
Event-Related Potentials Reveal the Impact of Conflict Strength in a Numerical Stroop Paradigm

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
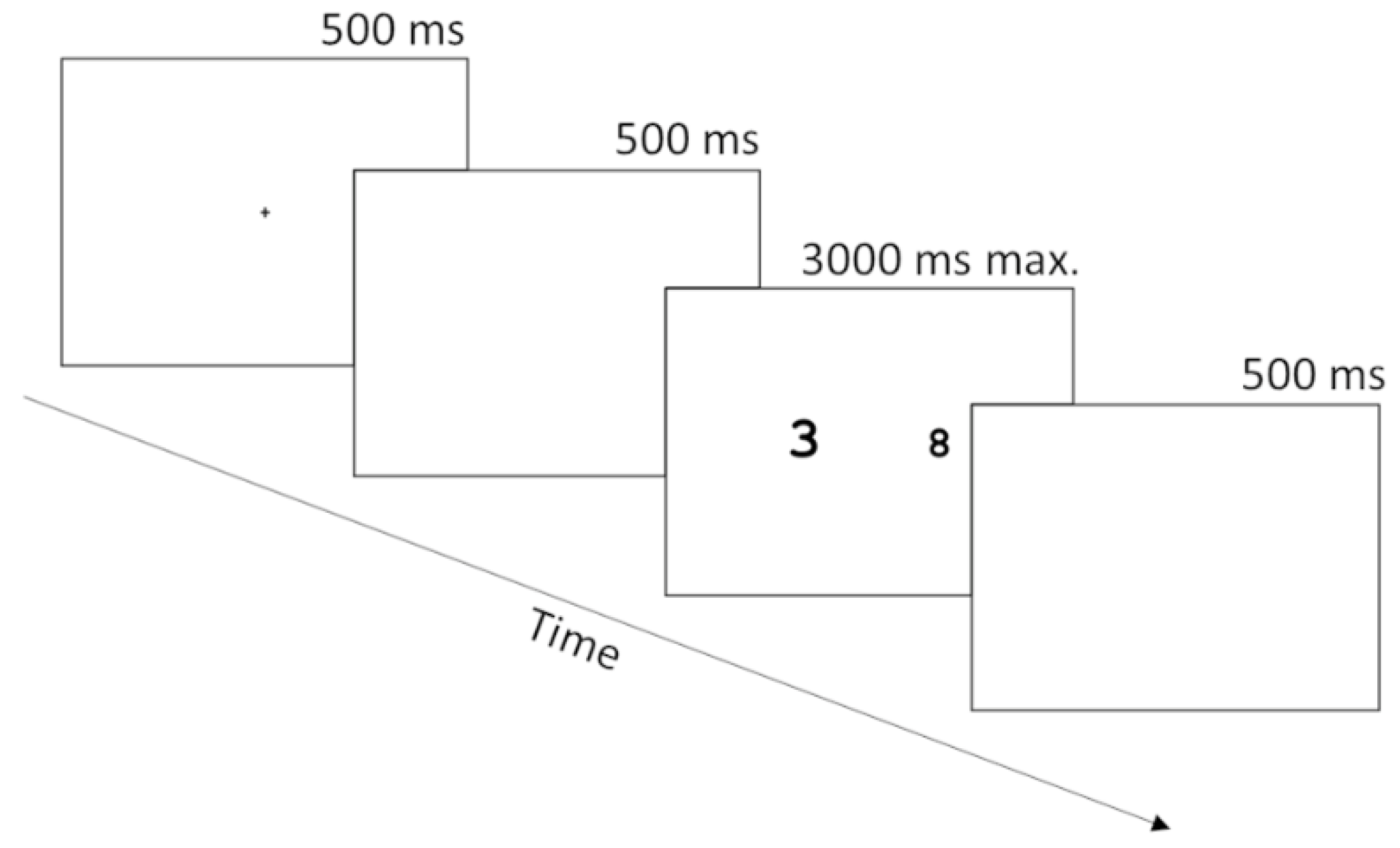
2.2. Task and Procedure
2.3. Electrophysiological Recordings
3. Results
3.1. Behavioral Analyses
3.2. ERP Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Piaget, J. The Child’s Conception of Number; Routledge & Kegan Paul, Ltd.: London, UK, 1952. [Google Scholar]
- Wilkey, E.D.; Price, G.R. Attention to number: The convergence of numerical magnitude processing, attention, and mathematics in the inferior frontal gyrus. Hum. Brain Mapp. 2019, 40, 928–943. [Google Scholar] [CrossRef] [PubMed]
- Gebuis, T.; Reynvoet, B. The interplay between nonsymbolic number and its continuous visual properties. J. Exp. Psychol. Gen. 2012, 141, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Henik, A.; Tzelgov, J. Is three greater than five: The relation between physical and semantic size in comparison tasks. Mem. Cogn. 1982, 10, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Borst, G. Inhibitory Control in Number-Conservation and Class-Inclusion Tasks: A Neo-Piagetian Inter-Task Priming Study. Cogn. Dev. 2012, 27, 283–298. [Google Scholar] [CrossRef]
- Viarouge, A.; Houdé, O.; Borst, G. Evidence for the role of inhibition in numerical comparison: A negative priming study in 7- to 8-year-olds and adults. J. Exp. Child. Psychol. 2019, 186, 131–141. [Google Scholar] [CrossRef]
- Roell, M.; Viarouge, A.; Houdé, O.; Borst, G. Inhibitory control and decimal number comparison in school-aged children. PLoS ONE 2017, 12, e0188276. [Google Scholar] [CrossRef]
- Roell, M.; Viarouge, A.; Houdé, O.; Borst, G. Inhibition of the whole number bias in decimal number comparison: A developmental negative priming study. J. Exp. Child. Psychol. 2019, 177, 240–247. [Google Scholar] [CrossRef]
- Viarouge, A.; Houdé, O.; Borst, G. The progressive 6-year-old conserver: Numerical saliency and sensitivity as core mechanisms of numerical abstraction in a Piaget-like estimation task. Cognition 2019, 190, 137–142. [Google Scholar] [CrossRef]
- Pansky, A.; Algom, D. Stroop and Garner effects in the comparative judgments of numerals: The role of attention. J. Exp. Psychol. Hum. 1999, 25, 39–58. [Google Scholar] [CrossRef]
- Pinel, P.; Piazza, M.; Le Bihan, D.; Dehaene, S. Distributed and Overlapping Cerebral Representations of Number, Size, and Luminance during Comparative Judgments. Neuron 2004, 41, 983–993. [Google Scholar] [CrossRef]
- Borghesani, V.; de Hevia, M.D.; Viarouge, A.; Pinheiro-Chagas, P.; Eger, E.; Piazza, M. Processing number and length in the parietal cortex: Sharing resources, not a common code. Cortex 2019, 114, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, D.; Soltész, F. Event-related potentials dissociate facilitation and interference effects in the numerical Stroop paradigm. Neuropsychologia 2007, 45, 3190–3202. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, D.; Soltész, F. The interaction of task-relevant and task-irrelevant stimulus features in the number/size congruency paradigm: An ERP study. Brain Res. 2008, 1190, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Gebuis, T.; Kenemans, J.L.; de Haan, E.H.F.; van der Smagt, M.J. Conflict processing of symbolic and non-symbolic numerosity. Neuropsychologia 2010, 48, 394–401. [Google Scholar] [CrossRef]
- Libertus, M.E.; Woldorff, M.G.; Brannon, E.M. Electrophysiological evidence for notation independence in numerical processing. Behav. Brain Funct. 2007, 3, 1. [Google Scholar] [CrossRef]
- Gebuis, T.; Reynvoet, B. The neural mechanisms underlying passive and active processing of numerosity. Neuroimage 2013, 70, 301–307. [Google Scholar] [CrossRef]
- Luck, S.J.; Hillyard, S.A. Electrophysiological correlates of feature analysis during visual search. Psychophysiology 1994, 31, 291–308. [Google Scholar] [CrossRef]
- Straube, S.; Grimsen, C.; Fahle, M. Electrophysiological correlates of figure–ground segregation directly reflect perceptual saliency. Vision Res. 2010, 50, 509–521. [Google Scholar] [CrossRef]
- Shoji, H.; Ozaki, H. Topographic change in ERP due to discrimination of geometric figures in the peripheral visual field. Int. J. Psychophysiol. 2006, 62, 115–121. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of singletrial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Mullen, T.R.; Kothe, C.A.E.; Chi, M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.J.; Cauwenberghs, G. Real-time neuroimaging and cognitive monitoring using wearable dry EEG. IEEE Trans. Biomed. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Verleger, R.; Görgen, S.; Jaśkowski, P.; Wascher, E. Evidence for an integrative role of P3b in linking reaction to perception. J. Psychophysiol. 2005, 19, 165–181. [Google Scholar] [CrossRef]
- Piazza, M.; De Feo, V.; Panzeri, S.; Dehaene, S. Learning to focus on number. Cognition 2018, 181, 35–45. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vurdah, N.; Vidal, J.; Viarouge, A. Event-Related Potentials Reveal the Impact of Conflict Strength in a Numerical Stroop Paradigm. Brain Sci. 2023, 13, 586. https://doi.org/10.3390/brainsci13040586
Vurdah N, Vidal J, Viarouge A. Event-Related Potentials Reveal the Impact of Conflict Strength in a Numerical Stroop Paradigm. Brain Sciences. 2023; 13(4):586. https://doi.org/10.3390/brainsci13040586
Chicago/Turabian StyleVurdah, Nydia, Julie Vidal, and Arnaud Viarouge. 2023. "Event-Related Potentials Reveal the Impact of Conflict Strength in a Numerical Stroop Paradigm" Brain Sciences 13, no. 4: 586. https://doi.org/10.3390/brainsci13040586
APA StyleVurdah, N., Vidal, J., & Viarouge, A. (2023). Event-Related Potentials Reveal the Impact of Conflict Strength in a Numerical Stroop Paradigm. Brain Sciences, 13(4), 586. https://doi.org/10.3390/brainsci13040586