Postnatal Development of Synaptic Plasticity at Hippocampal CA1 Synapses: Correlation of Learning Performance with Pathway-Specific Plasticity



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
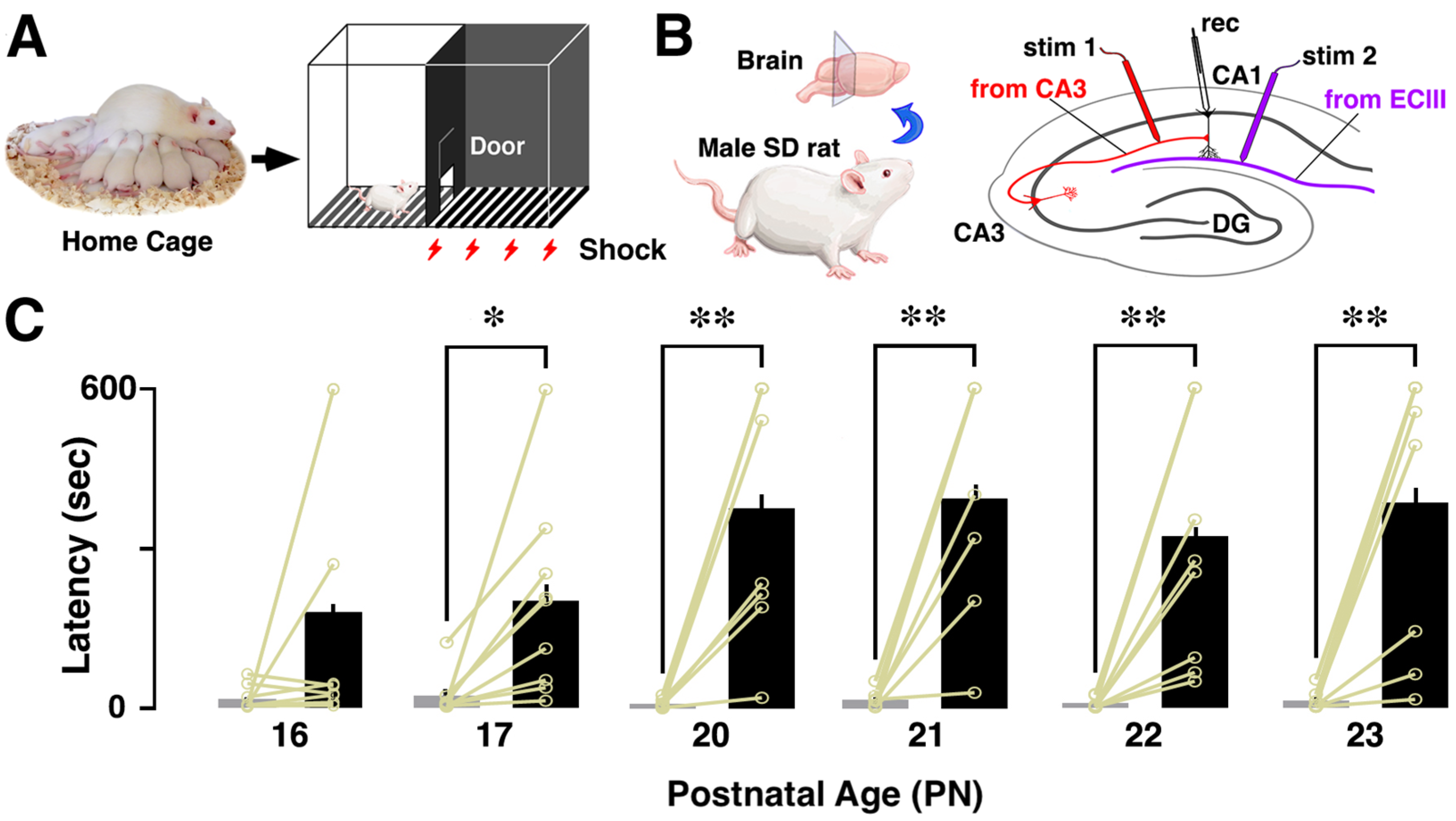
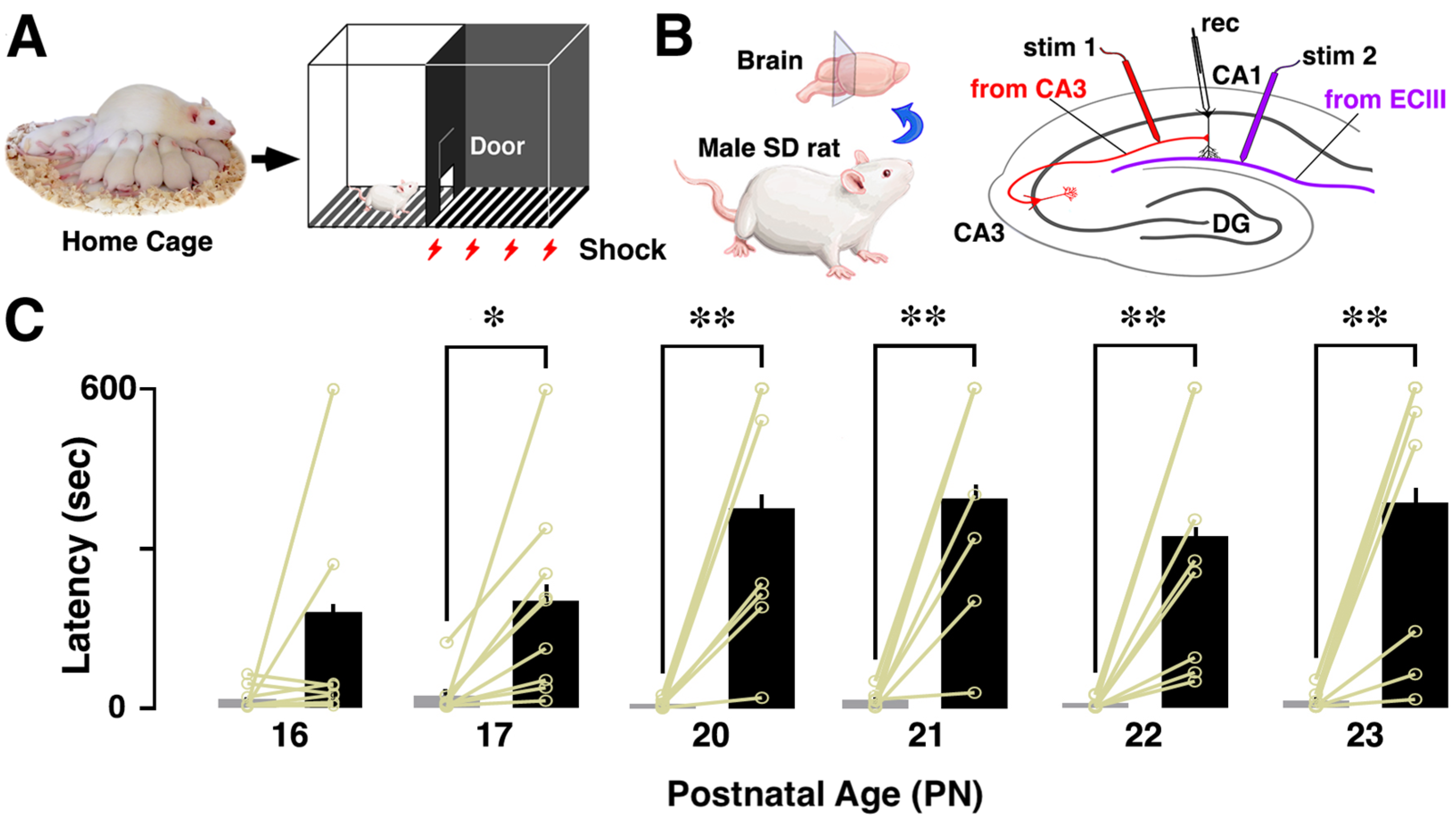
2.2. Inhibitory Avoidance (IA) Task
2.3. Electrophysiological Recording of Slice-Patch Clamping
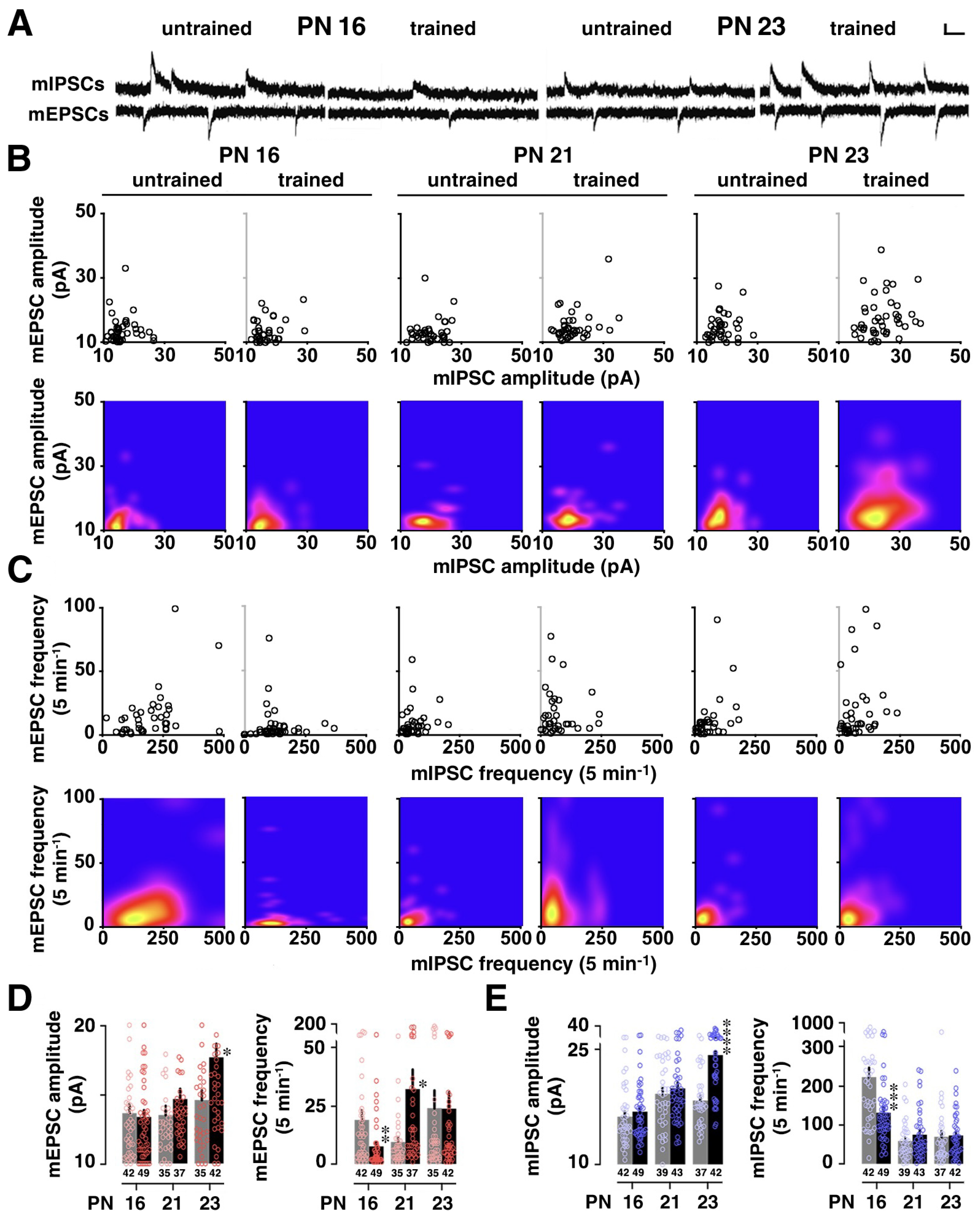
2.4. Miniature Postsynaptic Current Recordings
2.5. Statistical Analysis
3. Results
3.1. Developmental Change in Contextual Learning Performance
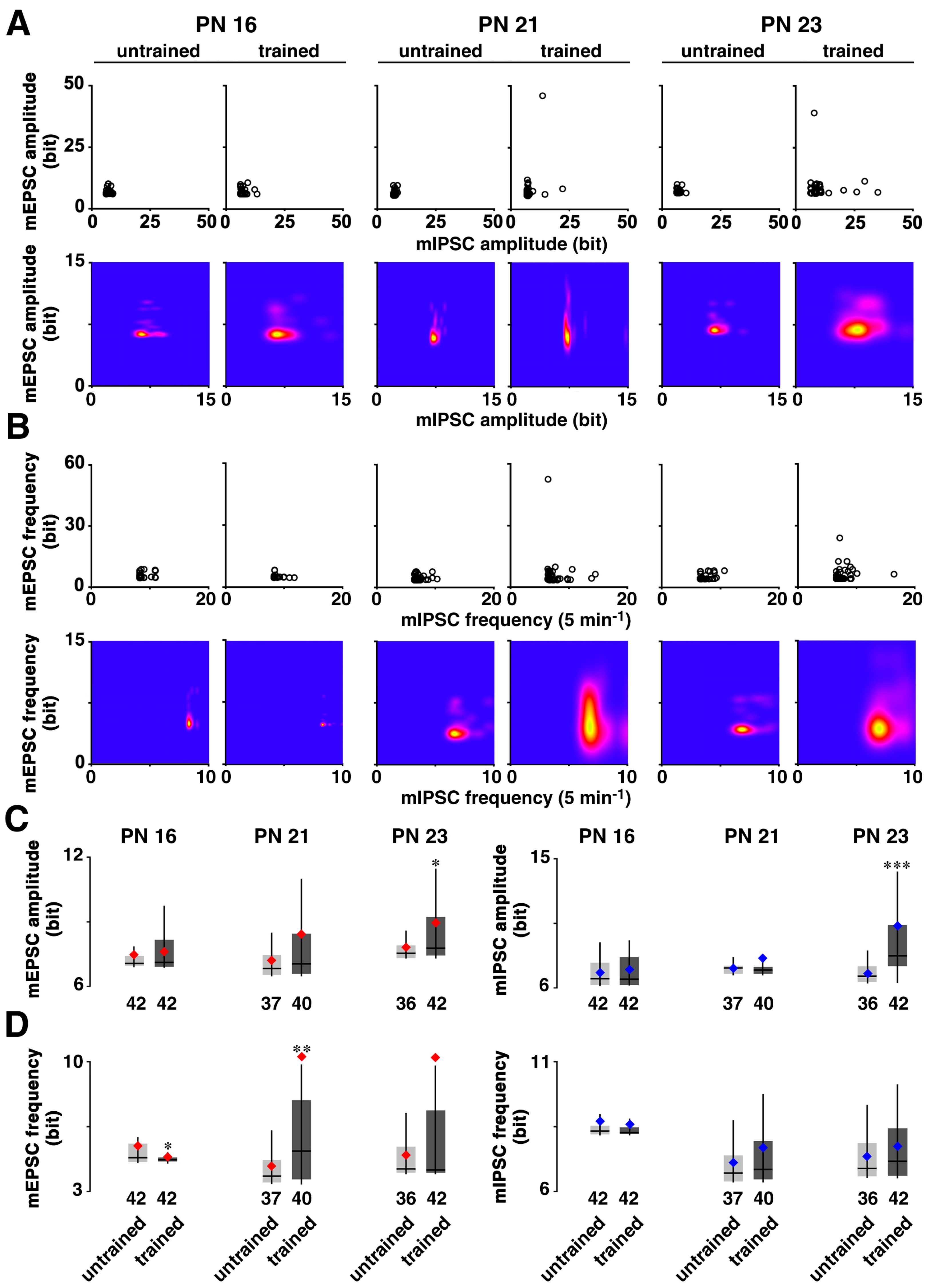
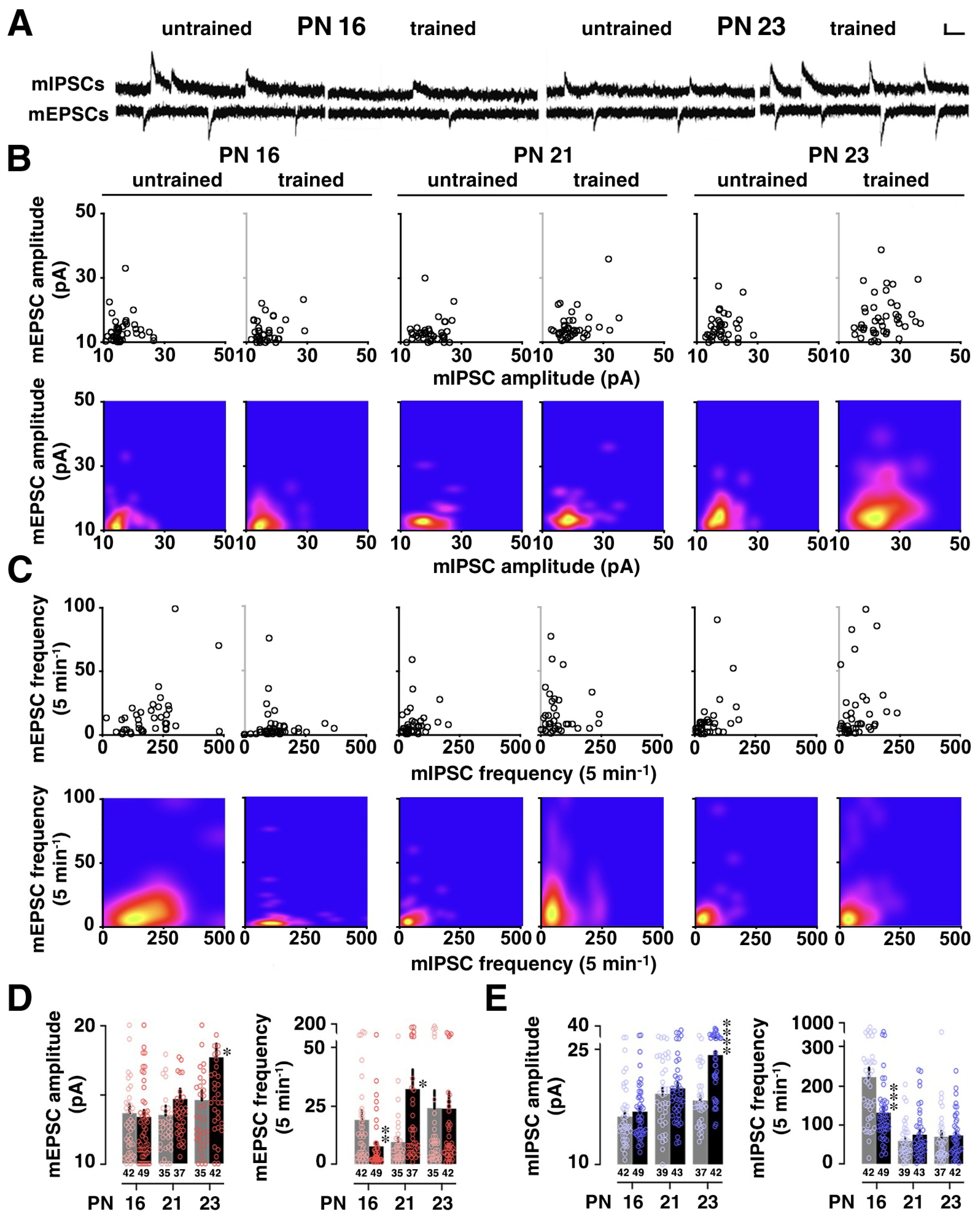
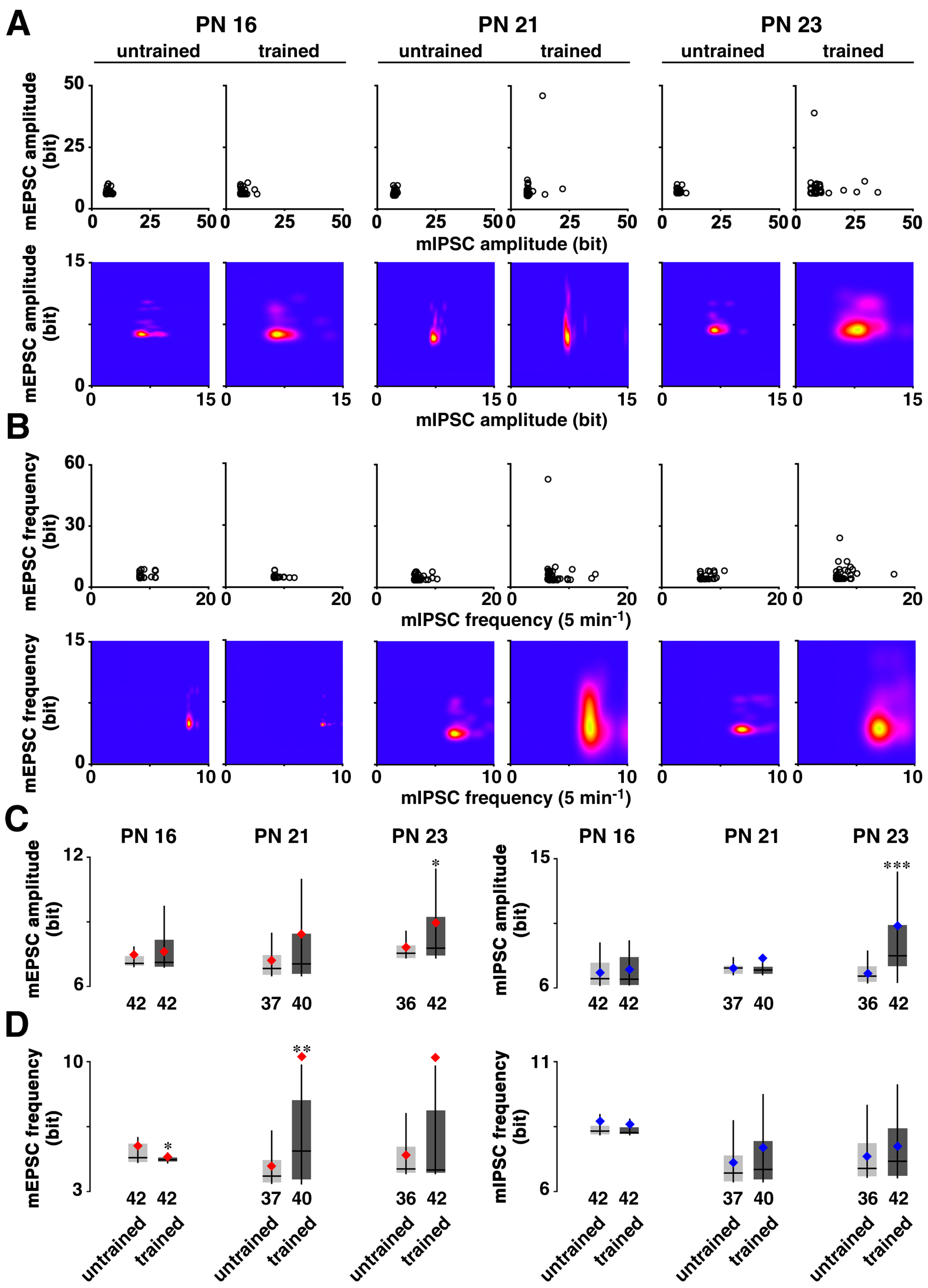
3.2. Developmental Change in Training-Induced Diversity at Excitatory and Inhibitory Synapses
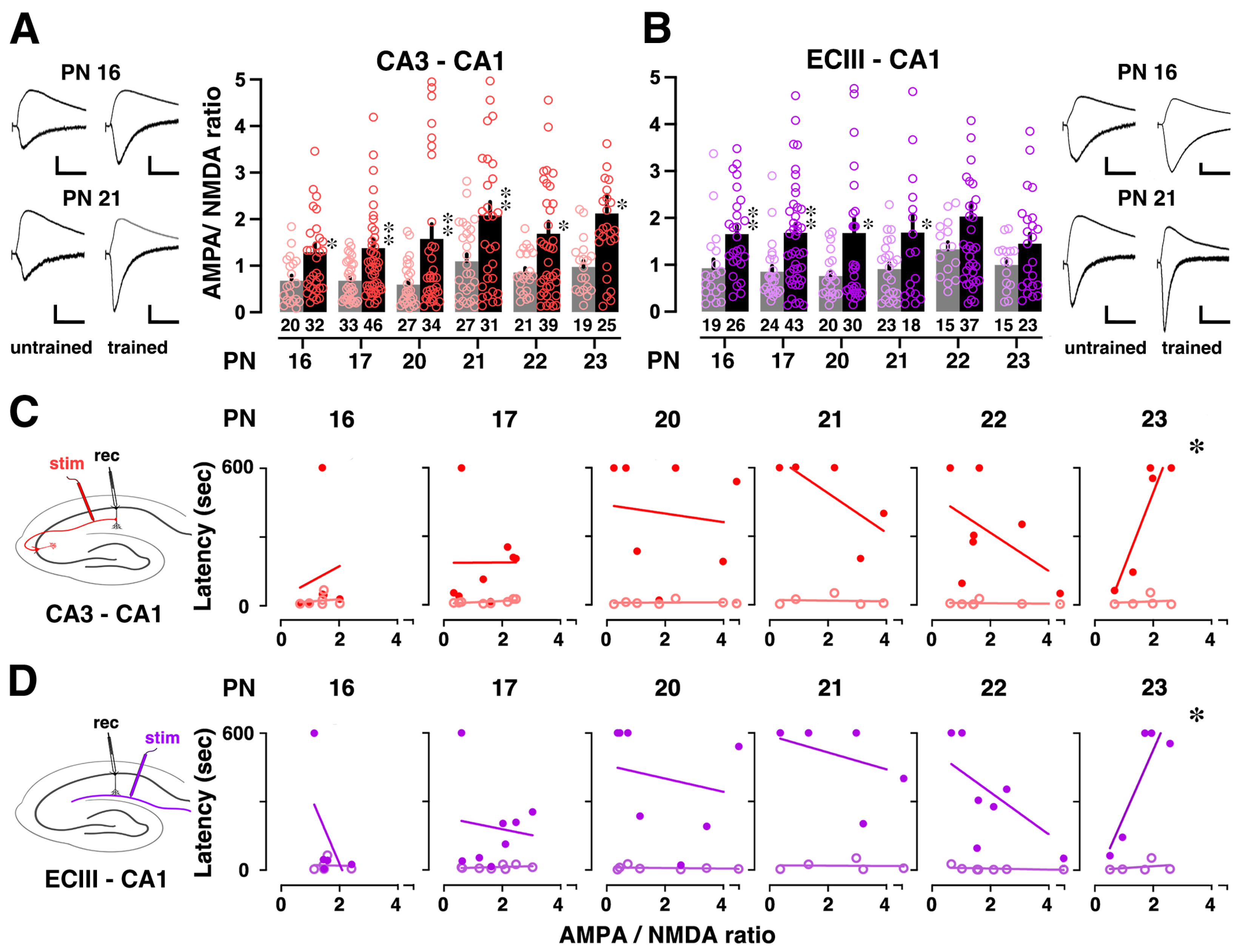
3.3. Training-Induced Plasticity at CA3-CA1 or ECIII-CA1 Synapses
3.4. Correlation between Training-Induced Plasticity and Learning Performance
4. Discussion
Implications and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scoville, W.B.; Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, 11. [Google Scholar] [CrossRef]
- O’Keefe, J.; Krupic, J. Do hippocampal pyramidal cells respond to nonspatial stimuli? Physiol. Rev. 2021, 101, 1427–1456. [Google Scholar] [CrossRef] [PubMed]
- Wills, T.J.; Cacucci, F.; Burgess, N.; O’Keefe, J. Development of the hippocampal cognitive map in preweanling rats. Science 2010, 328, 1573–1576. [Google Scholar] [CrossRef]
- Gelbard-Sagiv, H.; Mukamel, R.; Harel, M.; Malach, R.; Fried, I. Internally generated reactivation of single neurons in human hippocampus during free recall. Science 2008, 322, 96–101. [Google Scholar] [CrossRef]
- Ishikawa, J.; Tomokage, T.; Mitsushima, D. A possible coding for experience: Ripple-like events and synaptic diversity. BioRxiv 2019. [Google Scholar] [CrossRef]
- Campbell, B.A.; Campbell, E.H. Retention and extinction of learned fear in infant and adult rats. J. Comp. Physiol. Psychol. 1962, 55, 1. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.A.; Spear, N.E. Ontogeny of memory. Psychol. Rev. 1972, 79, 215. [Google Scholar] [CrossRef] [PubMed]
- Josselyn, S.A.; Frankland, P.W. Infantile amnesia: A neurogenic hypothesis. Learn. Mem. 2012, 19, 423–433. [Google Scholar] [CrossRef]
- Travaglia, A.; Bisaz, R.; Sweet, E.S.; Blitzer, R.D.; Alberini, C.M. Infantile amnesia reflects a developmental critical period for hippocampal learning. Nat. Neurosci. 2016, 19, 1225–1233. [Google Scholar] [CrossRef]
- Baudry, M.; Arst, D.S.; Lynch, G. Increased [3H] glutamate receptor binding in aged rats. Brain Res. 1981, 223, 195–198. [Google Scholar] [CrossRef]
- Baudry, M.; Arst, D.; Oliver, M.; Lynch, G. Development of glutamate binding sites and their regulation by calcium in rat hippocampus. Dev. Brain Res. 1981, 1, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Steward, O.; Falk, P.M. Selective localization of polyribosomes beneath developing synapses: A quantitative analysis of the relationships between polyribosomes and developing synapses in the hippocampus and dentate gyrus. J. Comp. Neurol. 1991, 314, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Collin, C.; Miyaguchi, K.; Segal, M. Dendritic spine density and LTP induction in cultured hippocampal slices. J. Neurophysiol. 1997, 77, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.J.; Carpenter, D.O. Development of synaptic responses and plasticity at the SC-CA1 and MF-CA3 synapses in rat hippocampus. Cell. Mol. Neurobiol. 2001, 21, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Cizeron, M.; Qiu, Z.; Koniaris, B.; Gokhale, R.; Komiyama, N.H.; Fransén, E.; Grant, S.G. A brainwide atlas of synapses across the mouse life span. Science 2020, 369, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Ramsaran, A.I.; Wang, Y.; Golbabaei, A.; Aleshin, S.; de Snoo, M.L.; Yeung, B.-r.A.; Rashid, A.J.; Awasthi, A.; Lau, J.; Tran, L.M. A shift in the mechanisms controlling hippocampal engram formation during brain maturation. Science 2023, 380, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Steward, O.; Scoville, S.A. Cells of origin of entorhinal cortical afferents to the hippocampus and fascia dentata of the rat. J. Comp. Neurol. 1976, 169, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P. The Hippocampus Book; Oxford University Press: Oxford, UK, 2007. [Google Scholar] [CrossRef]
- Kajiwara, R.; Wouterlood, F.G.; Sah, A.; Boekel, A.J.; Baks-te Bulte, L.T.; Witter, M.P. Convergence of entorhinal and CA3 inputs onto pyramidal neurons and interneurons in hippocampal area CA1—An anatomical study in the rat. Hippocampus 2008, 18, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Paw-Min-Thein-Oo; Sakimoto, Y.; Kida, H.; Mitsushima, D. Proximodistal heterogeneity in learning-promoted pathway-specific plasticity at dorsal CA1 synapses. Neuroscience 2020, 437, 184–195. [Google Scholar] [CrossRef]
- Mitsushima, D.; Sano, A.; Takahashi, T. A cholinergic trigger drives learning-induced plasticity at hippocampal synapses. Nat. Commun. 2013, 4, 2760. [Google Scholar] [CrossRef]
- Sakimoto, Y.; Shintani, A.; Yoshiura, D.; Goshima, M.; Kida, H.; Mitsushima, D. A critical period for learning and plastic changes at hippocampal CA1 synapses. Sci. Rep. 2022, 12, 7199. [Google Scholar] [CrossRef] [PubMed]
- Kida, H.; Sakimoto, Y.; Mitsushima, D. Slice patch clamp technique for analyzing learning-induced plasticity. J. Vis. Exp. 2017, 129, e55876. [Google Scholar] [CrossRef]
- Sakimoto, Y.; Mizuno, J.; Kida, H.; Kamiya, Y.; Ono, Y.; Mitsushima, D. Learning promotes subfield-specific synaptic diversity in hippocampal CA1 neurons. Cereb. Cortex 2019, 29, 2183–2195. [Google Scholar] [CrossRef] [PubMed]
- Barker, G.R.; Bird, F.; Alexander, V.; Warburton, E.C. Recognition memory for objects, place, and temporal order: A disconnection analysis of the role of the medial prefrontal cortex and perirhinal cortex. J. Neurosci. 2007, 27, 2948–2957. [Google Scholar] [CrossRef] [PubMed]
- Nadler, J.V.; Matthews, D.A.; Cotman, C.W.; Lynch, G.S. Development of cholinergic innervation in the hippocampal formation of the rat: II. Quantitative changes in choline acetyltransferase and acetylcholinesterase activities. Dev. Biol. 1974, 36, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Bronzino, J.D.; Abu-Hasaballah, K.; Austin-LaFrance, R.J.; Morgane, P.J. Maturation of long-term potentiation in the hippocampal dentate gyrus of the freely moving rat. Hippocampus 1994, 4, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Takumi, Y.; Ramírez-León, V.; Laake, P.; Rinvik, E.; Ottersen, O.P. Different modes of expression of AMPA and NMDA receptors in hippocampal synapses. Nat. Neurosci. 1999, 2, 618–624. [Google Scholar] [CrossRef]
- Pickard, L.; Noel, J.; Henley, J.M.; Collingridge, G.L.; Molnar, E. Developmental changes in synaptic AMPA and NMDA receptor distribution and AMPA receptor subunit composition in living hippocampal neurons. J. Neurosci. 2000, 20, 7922–7931. [Google Scholar] [CrossRef]
- Travaglia, A.; Bisaz, R.; Cruz, E.; Alberini, C.M. Developmental changes in plasticity, synaptic, glia and connectivity protein levels in rat dorsal hippocampus. Neurobiol. Learn. Mem. 2016, 135, 125–138. [Google Scholar] [CrossRef]
- Kudryashov, I.E.; Onufriev, M.V.; Kudryashova, I.V.; Gulyaeva, N.V. Periods of postnatal maturation of hippocampus: Synaptic modifications and neuronal disconnection. Dev. Brain Res. 2001, 132, 113–120. [Google Scholar] [CrossRef]
- Maldonado, P.P.; Vélez-Fort, M.; Levavasseur, F.; Angulo, M.C. Oligodendrocyte precursor cells are accurate sensors of local K+ in mature gray matter. J. Neurosci. 2013, 33, 2432–2442. [Google Scholar] [CrossRef] [PubMed]
- Vélez-Fort, M.; Maldonado, P.P.; Butt, A.M.; Audinat, E.; Angulo, M.C. Postnatal switch from synaptic to extrasynaptic transmission between interneurons and NG2 cells. J. Neurosci. 2010, 30, 6921–6929. [Google Scholar] [CrossRef] [PubMed]
- Behuet, S.; Cremer, J.N.; Cremer, M.; Palomero-Gallagher, N.; Zilles, K.; Amunts, K. Developmental changes of glutamate and GABA receptor densities in wistar rats. Front. Neuroanat. 2019, 13, 100. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Harris, K.M. Developmental regulation of the late phase of long-term potentiation (L-LTP) and metaplasticity in hippocampal area CA1 of the rat. J. Neurophysiol. 2012, 107, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Teyler, T.; Perkins IV, A.; Harris, K.M. The development of long-term potentiation in hippocampus and neocortex. Neuropsychologia 1989, 27, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Dumas, T.C. Postnatal alterations in induction threshold and expression magnitude of long-term potentiation and long-term depression at hippocampal synapses. Hippocampus 2012, 22, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Bolshakov, V.Y.; Siegelbaum, S.A. Postsynaptic induction and presynaptic expression of hippocampal long-term depression. Science 1994, 264, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Kemp, N.; McQueen, J.; Faulkes, S.; Bashir, Z.I. Different forms of LTD in the CA1 region of the hippocampus: Role of age and stimulus protocol. Eur. J. Neurosci. 2000, 12, 360–366. [Google Scholar] [CrossRef]
- Bekenstein, J.W.; Lothman, E.W. A comparison of the ontogeny of excitatory and inhibitory neurotransmission in the CA1 region and dentate gyrus of the rat hippocampal formation. Dev. Brain Res. 1991, 63, 237–243. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Postnatal Days | Body Weight (g) | Number of Rats |
|---|---|---|
| 16 | 36.1 ± 1.3 | 14 |
| 17 | 38.8 ± 1.4 | 14 |
| 20 | 45.3 ± 1.8 | 13 |
| 21 | 49.5 ± 1.6 | 16 |
| 22 | 51.9 ± 1.6 | 22 |
| 23 | 55.3 ± 2.4 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Sakimoto, Y.; Mitsushima, D. Postnatal Development of Synaptic Plasticity at Hippocampal CA1 Synapses: Correlation of Learning Performance with Pathway-Specific Plasticity. Brain Sci. 2024, 14, 382. https://doi.org/10.3390/brainsci14040382
Yang Y, Sakimoto Y, Mitsushima D. Postnatal Development of Synaptic Plasticity at Hippocampal CA1 Synapses: Correlation of Learning Performance with Pathway-Specific Plasticity. Brain Sciences. 2024; 14(4):382. https://doi.org/10.3390/brainsci14040382
Chicago/Turabian StyleYang, Yuheng, Yuya Sakimoto, and Dai Mitsushima. 2024. "Postnatal Development of Synaptic Plasticity at Hippocampal CA1 Synapses: Correlation of Learning Performance with Pathway-Specific Plasticity" Brain Sciences 14, no. 4: 382. https://doi.org/10.3390/brainsci14040382
APA StyleYang, Y., Sakimoto, Y., & Mitsushima, D. (2024). Postnatal Development of Synaptic Plasticity at Hippocampal CA1 Synapses: Correlation of Learning Performance with Pathway-Specific Plasticity. Brain Sciences, 14(4), 382. https://doi.org/10.3390/brainsci14040382