1. Introduction
It is well known that, as one of the important nutrients of aquatic animals, dietary lipids not only provide energy for growth and development in fish, but also provide the essential fatty-acid and fat-soluble vitamins that maintain normal structure and biological function [
1]. At present, the main lipid sources in aquatic feed are fish oil and soybean oil. Compared with soybean oil, fish oil has a high content of unsaturated fatty acids (HUFAs), such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), and a good feeding attraction effect, so it is the best lipid source for aquatic animals [
2,
3]. However, EPA and DHA are easily oxidized during the storage and processing of fish oil and feed, producing harmful substances such as lipid hydroperoxides, ketones, aldehydes, and acids [
4,
5]. Studies have also reported that feeding an oxidized-fish-oil diet has been found to decrease growth performance and cause oxidative stress of tilapia (
Oreochromis niloticus) [
6], Wuchang bream (
Megalobrama amblycephala) [
7], and orange spotted grouper (
Epinephelus coioides) [
5], disrupt lipid metabolism of
Rhynchocypris lagowski Dybowski [
8], and induce intestinal injury of
Megalobrama amblycephala [
9] and rice field eel (
Monopterus albus) [
10]. Therefore, exploring effective dietary strategies is imperative to alleviate the negative effects of oxidized-fish-oil diets on aquatic animals.
Taurine is a type of non-protein amino acid in the form of a free amino acid, which has a wide range of physiological functions, such as calcium homeostasis, osmotic regulation, membrane stability, and antioxidant and anti-inflammatory functions [
11,
12]. Many studies have also demonstrated the versatility of taurine in aquatic animals. Previous studies in our laboratory indicated that taurine supplementation in a low-fish-meal diet increased growth performance and immunity function and enhanced anti-stress ability in black carp (
Mylopharyngodon piceus) [
13] and rice field eel [
14]. Similar studies have found, in other aquatic animals, that taurine can increase growth performance, enhance antioxidant ability, improve intestinal health, and reduce lipid deposition of seabass (
Dicentrarchus labrax) [
15], grass carp [
16], and California yellowtail (
Seriola dorsalis) [
17]. However, there is no report on whether taurine can alleviate the negative effects caused by oxidized-fish-oil diets in aquatic animals. Therefore, we have carried out related research.
Channel catfish (
Ictalurus punctatus), which belongs to catfish family (
Siluriformes), is an important freshwater aquaculture fish in China. Because of its delicious meat, high nutritional value, and fast growth, it is welcomed by producers and consumers. In 2018, the production of channel catfish exceeded 390,000 tons in the world, an increase of 3.34% over the previous year [
18]. Channel catfish has a high requirement for feed freshness, and feed mildew, deterioration, and oxidation will have a negative impact on growth and health. Therefore, this study aimed to investigate whether taurine can alleviate the negative effects of lipid deposition, oxidative stress, and intestinal damage induced by oxidized-fish-oil diets in juvenile channel catfish. It is of great significance to explore the side-effects of oxidized fat ingestion on the growth and health of aquatic animals for the study of fish nutrition and health, and to provide solutions for practical production and a theoretical basis for the application of taurine.
4. Discussion
A fresh fish oil can provide the HUFAs needed during the growth of fish [
23]. HUFAs are prone to oxidative rancidity and have a negative impact on fish. In this study, oxidized-fish-oil diets significantly reduced the growth performance of channel catfish, which is similar to results obtained in juvenile hybrid grouper (♀
Epinephelus fuscoguttatus × ♂
Epinephelus lanceolatus) [
24], farmed tilapia [
6], orange spotted grouper [
5], and yellow catfish (
Pelteobagrus fulvidraco) [
25]. One of the reasons is that toxic and harmful substances such as lipid hydroperoxides, ketones, aldehydes, and acids are produced after oxidation of fish oil, which induces oxidative stress, leads to inflammatory response, and then inhibits growth [
5]. Another reason is that oxidized fish oil has reduced nutritional value compared with non-oxidized fish oil [
26]. Taurine has been widely used in aquatic feeds. Adding an appropriate amount of taurine to diets can obviously increase growth performance of yellowtail kingfish (
Seriola lalandi) [
27], turbot [
28], and tiger puffer (
Takifugu rubripes) [
29]. Experimental results also showed that the addition of 0.2% taurine to the oxidized-fish-oil diet obviously increased the growth performance of channel catfish, and there was no significant difference from the FFO group. There are two main reasons why taurine promotes fish growth: first, taurine has a good attractant effect [
30]; second, taurine may alleviate the negative effects caused by oxidized-fish-oil diets, such as lipid deposition, oxidative damage, and inflammatory response.
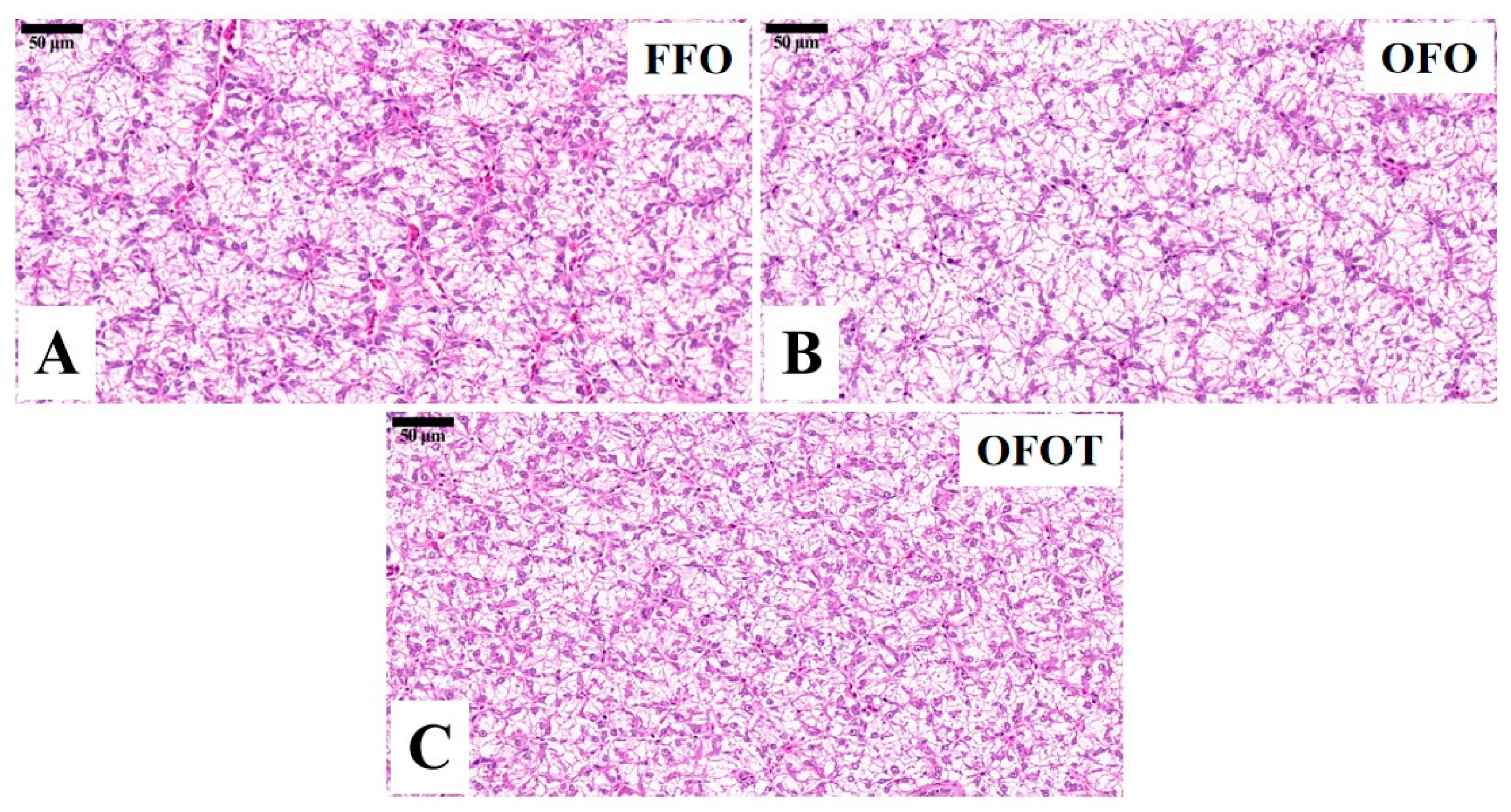
Long-term feeding of oxidized-fish-oil diets can lead to the lipid deposition of liver [
31]. Based on H&E staining, liver fat vacuolation is usually expressed as the size of hepatocytes and their nuclei [
32,
33]. In this study, oxidized-fish-oil diets led to lipid deposition in the liver of channel catfish, which was supported by the phenomenon of increased lipid vacuolization in the liver (such as smaller nuclei and larger hepatocytes), HSI, serum TC, and TG contents. Similar studies have been found in yellow catfish [
25], loach (
Misgurnus anguillicaudatus) [
34] and largemouth bass (
Micropterus salmoides) [
35]. Further studies showed that oxidized-fish-oil diets resulted in liver lipid deposition due to the upregulation of the expression of lipid synthesis gene (
fas) and the downregulation of the expression of lipolysis gene (
lpl). Sterol-regulatory element binding protein 1 (
srebp1) is mainly involved in the activation of enzymes related to lipid synthesis, and can promote lipid synthesis by targeting the expression of fatty-acid-catalyzing enzymes such as
fas [
36,
37]. The present study showed that feeding oxidized-fish-oil diets significantly upregulated
srebp1 mRNA expression, indicating that oxidized-fish-oil diets can induce lipid deposition by regulating the mRNA expression of lipid synthesis and lipolysis. Taurine has a good function of reducing lipid deposition. It has been found in broiler chickens that taurine can reduce blood lipid content [
38]. There are also studies in aquatic animals that have found that taurine can promote lipolysis of European seabass [
15], white seabream (
Diplodus sargus) [
39], and Persian sturgeon (
Acipenser persicus) [
40]. In this study, the addition of taurine to oxidized-fish-oil diets remarkably reduced lipid vacuolization in the liver, HSI, serum TC, and TG contents. Furthermore, taurine remarkably downregulated the transcriptional levels of
fas and
srebp1 in the liver, and upregulated the transcriptional level of
lpl, indicating that taurine alleviated lipid deposition induced by oxidized-fish-oil diets. Studies have speculated that taurine has a good lipolysis effect, which may be related to the AMPK/SIRT1 signaling pathway [
38]. Previous studies confirmed that activation of AMPK can inhibit the activities of FAS and ACC, thereby reducing the concentration of malonyl-CoA and enhancing CPT1 activity, thus increasing lipid catabolism and reducing lipid deposition [
41,
42]. However, the mechanism of taurine alleviating lipid deposition needs further study.
For animals, the oxidation diet is one of important exogenous factors leading to oxidative stress. Long-term feeding of oxidized-fish-oil diets can induce the production of reactive oxygen species in mitochondria, and excessive reactive oxygen species (ROS) can lead to tissue oxidative damage [
43,
44,
45]. Malondialdehyde (MDA) is the final decomposition product of lipid peroxidation caused by ROS, and its content reflects the degree of peroxidation [
46]. In the process of ROS removal, CAT breaks down hydrogen peroxide into oxygen and water, and SOD and GPx also play an important role, which can decrease hydrogen peroxide [
47,
48]. The present study showed that oxidized-fish-oil-diet feeding led to markedly a higher the content of MDA and lower the levels of CAT, SOD, GPx, GR, and T-AOC in the liver than in the FFO group. As is well known, the increase of serum AST and ALT activities is one of the important markers of liver injury [
49]. Besides, the present study has showed that oxidized-fish-oil diets significantly increased serum AST and ALT activities of channel catfish, indicating that oxidized-fish-oil diets leads to oxidative stress and damage in the liver. Similar studies have found in other aquatic animals that oxidized-fish-oil diets significantly decreased antioxidant enzyme activities and increased AST and ALT activities of Wuchang bream [
7] and tilapia (
Oreochromis niloticus) [
6]. Some studies have shown that taurine is a powerful antioxidant, mainly due to its stable biofilm and direct scavenging ability of ROS [
50]. Furthermore, taurine can also improve antioxidant capacity by increasing the activity of antioxidant enzymes [
51]. The results of this experiment also showed that the addition of taurine to the oxidized-fish-oil diet remarkably promoted CAT, GPx, GR, SOD, and T-AOC levels, whereas the opposite result was observed for the MDA level. The antioxidant capacity of taurine is related to its role as a precursor of glutathione [
52], and taurine can also enhance the regeneration of glutathione from glutathione disulfide [
53].
Antioxidant enzyme activity is regulated by the nrf2/keap1 signaling pathway [
54].
keap1 inhibits the expression of antioxidant genes by inhibiting the nuclear translocation of
nrf2 [
55]. The present study showed that oxidized-fish-oil diets remarkably downregulated the transcriptional levels of
nrf2,
gr, and
gpx1 in the liver, while the transcriptional levels of
keap1 were reversed. These results were consistent with the results of antioxidant enzyme activities, indicating that long-term feeding of oxidized-fish-oil diets can reduce the antioxidant capacity of channel catfish. Previous studies in pufferfish (
Takifugu obscurus) [
56] and yellow catfish (
Pelteobagrus fulvidraco) [
57] have found that when fish are under oxidative stress, dietary taurine can increase the expression levels of antioxidant enzyme genes in the liver, thus improving antioxidant capacity. In this study, the addition of taurine to oxidized-fish-oil diets remarkably upregulated
nrf2,
gr, and
gpx1 mRNA expression, whereas the opposite result was observed for the transcriptional level of
keap1. These results were consistent with the results of antioxidant enzyme activities, indicating that taurine can relieve peroxidation injury of channel catfish caused by oxidized-fish-oil diets. Similar studies have confirmed that taurine can remarkably improve antioxidant ability in juvenile turbot by regulating the
nrf2/
keap1 signaling pathway [
58]. Therefore, we speculate that taurine regulates the activity of antioxidant enzymes through the
nrf2/
keap1 signaling pathway, thereby enhancing the ability of fish to resist oxidative stress.
Immune-active substances such as immunoglobulin and complement factor in serum play an important role in animal immune response [
59]. Fish mainly rely on the non-specific immune system to respond to external environmental stimuli and pathogen invasion [
60]. As a protein response system, the complement system is mainly responsible for destroying or removing pathogenic microorganisms, and is an important part of the non-specific immunity [
61]. The present study showed that oxidized-fish-oil diets significantly decreased immune function, which was supported by the phenomenon of decreased serum C3, C4, and IgM contents. Some studies have indicated that dietary supplementation of taurine can alleviate acute ammonia poisoning of yellow catfish by increasing the content of total immunoglobulin in serum [
57]. In addition, our previous study revealed that taurine can improve serum C3 and C4 levels in rice field eel to alleviate the immune response induced by high-fat diets [
62]. The results of this study showed that the dietary supplementation of taurine to a oxidized-fish-oil diet increased serum C3, C4, and IgM contents, which indicated that taurine can improve immune function of channel catfish. Similar experimental results were found in Chinese mitten crab (
Eriocheir sinensis) [
63] and yellow catfish [
64].
Further research has shown that taurine can enhance immune function though controlling intestinal inflammatory response [
65]. Intestinal inflammatory response is mainly regulated by cytokines, including anti-inflammatory cytokines (including
tgf-β and
il-10) and pro-inflammatory cytokines (including
tnf-α il-1β,
il-6, and
il-8) [
66]. The present study showed that the mRNA transcription levels of
tnf-α il-1β,
il-6, and
il-8 were remarkably upregulated when channel catfish fed oxidized-fish-oil diets, whereas the opposite result was observed for the mRNA transcription levels of
tgf-β1,
tgf-β2, and
tgf-β3. A similar study has been conducted in
Rhynchocypris lagowski, which showed that oxidized-fish-oil diets lead to high expression of pro-inflammatory cytokines (
tnf-α,
il-1β, and
il-8) and low expression of anti-inflammatory cytokines (
il-10 and
tgf-β) [
67]. Finding how to alleviate the inflammatory reaction caused by an oxidized-fish-oil diet is very important to improving the utilization rate of aquatic feed. Previous studies have reported that adding taurine significantly downregulated the expression levels of anti-inflammatory cytokines in grass carp [
16] and yellow catfish [
57]. The results of this experiment also shown that the addition of taurine to a oxidized-fish-oil diet dramatically downregulated
tnf-α il-1β,
il-6, and
il-8 mRNA expression in the liver, and upregulated
tgf-β1,
tgf-β2, and
tgf-β3 mRNA expression, indicating that taurine can reduce inflammatory response in the intestine induced by oxidized-fish-oil diets.
Cytokine expression in inflammatory response is regulated by various signaling pathways, among which nuclear transcription factor-κB (NF-κB), as an important signaling factor, plays an important role in inflammatory response [
68]. In this study, long-term feeding of oxidized-fish-oil diets remarkably upregulated the transcriptional level of
nf-κb. However, the addition of taurine to oxidized-fish-oil diets reversed this trend. Furthermore, correlation analyses showed that the mRNA expression level of
nf-κb was negatively correlated with the mRNA expression levels of
tnf-α,
il-1β,
il-6, and
il-8, and positively correlated with the mRNA expression levels of
tgf-β1,
tgf-β2, and
tgf-β3, which suggested that taurine inhibited the NF-κB signaling pathway to protecting oxidized fish-oil-induced inflammation response in channel catfish.
Intestinal physical-barrier function is an indispensable part of intestinal health of aquatic animals [
69]. Generally speaking, the muscular thickness and villi length in the intestine are important criteria to measure the efficiency of digestion and absorption [
70]. Goblet cells on intestinal villi, as typical mucous cells, play an important role in regulating the integrity of intestinal epithelial cells and the immune response to foreign antigens [
71]. The present study showed that oxidized-fish-oil diets significantly reduced the villi length, goblet cell quantity, and muscular thickness of intestine. However, the addition of taurine to oxidized-fish-oil diets reversed this trend, indicating taurine can maintain the structural integrity of the intestine. An important component of the intestinal physical barrier is tight junction protein. Studies have reported that tight junction proteins are closely related to the integrity of intestinal structure, and the upregulation of transmembrane protein-related genes (including
occluding,
zo-1, and
zo-2) can maintain the structural integrity of intestinal epithelial cells, while the upregulation of cytoplasmic protein-related genes (including
claudin-12 and
claudin-15) can damage the structural integrity of intestinal epithelial cells [
54,
72]. This present study found that oxidized-fish-oil diets substantially downregulated intestinal
zo-1 and
zo-2 mRNA transcriptional levels of channel catfish, and upregulated
claudin-12 and
claudin-15 mRNA transcriptional levels, indicating that oxidized-fish-oil diets may increase the intestinal barrier structure damage caused by intercellular space by regulating tight junction protein genes. Taurine has been reported to enhance intestinal morphology and barrier function [
73]. The results of this experiment also showed that the addition of taurine to a oxidized-fish-oil diet remarkably upregulated intestinal
zo-1 and
zo-2 mRNA transcriptional levels, and downregulated
claudin-12 and
claudin-15 mRNA transcriptional levels, indicating that taurine can repair the intestinal physical barrier damage induced by oxidized-fish-oil diets. However, the specific regulatory mechanism needs to be further studied.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}