
Combining Anti-Mitochondrial Antibodies, Anti-Histone, and PLA2/COX Biomarkers to Increase Their Diagnostic Accuracy for Autism Spectrum Disorders



Abstract
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Blood Sampling
3.2. Ethics Approval and Consent
3.3. Biochemical Assays
3.3.1. Anti-Histone Antibodies Assay
3.3.2. Plasma AMA-M2 Assay
3.3.3. Assay of cPLA2
3.3.4. Assay of COX-2
3.4. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Limitations
References
- Sesay, M.M.; McCracken, C.E.; Stewart, C.; Simon, G.; Penfold, R.; Ahmedani, B.; Rossom, R.C.; Lu, C.Y.; Beck, A.; Coleman, K.J.; et al. Short report: Transition to International Classification of Diseases, 10th Revision and the prevalence of autism in a cohort of healthcare systems. Autism 2024, 19, 13623613231220687. [Google Scholar] [CrossRef]
- Arbuckle, M.R.; McClain, M.T.; Rubertone, M.V.; Scofield, R.H.; Dennis, G.J.; James, J.A.; Harley, J.B. Development of autoantibodies before the clinical onset of systemic lupus erythematosus. N. Engl. J. Med. 2003, 349, 1526–1533. [Google Scholar] [CrossRef]
- Colapietro, F.; Lleo, A.; Generali, E. Antimitochondrial Antibodies: From Bench to Bedside. Clin. Rev. Allergy Immunol. 2022, 63, 166–177. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism. Front. Physiol. 2014, 5, 150. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, X.; Xia, Y. Anti-double stranded DNA antibodies: Origin, pathogenicity, and targeted therapies. Front. Immunol. 2019, 10, 1667. [Google Scholar] [CrossRef]
- Rodríguez, C.G.; Hernández, M.A.; Torres, I.A. Update and clinical management of anti-DNA auto-antibodies. Adv. Lab. Med. 2021, 2, 313–321. [Google Scholar]
- Khan, M.A.; Alam, K.; Zafaryab, M.; Rizvi, M.M.A. Peroxynitrite-modified histone as a pathophysiological biomarker in autoimmune diseases. Biochimie 2017, 140, 1–9. [Google Scholar] [CrossRef]
- Lee, A.Y. Clinical use of anti-histone antibodies in idiopathic and drug-induced lupus. Immunol. Med. 2022, 45, 180–185. [Google Scholar] [CrossRef]
- Mecocci, P.; Ekman, R.; Parnetti, L.; Senin, U. Antihistone and anti-dsDNA autoantibodies in Alzheimer’s disease and vascular dementia. Biol. Psychiatry 1993, 34, 380–385. [Google Scholar] [CrossRef]
- Pereira, L.F.; Marco, F.M.; Boimorto, R.; Caturla, A.; Bustos, A.; Concha, E.D.L.; Subiza, J.L. Histones interact with anionic phospholipids with high avidity; its relevance for the binding of histone-antihistone immune complexes. Clin. Exp. Immunol. 1994, 97, 175–180. [Google Scholar] [CrossRef]
- D’Angelo, C.; Franch, O.; Fernández-Paredes, L.; Oreja-Guevara, C.; Núñez-Beltrán, M.; Comins-Boo, A.; Sánchez-Ramón, S. Antiphospholipid antibodies overlapping in isolated neurological syndrome and multiple sclerosis: Neurobiological insights and diagnostic challenges. Front. Cell. Neurosci. 2019, 107, 425124. [Google Scholar] [CrossRef]
- Premrajan, P.; Jayanandan, A.; Variyar, E.J. Chapter 8—PLA2: Implications in neurological disorders as a prospective therapeutic target. In Phospholipases in Physiology and Pathology; Chakraborti, S., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 139–158. [Google Scholar]
- Kaufmann, W.E.; Worley, P.F.; Pegg, J.; Bremer, M.; Isakson, P. COX-2, a synaptically induced enzyme, is expressed by excitatory neurons at postsynaptic sites in rat cerebral cortex. Proc. Natl. Acad. Sci. USA 1996, 93, 2317–2321. [Google Scholar] [CrossRef]
- Yagami, T.; Koma, H.; Yamamoto, Y. Pathophysiological Roles of Cyclooxygenases and Prostaglandins in the Central Nervous System. Mol. Neurobiol. 2016, 53, 4754–4771. [Google Scholar] [CrossRef]
- Chen, C.; Magee, J.C.; Bazan, N.G. Cyclooxygenase-2 regulates prostaglandin E2 signaling in hippocampal long-term synaptic plasticity. J. Neurophysiol. 2002, 87, 2851–2857. [Google Scholar] [CrossRef]
- Yang, H.; Chen, C. Cyclooxygenase-2 in synaptic signaling. Curr. Pharm. Des. 2008, 14, 1443–1451. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Receiver operating characteristic (ROC) curve analysis for medical diagnostic test evaluation. Casp. J. Intern. Med. 2013, 4, 627–635. [Google Scholar]
- Hassan, W.M.; Al-Ayadhi, L.; Bjørklund, G.; Alabdali, A.; Chirumbolo, S.; El-Ansary, A. The Use of Multi-parametric Biomarker Profiles May Increase the Accuracy of ASD Prediction. J. Mol. Neurosci. 2018, 66, 85–101. [Google Scholar] [CrossRef]
- Hassan, W.M.; Al-Dbass, A.; Al-Ayadhi, L.; Bhat, R.S.; El-Ansary, A. Discriminant analysis and binary logistic regression enable more accurate prediction of autism spectrum disorder than principal component analysis. Sci. Rep. 2022, 12, 3764. [Google Scholar] [CrossRef]
- Zhou, X.; McClish, D.; Obuchowski, N. Statistical Methods in Diagnostic Medicine; John Wiley & Sons: New York, NY, USA, 2002. [Google Scholar]
- Pepe, M. The Statistical Evaluation of Medical Tests for Classification and Prediction; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Bell, J.G.; Sargent, J.R.; Tocher, D.R.; Dick, J.R. Red blood cell fatty acid compositions in a patient with autistic spectrum disorder: A characteristic abnormality in neurodevelopmental disorders. Prostaglandins Leukot. Essent. Fat. Acids 2000, 63, 21–25. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.G.; Al-Ayahdi, L.Y. Plasma fatty acids as diagnostic markers in autistic patients from Saudi Arabia. Lipids Health Dis. 2011, 10, 62. [Google Scholar] [CrossRef]
- Bell, J.G.; MacKinlay, E.E.; Dick, J.R.; MacDonald, D.J.; Boyle, R.M.; Glen, A.C. Essential fatty acids and phospholipase A2 in autistic spectrum disorders. Prostaglandins Leukot. Essent. Fat. Acids 2004, 71, 201–204. [Google Scholar] [CrossRef]
- Qasem, H.; Al-Ayadhi, L.; Al Dera, H.; El-Ansary, A. Increase of cytosolic phospholipase A2 as hydrolytic enzyme of phospholipids and autism cognitive, social and sensory dysfunction severity. Lipids Health Dis. 2017, 16, 117. [Google Scholar] [CrossRef]
- Tostes, M.H.; Teixeira, H.C.; Gattaz, W.F.; Brandão, M.A.; Raposo, N.R. Altered neurotrophin, neuropeptide, cytokines and nitric oxide levels in autism. Pharmacopsychiatry 2012, 45, 241–243. [Google Scholar] [CrossRef]
- Liu, X.; Lin, J.; Zhang, H.; Khan, N.U.; Zhang, J.; Tang, X.; Cao, X.; Shen, L. Oxidative Stress in Autism Spectrum Disorder—Current Progress of Mechanisms and Biomarkers. Front. Psychiatry 2022, 13, 813304. [Google Scholar] [CrossRef]
- Su, K.P.; Huang, S.Y.; Peng, C.Y.; Lai, H.C.; Huang, C.L.; Chen, Y.C.; Aitchison, K.J.; Pariante, C.M. Phospholipase A2 and cyclooxygenase 2 genes influence the risk of interferon-alpha-induced depression by regulating polyunsaturated fatty acids levels. Biol. Psychiatry 2010, 67, 550–557. [Google Scholar] [CrossRef]
- Siddiqui, M.F.; Elwell, C.; Johnson, M.H. Mitochondrial Dysfunction in Autism Spectrum Disorders. Autism Open Access 2016, 6, 1000190. [Google Scholar] [CrossRef]
- Prata, J.; Machado, A.S.; von Doellinger, O.; Almeida, M.I.; Barbosa, M.A.; Coelho, R.; Santos, S.G. The Contribution of Inflammation to Autism Spectrum Disorders: Recent Clinical Evidence. Methods Mol. Biol. 2019, 2011, 493–510. [Google Scholar]
- Pecorelli, A.; Ferrara, F.; Messano, N.; Cordone, V.; Schiavone, M.L.; Cervellati, F.; Woodby, B.; Cervellati, C.; Hayek, J.; Valacchi, G. Alterations of mitochondrial bioenergetics, dynamics, and morphology support the theory of oxidative damage involvement in autism spectrum disorder. FASEB J. 2020, 34, 6521–6538. [Google Scholar] [CrossRef]
- Gu, F.; Chauhan, V.; Kaur, K.; Brown, W.T.; LaFauci, G.; Wegiel, J.; Chauhan, A. Alterations in mitochondrial DNA copy number and the activities of electron transport chain complexes and pyruvate dehydrogenase in the frontal cortex from subjects with autism. Transl. Psychiatry 2013, 3, e299. [Google Scholar] [CrossRef]
- Khemakhem, A.M.; Frye, R.E.; El-Ansary, A.; Al-Ayadhi, L.; Bacha, A.B. Novel biomarkers of metabolic dysfunction is autism spectrum disorder: Potential for biological diagnostic markers. Metab. Brain Dis. 2017, 32, 1983–1997. [Google Scholar] [CrossRef]
- El-Ansary, A.; Bjørklund, G.; Khemakhem, A.M.; Al-Ayadhi, L.; Chirumbolo, S.; Ben Bacha, A. Metabolism-associated markers and childhood autism rating scales (CARS) as a measure of autism severity. J. Mol. Neurosci. 2018, 65, 265–276. [Google Scholar] [CrossRef]
- Cascone, A.; Bruelle, C.; Lindholm, D.; Bernardi, P.; Eriksson, O. Destabilization of the Outer and Inner Mitochondrial Membranes by Core and Linker Histones. PLoS ONE 2012, 7, e35357. [Google Scholar] [CrossRef]
- Hughes, H.K.; Mills Ko, E.; Rose, D.; Ashwood, P. Immune Dysfunction and Autoimmunity as Pathological Mechanisms in Autism Spectrum Disorders. Front Cell. Neurosci. 2018, 12, 405. [Google Scholar] [CrossRef]
- Kolevzon, A.; Gross, R.; Reichenberg, A. Prenatal and perinatal risk factors for autism: A review and integration of findings. Arch. Pediatr. Adolesc. Med. 2007, 161, 326–333. [Google Scholar] [CrossRef]
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental risk factors for autism: An evidence-based review of systematic reviews and meta-analyses. Mol. Autism 2017, 8, 13. [Google Scholar] [CrossRef]
- Wilson, E.N.; Mabry, S.; Bradshaw, J.L.; Gardner, J.J.; Rybalchenko, N.; Engelland, R.; Cunningham, R.L. Gestational hypoxia in late pregnancy differentially programs subcortical brain maturation in male and female rat offspring. Biol. Sex Differ. 2022, 13, 54. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | AUC | Cut-Off Value | Sensitivity % | Specificity % | p Value | 95% CI |
|---|---|---|---|---|---|---|
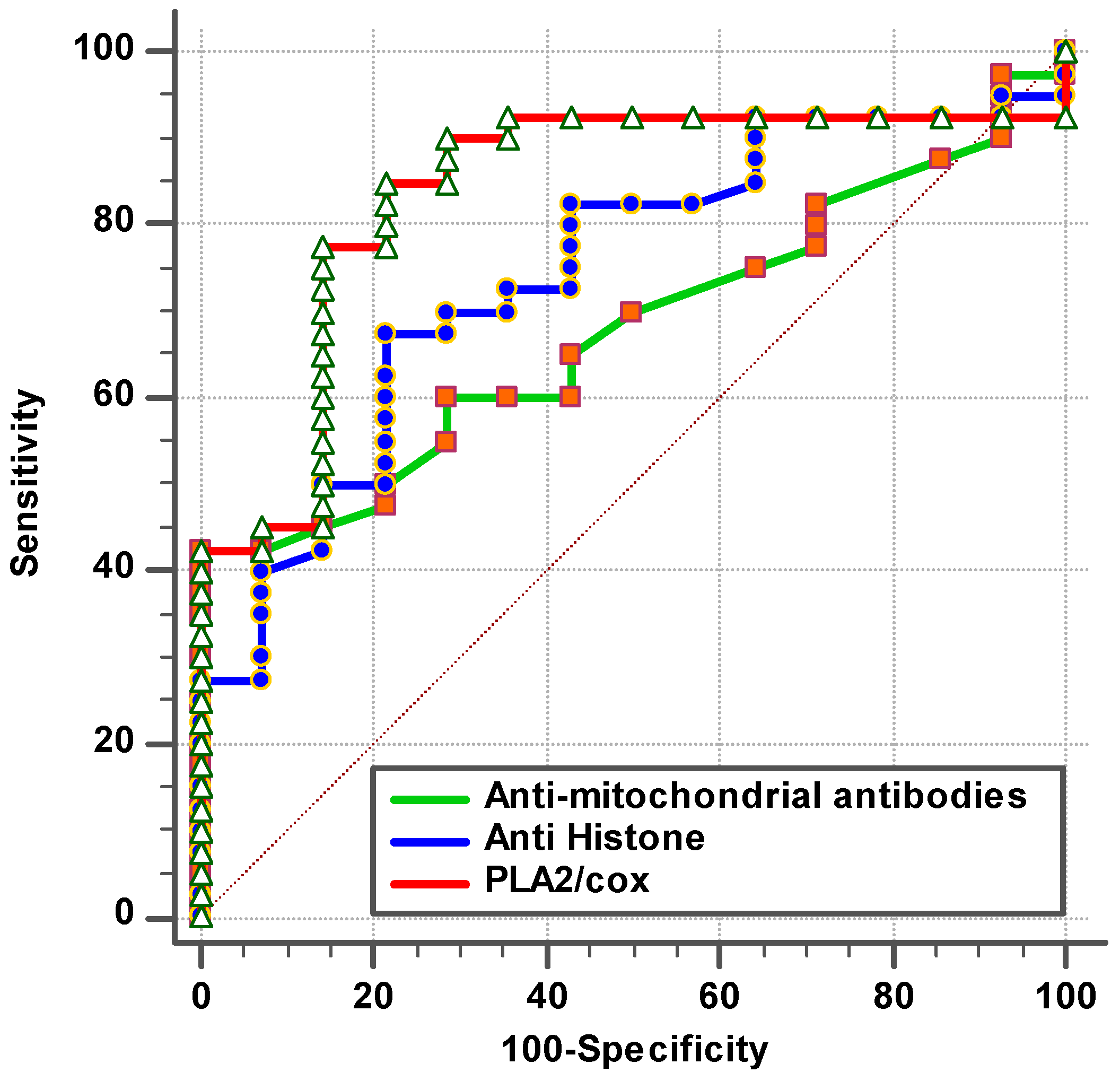
| Anti-mitochondrial antibodies (AMA-M2) | 0.663 | 0.162 | 40.7% | 100.0% | 0.062 | 0.530–0.796 |
| Anti-histone antibodies | 0.770 | 0.672 | 71.1% | 75.0% | 0.001 | 0.653–0.887 |
| PLA2/COX | 0.869 | 0.205 | 85.0% | 85.0% | 0.001 | 0.780–0.959 |
| Parameters | AUC | Sensitivity % | Specificity % | p Value | 95% CI |
|---|---|---|---|---|---|
| AMA-M2 with anti-histone antibodies | 0.743 | 40.0% | 100.0% | 0.006 | 0.601–0.885 |
| AMA-M2 with PLA2/COX | 0.871 | 87.5% | 78.6% | 0.000 | 0.774–0.969 |
| Anti-histone antibodies with PLA2/COX | 0.909 | 90.0% | 75.0% | 0.000 | 0.834–0.984 |
| AMA-M2 with anti-histone antibodies with PLA2/COX | 0.941 | 95.0% | 78.6% | 0.000 | 0.881–1.000 |
| Parameters | Groups | N | Min. | Max. | Mean ± S.D. | Median | Percent Change | p Value |
|---|---|---|---|---|---|---|---|---|
| Anti-mitochondrial antibodies (AMA-M2) | Control | 14 | 0.08 | 0.11 | 0.10 ± 0.01 | 0.10 | 100.00 | 0.050 |
| Patient | 54 | 0.08 | 0.33 | 0.22 ± 0.03 | 0.10 | 227.13 | ||
| Anti-histone antibodies | Control | 20 | 0.25 | 7.70 | 1.82 ± 1.84 | 1.15 | 100.00 | 0.001 |
| Patient | 45 | 0.05 | 8.13 | 0.87 ± 1.47 | 0.37 | 47.71 | ||
| PLA2/COX-2 | Control | 40 | 0.00 | 0.56 | 0.11 ± 0.13 | 0.07 | 100.00 | 0.001 |
| Patient | 40 | 0.00 | 3.07 | 0.62 ± 0.58 | 0.44 | 537.34 |
| Parameters | R (Correlation Coefficient) | p Value | |
|---|---|---|---|
| Anti-mitochondrial antibodies (AMA-M2) with anti-histone | 0.043 | 0.746 | P a |
| Anti-mitochondrial antibodies (AMA-M2) with PLA2/COX-2 | 0.087 | 0.533 | P a |
| Anti-histone with PLA2/COX-2 | −0.013 | 0.921 | N b |
| Parameters | Regression Coefficient | Standard Error | Odds Ratio | 95% CI for Odds Ratio | p Value | |
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| Anti-mitochondrial antibodies (AMA-M2) | 55.540 | 25.335 | 1.32 ×1024 | 359.336 | 4.85 × 1045 | 0.028 |
| Anti-histone | −0.501 | 0.237 | 0.606 | 0.381 | 0.965 | 0.035 |
| Anti-mitochondrial antibodies (AMA-M2) | 63.070 | 34.255 | 2.46 × 1027 | 0.017 | 3.54 × 1056 | 0.066 |
| PLA2/COX-2 | 5.823 | 2.163 | 337.866 | 4.868 | 23,449.68 | 0.007 |
| Anti-histone | −1.090 | 0.378 | 0.336 | 0.160 | 0.706 | 0.004 |
| PLA2/COX-2 | 8.819 | 2.653 | 6.76 × 103 | 37.325 | 1.22 × 106 | 0.001 |
| Anti-mitochondrial antibodies (AMA-M2) | 76.848 | 39.487 | 2.37 × 1033 | 0.580 | 9.69 × 1066 | 0.052 |
| Anti-histone antibodies | −1.624 | 0.583 | 0.197 | 0.063 | 0.618 | 0.005 |
| PLA2/COX-2 | 10.503 | 3.702 | 3.64 × 104 | 25.726 | 5.15 × 107 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Ansary, A.; Alfawaz, H.A.; Bacha, A.B.; Al-Ayadhi, L.Y. Combining Anti-Mitochondrial Antibodies, Anti-Histone, and PLA2/COX Biomarkers to Increase Their Diagnostic Accuracy for Autism Spectrum Disorders. Brain Sci. 2024, 14, 576. https://doi.org/10.3390/brainsci14060576
El-Ansary A, Alfawaz HA, Bacha AB, Al-Ayadhi LY. Combining Anti-Mitochondrial Antibodies, Anti-Histone, and PLA2/COX Biomarkers to Increase Their Diagnostic Accuracy for Autism Spectrum Disorders. Brain Sciences. 2024; 14(6):576. https://doi.org/10.3390/brainsci14060576
Chicago/Turabian StyleEl-Ansary, Afaf, Hanan A. Alfawaz, Abir Ben Bacha, and Laila Y. Al-Ayadhi. 2024. "Combining Anti-Mitochondrial Antibodies, Anti-Histone, and PLA2/COX Biomarkers to Increase Their Diagnostic Accuracy for Autism Spectrum Disorders" Brain Sciences 14, no. 6: 576. https://doi.org/10.3390/brainsci14060576
APA StyleEl-Ansary, A., Alfawaz, H. A., Bacha, A. B., & Al-Ayadhi, L. Y. (2024). Combining Anti-Mitochondrial Antibodies, Anti-Histone, and PLA2/COX Biomarkers to Increase Their Diagnostic Accuracy for Autism Spectrum Disorders. Brain Sciences, 14(6), 576. https://doi.org/10.3390/brainsci14060576