Direct Current Stimulation on decision-making and stress reactivity. A pilot study. J. Psychiatr. Res. 2021, 135, 15–19. [Google Scholar] [CrossRef]
- Colombo, B.; Iannello, P.; Puglisi, G.; Antonietti, A. Personal Impulsivity Mediates the Effects of Neuromodulation in Economic Intertemporal Choices: A Pilot Study. Stud. Psychol. 2020, 62, 123–137. [Google Scholar] [CrossRef]
- Nejati, V.; Sarraj Khorrami, A.; Nitsche, M.A. Transcranial Direct Current Stimulation Improves Reward Processing in Children With ADHD. J. Atten. Disord. 2021, 25, 1623–1631. [Google Scholar] [CrossRef]
- Terenzi, D.; Catalan, M.; Polverino, P.; Bertolotti, C.; Manganotti, P.; Rumiati, R.I.; Aiello, M. Effects of tDCS on reward responsiveness and valuation in Parkinson’s patients with impulse control disorders. J. Neurol. 2022, 269, 1557–1565. [Google Scholar] [CrossRef]
- Manuel, A.L.; Murray, N.W.G.; Piguet, O. Transcranial direct current stimulation (tDCS) over vmPFC modulates interactions between reward and emotion in delay discounting. Sci. Rep. 2019, 9, 18735. [Google Scholar] [CrossRef]
- Wynn, S.C.; Driessen, J.M.A.; Glennon, J.C.; Brazil, I.A.; Schutter, D.J.L.G. Cerebellar Transcranial Direct Current Stimulation Improves Reactive Response Inhibition in Healthy Volunteers. Cerebellum 2019, 18, 983–988. [Google Scholar] [CrossRef]
- To, C.; Falcone, M.; Loughead, J.; Logue-Chamberlain, E.; Hamilton, R.; Kable, J.; Lerman, C.; Ashare, R.L. Got chocolate? Bilateral prefrontal cortex stimulation augments chocolate consumption. Appetite 2018, 131, 28–35. [Google Scholar] [CrossRef]
- Sparing, R.; Mottaghy, F.M. Noninvasive brain stimulation with transcranial magnetic or direct current stimulation (TMS/tDCS)—From insights into human memory to therapy of its dysfunction. Methods 2008, 44, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Datta, A.; Bikson, M.; Parra, L.C. ROAST: An Open-Source, Fully-Automated, Realistic Volumetric-Approach-Based Simulator For TES. In Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Honolulu, HI, USA, 18–21 July 2018; pp. 3072–3075. [Google Scholar]
- Kekic, M.; McClelland, J.; Campbell, I.; Nestler, S.; Rubia, K.; David, A.S.; Schmidt, U. The effects of prefrontal cortex transcranial direct current stimulation (tDCS) on food craving and temporal discounting in women with frequent food cravings. Appetite 2014, 78, 55–62. [Google Scholar] [CrossRef]
- Soutschek, A.; Ugazio, G.; Crockett, M.J.; Ruff, C.C.; Kalenscher, T.; Tobler, P.N. Binding oneself to the mast: Stimulating frontopolar cortex enhances precommitment. Soc. Cogn. Affect. Neurosci. 2017, 12, 635–642. [Google Scholar] [CrossRef]
- Wittmann, M.; Leland, D.S.; Paulus, M.P. Time and decision making: Differential contribution of the posterior insular cortex and the striatum during a delay discounting task. Exp. Brain Res. 2007, 179, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Schoenbaum, G.; Shaham, Y. The Role of Orbitofrontal Cortex in Drug Addiction: A Review of Preclinical Studies. Biol. Psychiatry 2008, 63, 256–262. [Google Scholar] [CrossRef]
- Miyazaki, K.; Miyazaki, K.W.; Sivori, G.; Yamanaka, A.; Tanaka, K.F.; Doya, K. Serotonergic projections to the orbitofrontal and medial prefrontal cortices differentially modulate waiting for future rewards. Sci. Adv. 2020, 6, eabc7246. [Google Scholar] [CrossRef] [PubMed]
- Fettes, P.; Schulze, L.; Downar, J. Cortico-Striatal-Thalamic Loop Circuits of the Orbitofrontal Cortex: Promising Therapeutic Targets in Psychiatric Illness. Front. Syst. Neurosci. 2017, 11, 25. [Google Scholar] [CrossRef]
- Marinescu, R.V.; Eshaghi, A.; Alexander, D.C.; Golland, P. BrainPainter: A software for the visualisation of brain structures, biomarkers and associated pathological processes. In Proceedings of the Multimodal Brain Image Analysis and Mathematical Foundations of Computational Anatomy: 4th International Workshop, MBIA 2019, and 7th International Workshop, MFCA 2019, Shenzhen, China, 17 October 2019; pp. 112–120. [Google Scholar]
- McNamara, R.K.; Liu, Y.; Jandacek, R.; Rider, T.; Tso, P. The aging human orbitofrontal cortex: Decreasing polyunsaturated fatty acid composition and associated increases in lipogenic gene expression and stearoyl-CoA desaturase activity. Prostaglandins Leukot. Essent. Fat. Acids 2008, 78, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Shott, M.E.; Pryor, T.L.; Yang, T.T.; Frank, G.K.W. Greater Insula White Matter Fiber Connectivity in Women Recovered from Anorexia Nervosa. Neuropsychopharmacology 2016, 41, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Miniussi, C.; Ruzzoli, M.; Walsh, V. The mechanism of transcranial magnetic stimulation in cognition. Cortex 2010, 46, 128–130. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moro, A.S.; Saccenti, D.; Ferro, M.; Scaini, S.; Malgaroli, A.; Lamanna, J. Neural Correlates of Delay Discounting in the Light of Brain Imaging and Non-Invasive Brain Stimulation: What We Know and What Is Missed. Brain Sci. 2023, 13, 403. https://doi.org/10.3390/brainsci13030403
Moro AS, Saccenti D, Ferro M, Scaini S, Malgaroli A, Lamanna J. Neural Correlates of Delay Discounting in the Light of Brain Imaging and Non-Invasive Brain Stimulation: What We Know and What Is Missed. Brain Sciences. 2023; 13(3):403. https://doi.org/10.3390/brainsci13030403
Chicago/Turabian StyleMoro, Andrea Stefano, Daniele Saccenti, Mattia Ferro, Simona Scaini, Antonio Malgaroli, and Jacopo Lamanna. 2023. "Neural Correlates of Delay Discounting in the Light of Brain Imaging and Non-Invasive Brain Stimulation: What We Know and What Is Missed" Brain Sciences 13, no. 3: 403. https://doi.org/10.3390/brainsci13030403
APA StyleMoro, A. S., Saccenti, D., Ferro, M., Scaini, S., Malgaroli, A., & Lamanna, J. (2023). Neural Correlates of Delay Discounting in the Light of Brain Imaging and Non-Invasive Brain Stimulation: What We Know and What Is Missed. Brain Sciences, 13(3), 403. https://doi.org/10.3390/brainsci13030403