
Cations and Phenolic Compounds Concentrations in Fruits of Fig Plants Exposed to Moderate Levels of Salinity

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Salt Treatment
2.2. Cation Concentration
2.3. Proline Concentration
2.4. Fruit Phenolic Compounds
2.5. Statistical Analysis
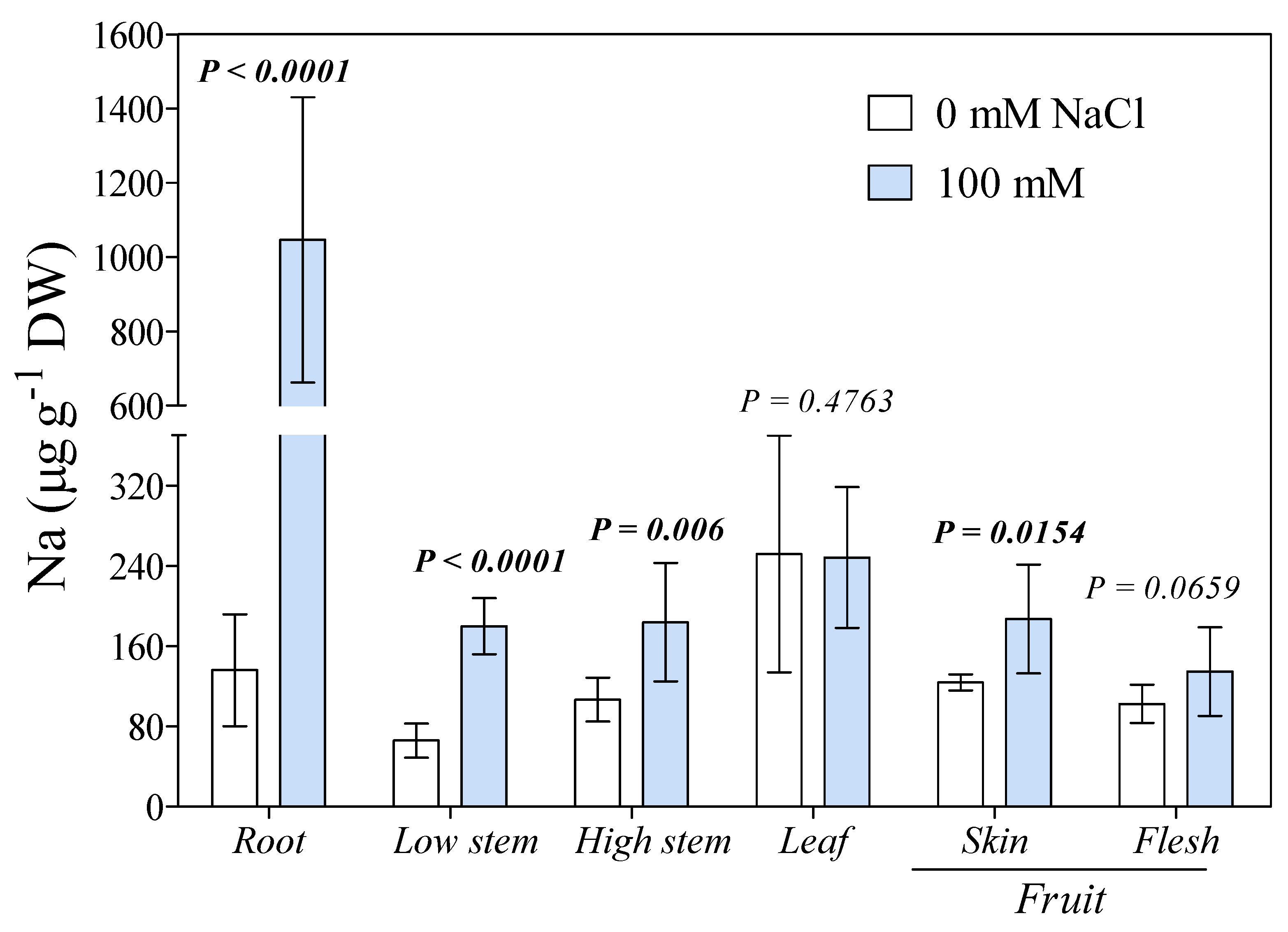
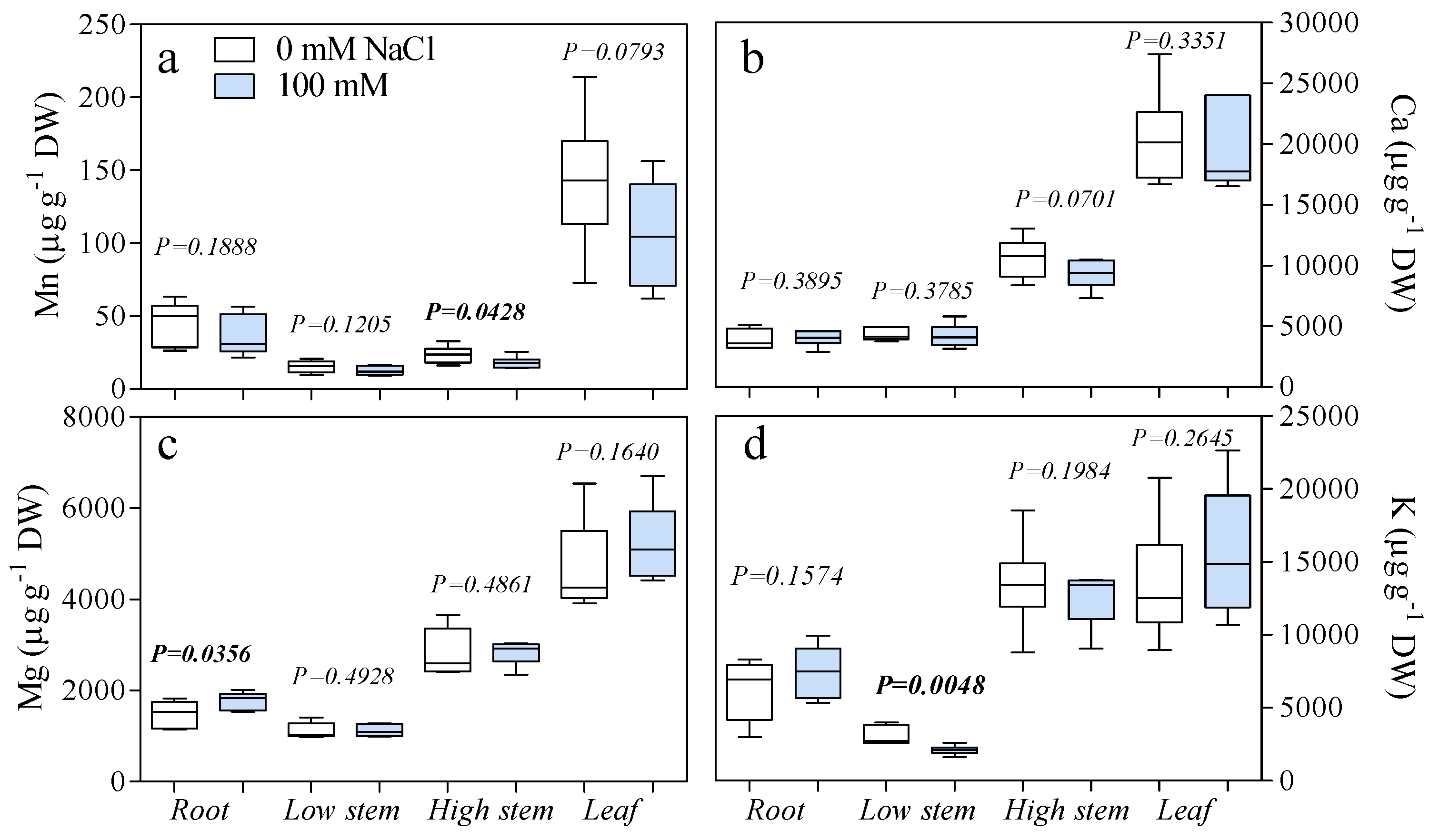
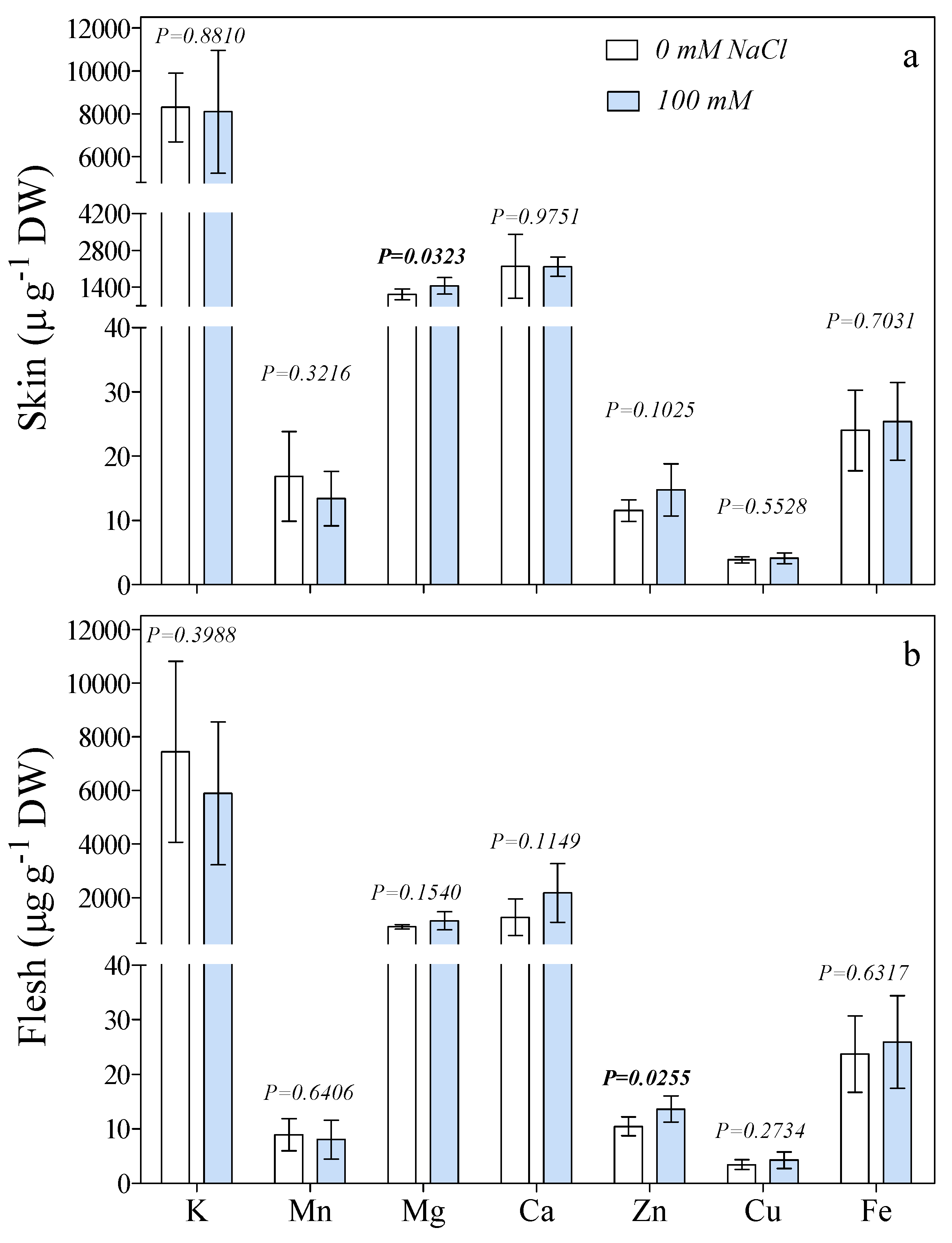
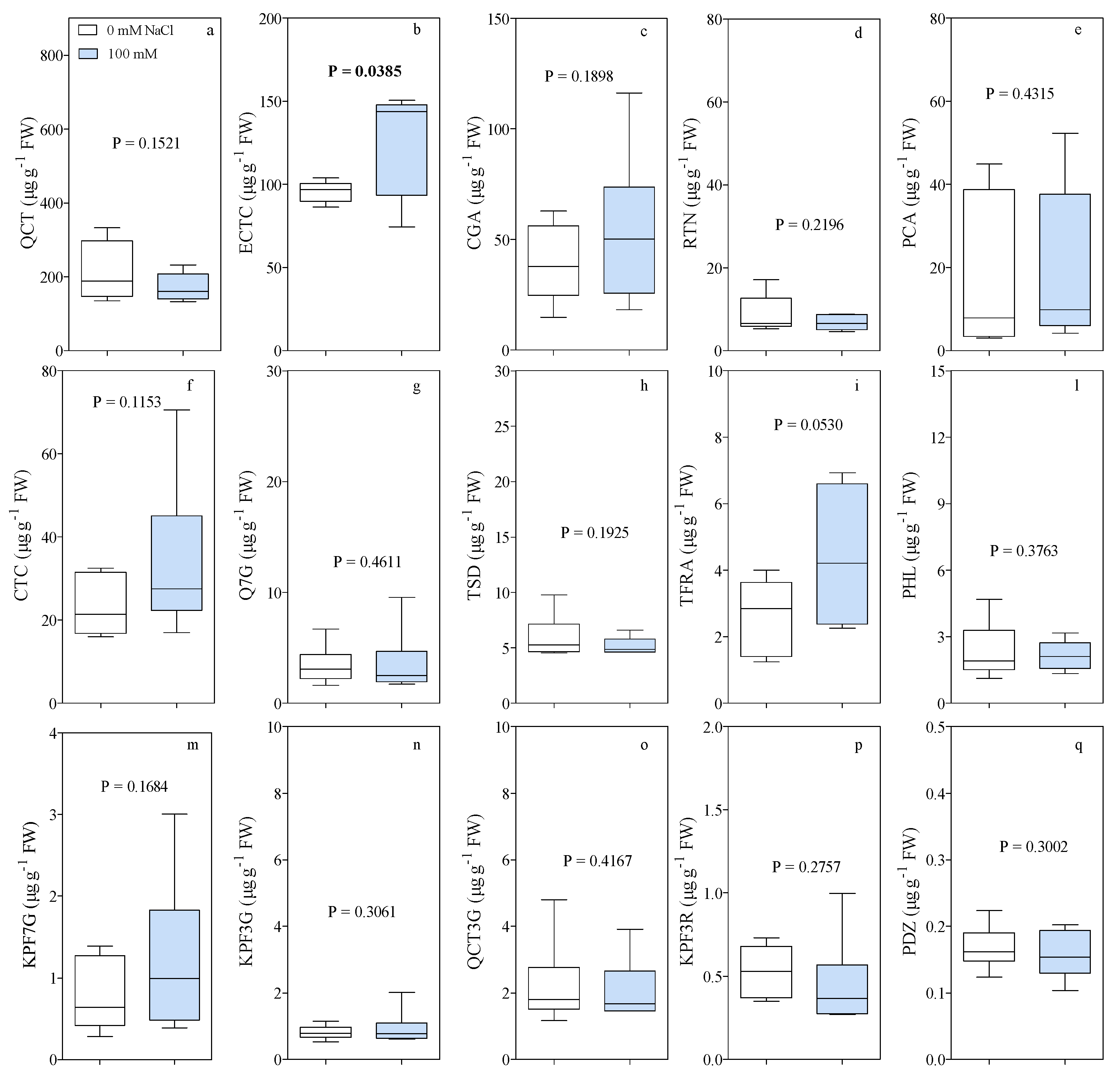
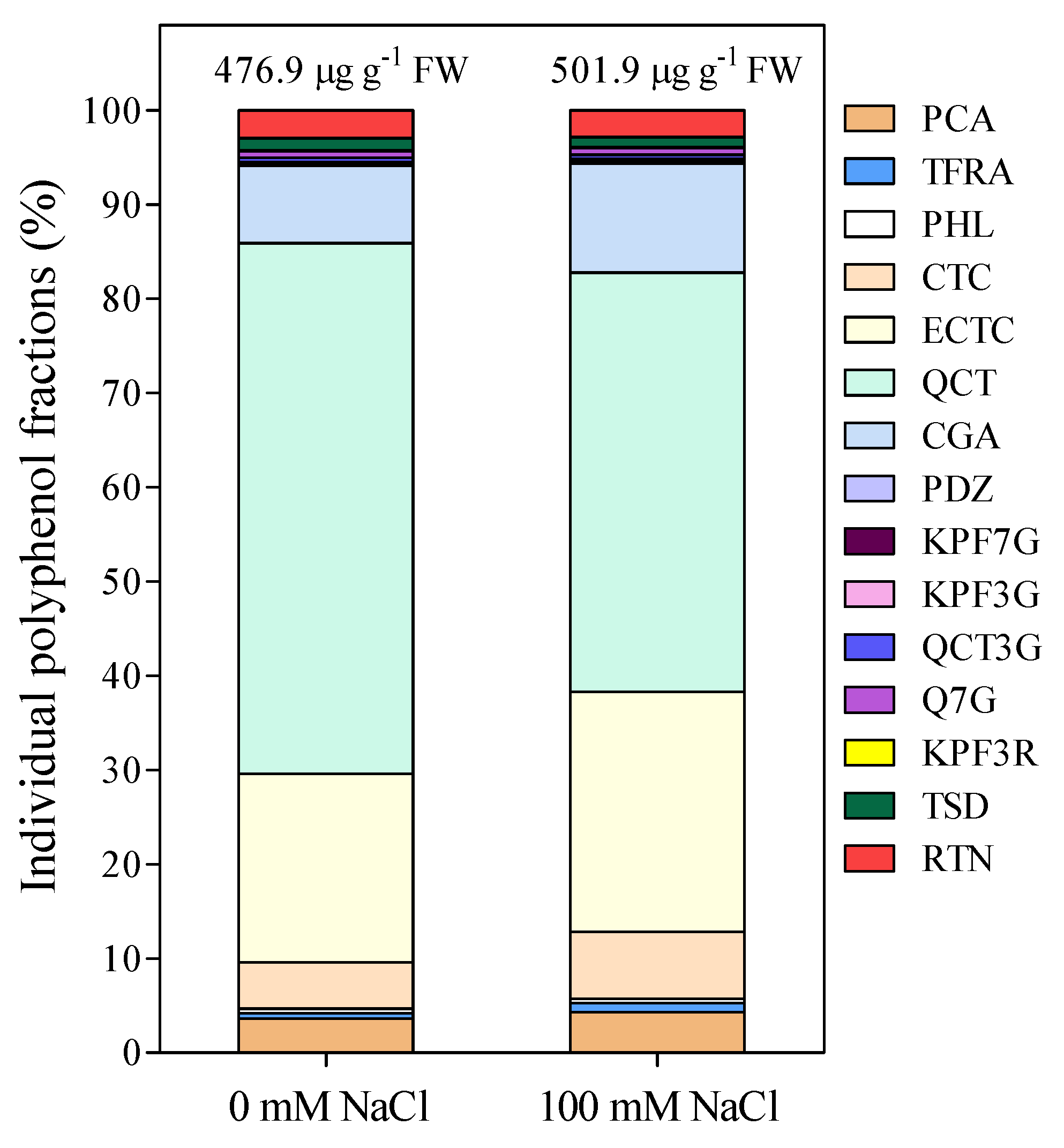
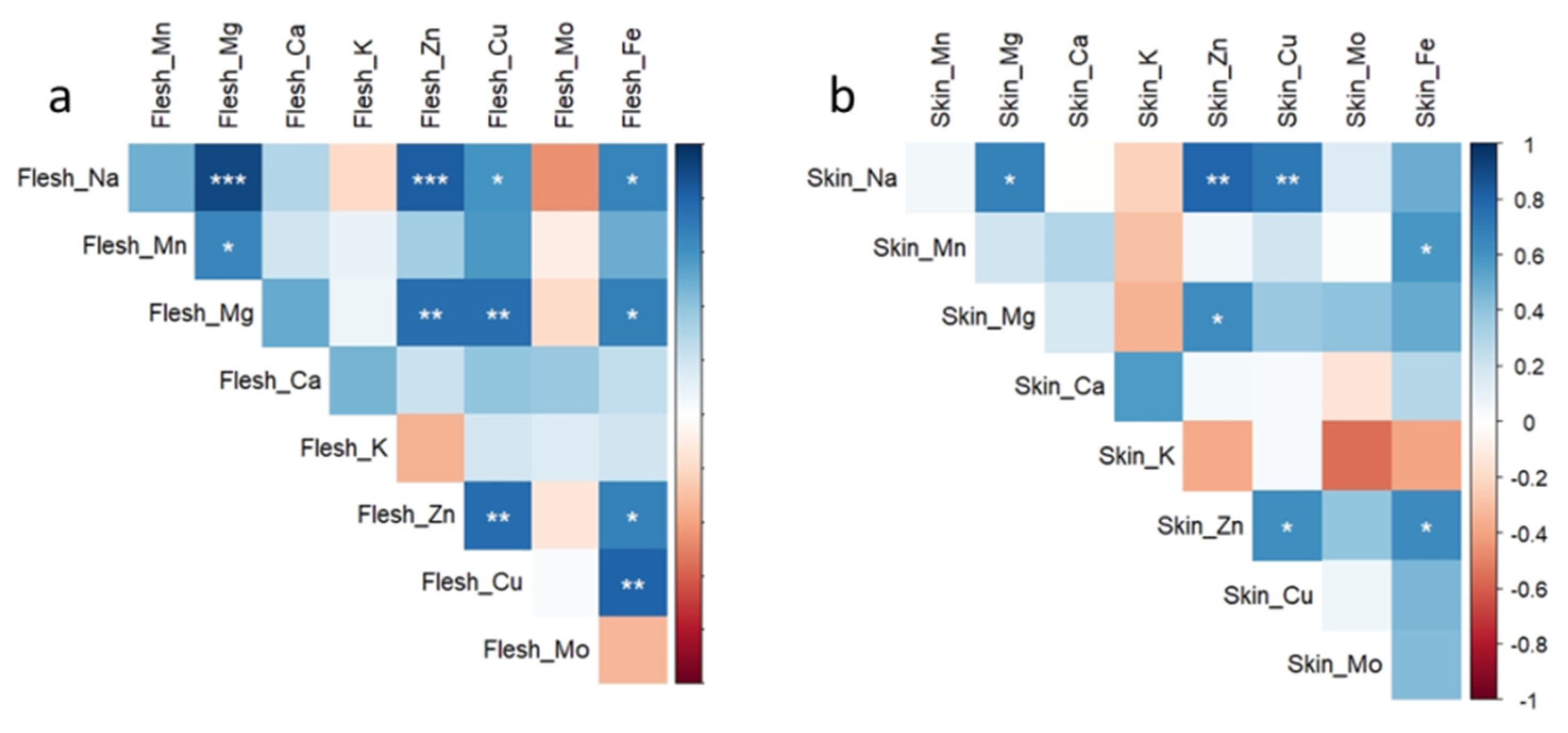
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mawa, S.; Husain, K.; Jantan, I. Ficus carica L. (Moraceae): Phytochemistry, traditional uses and biological activities. Evid.-Based Complement. Altern. Med. 2013, 2013, 974256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobaner, M.; Yurtal, R.; Dogan, A.; Motz, L.H. Three dimensional simulation of seawater intrusion in coastal aquifers: A case study in the Goksu Deltaic Plain. J. Hydrol. 2012, 464, 262–280. [Google Scholar] [CrossRef]
- Mollema, P.N.; Antonellini, M.; Dinelli, E.; Gabbianelli, G.; Greggio, N.; Stuyfzand, P.J. Hydrochemical and physical processes influencing salinization and freshening in Mediterranean low-lying coastal environments. Appl. Geochem. 2013, 34, 207–221. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS Homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 1750. [Google Scholar] [CrossRef]
- Irato, P.; Santovito, G. Enzymatic and non-enzymatic molecules with antioxidant function. Antioxidants 2021, 10, 579. [Google Scholar] [CrossRef] [PubMed]
- Petridis, A.; Therios, I.; Samouris, G.; Tananaki, C. Salinity-induced changes in phenolic compounds in leaves and roots of four olive cultivars (Olea europaea L.) and their relationship to antioxidant activity. Environ. Exp. Bot. 2012, 79, 37–43. [Google Scholar] [CrossRef]
- Caruso, G.; Palai, G.; Macheda, D.; Tozzini, L.; Gucci, R. Growth, gas exchange, water relations fresh and dry matter partitioning in young fig (Ficus carica L.) plants irrigated with saline water. Eur. J. Hort. Sci. 2022, 87. in press. [Google Scholar]
- Mascellani, A.; Natali, L.; Cavallini, A.; Mascagni, F.; Caruso, G.; Gucci, R.; Havlik, J.; Bernardi, R. Moderate salinity stress affects expression of main sugar metabolism and transport genes and soluble carbohydrate content in ripe fig fruits (Ficus carica L. cv. Dottato). Plants 2021, 10, 1861. [Google Scholar] [CrossRef]
- Vangelisti, A.; Zambrano, L.S.; Caruso, G.; Macheda, D.; Bernardi, R.; Usai, G.; Mascagni, F.; Giordani, T.; Gucci, R.; Cavallini, A.; et al. How an ancient, salt-tolerant fruit crop, Ficus carica L., copes with salinity: A transcriptome analysis. Sci. Rep. 2019, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Khayyat, M.; Tehranifar, A.; Davarynejad, G.H.; Sayyari-Zahan, M.H. Vegetative growth, compatible solute accumulation, ion partitioning and chlorophyll fluorescence of “Malas-e-Saveh” and “Shishe-Kab” pomegranates in response to salinity stress. Photosynthetica 2014, 52, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Tattini, M.; Gucci, R.; Coradeschi, M.A.; Ponzio, C.; Everard, J.D. Growth, gas exchange and ion content in Olea europaea plants during salinity stress and subsequent relief. Physiol. Plant. 1995, 95, 203–210. [Google Scholar] [CrossRef]
- Zarei, M.; Azizi, M.; Rahemi, M.; Tehranifar, A. Evaluation of NaCl salinity tolerance of four fig genotypes based on vegetative growth and ion content in leaves, shoots, and roots. HortScience 2016, 51, 1427–1434. [Google Scholar] [CrossRef] [Green Version]
- Metwali, E.M.R.; Soliman, H.I.A.; Fuller, M.P.; Al-Zahrani, H.S.; Howladar, S.M. Molecular cloning and expression of a vacuolar Na+/H+ antiporter gene (AgNHX1) in fig (Ficus carica L.) under salt stress. Plant Cell Tiss. Organ. Cult. 2015, 123, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Nejad, R.A.; Shekafandeh, A. Salt stress-induced change in leaf antioxidant activity, proline and protein content in ‘Shah Anjir’ and ‘Anjir Sabz’ fig seedlings. Int. J. Hort. Sci. Technol. 2014, 1, 121–129. [Google Scholar]
- Barolo, M.I.; Mostacero, N.R.; López, S.N. Ficus carica L. (Moraceae): An ancient source of food and health. Food Chem. 2014, 164, 119–127. [Google Scholar] [CrossRef]
- Solomon, A.; Golubowicz, S.; Yablowicz, Z.; Grossman, S.; Bergman, M.; Gottlieb, H.E.; Altman, A.; Kerem, Z.; Flaishman, M.A. Antioxidant activities and anthocyanin content of fresh fruits of common fig (Ficus carica L.). J. Agric. Food Chem. 2006, 54, 7717–7723. [Google Scholar] [CrossRef]
- Vallejo, F.; Marín, J.G.; Tomás-Barberán, F.A. Phenolic compound content of fresh and dried figs (Ficus carica L.). Food Chem. 2012, 130, 485–492. [Google Scholar] [CrossRef]
- Doaa, A.D.; El-Berry, I.M.; Mustafa, N.S.; Moursy, F.S.; Hagagg, L.F. Detecting drought tolerance of fig (Ficus carica, L.) cultivars depending on vegetative growth and peroxidase activity. Int. J. ChemTech Res. 2015, 8, 1520–1532. [Google Scholar]
- Oukabli, A.; Mekaoui, A.; Ibnouali-El-Aloui, M.; Bari, A. Contribution to identification of fig (Ficus carica) genotypes tolerant to drought. Acta Hortic. 2008, 798, 87–93. [Google Scholar] [CrossRef]
- Gholami, M.; Rahemi, M.; Kholdebarin, B.; Rastegar, S. Biochemical responses in leaves of four fig cultivars subjected to water stress and recovery. Sci. Hortic. 2012, 148, 109–117. [Google Scholar] [CrossRef]
- Fattorini, C.; Bernardi, R.; Quartacci, M.F.; Mascagni, F.; Caruso, G.; Cavallini, A.; Gucci, R.; Natali, L. Expression of genes involved in metabolism and transport of soluble carbohydrates during fruit ripening in two cultivars of Ficus carica L. Agrochimica 2021, 65, 117–136. [Google Scholar]
- Norfarah, I.R.; Adlina, M.; Nadia, M.E. A systematic review on the anti-oxidative and anti-inflammatory properties of Ficus carica. ASM Sci. J. 2018, 11, 124–145. [Google Scholar]
- Wojdyło, A.; Nowicka, P.; Carbonell-Barrachina, Á.A.; Hernández, F. Phenolic compounds, antioxidant and antidiabetic activity of different cultivars of Ficus carica L. fruits. J. Funct. Foods 2016, 25, 421–432. [Google Scholar] [CrossRef]
- Palmeira, L.; Pereira, C.; Dias, M.I.; Abreu, R.M.V.; Corrêa, R.C.G.; Pires, T.C.S.P.; Alves, M.J.; Barros, L.; Ferreira, I.C.F.R. Nutritional, chemical and bioactive profiles of different parts of a Portuguese common fig (Ficus carica L.) variety. Food Res. Int. 2019, 126, 108572. [Google Scholar] [CrossRef] [Green Version]
- Aljane, F.; Toumi, I.; Ferchichi, A. HPLC determination of sugars and atomic absorption analysis of mineral salts in fresh figs of Tunisian cultivars. Afr. J. Biotechnol. 2007, 6, 599–602. [Google Scholar] [CrossRef]
- Arvaniti, O.S.; Samaras, Y.; Gatidou, G.; Thomaidis, N.S.; Stasinakis, A.S. Review on fresh and dried figs: Chemical analysis and occurrence of phytochemical compounds, antioxidant capacity and health effects. Food Res. Int. 2019, 19, 244–267. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Khali, M.; Benkhaled, A.; Boucetta, I.; Dahmani, Y.; Attallah, Z.; Belbraouet, S. Fresh figs (Ficus carica L.): Pomological characteristics, nutritional value, and phytochemical properties. Eur. J. Hortic. Sci. 2018, 83, 104–113. [Google Scholar] [CrossRef]
- Bey, M.B.; Louaileche, H.; Zemouri, S. Optimization of phenolic compound recovery and antioxidant activity of light and dark dried fig (Ficus carica L.) varieties. Food Sci. Biotechnol. 2013, 22, 1613–1619. [Google Scholar] [CrossRef]
- Pereira, C.; López-Corrales, M.; Serradilla, M.J.; del Carmen Villalobos, M.; Ruiz-Moyano, S.; Martín, A. Influence of ripening stage on bioactive compounds and antioxidant activity in nine fig (Ficus carica L.) varieties grown in Extremadura, Spain. J. Food Comp. Anal. 2017, 64, 203–212. [Google Scholar] [CrossRef]
- Šamec, D.; Liníc, I.; Salopek-Sondi, B. Salinity stress as an elicitor for phytochemicals and minerals accumulation in selected leafy vegetables of Brassicaceae. Agronomy 2021, 11, 361. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E. Effect of salinity on yield, fruit quality, leaf gas exchange, and mineral composition of grafted watermelon plants. HortScience 2006, 41, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Saied, A.S.; Keutgen, A.J.; Noga, G. The influence of NaCl salinity on growth, yield and fruit quality of strawberry cvs. “Elsanta” and “Korona”. Sci. Hortic. 2005, 103, 289–303. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tukey, J.W. Exploratory Data Analysis; Addison-Wesley Publishing, Co.: Reading, MA, USA, 1977; p. 688. ISBN 0-201-07616-0. [Google Scholar]
- Ficsor, E.; Szentmihályi, K.; Lemberkovics, É.; Blázovics, A.; Balázs, A. Analysis of Ficus carica L.–volatile components and mineral content. Eur. Chem. Bull. 2013, 2, 126–129. [Google Scholar]
- Garza-Alonso, C.A.; Olivares-Sáenz, E.; Gutiérrez-Díez, A.; Vázquez-Alvarado, R.E.; López-Jiménez, A. Visual Symptoms, Vegetative Growth, and Mineral Concentration in Fig Tree (Ficus carica L.) Under Macronutrient Deficiencies. Agronomy 2019, 9, 787. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Li, Y.; Han, G.; Song, J.; Wang, B. NaCl markedly improved the reproductive capacity of the euhalophyte Suaeda salsa. Funct. Plant Biol. 2018, 45, 350–361. [Google Scholar] [CrossRef]
- Khan, M.A.; Ungar, I.A.; Showalter, A.M. Effects of sodium chloride treatments on growth and ion accumulation of the halophyte Haloxylon recurvum. Commun. Soil Sci. Plant Anal. 2000, 31, 2763–2774. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Aloisi, I.; Parrotta, L.; Ruiz, K.B.; Landi, C.; Bini, L.; Cai, G.; Biondi, S.; Del Duca, S. New insight into quinoa seed quality under salinity: Changes in proteomic and amino acid profiles, phenolic content, and antioxidant activity of protein extracts. Front. Plant Sci. 2016, 7, 656. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Perin, E.C.; da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-dependent salt and drought stress improve strawberry fruit quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef]
- Cuong, D.M.; Kwon, S.J.; Van Nguyen, B.; Chun, S.W.; Kim, J.K.; Park, S.U. Effect of salinity stress on phenylpropanoid genes expression and related gene expression in wheat sprout. Agronomy 2020, 10, 390. [Google Scholar] [CrossRef] [Green Version]
- Yiu, J.C.; Tseng, M.J.; Liu, C.W.; Kuo, C.T. Modulation of NaCl stress in Capsicum annuum L. seedlings by catechin. Sci. Hortic. 2012, 134, 200–209. [Google Scholar] [CrossRef]
- Scagel, C.F.; Lee, J.; Mitchell, J.N. Salinity from NaCl changes the nutrient and polyphenolic composition of basil leaves. Ind. Crop. Prod. 2019, 127, 119–128. [Google Scholar] [CrossRef]
- Ghafari, H.; Hassanpour, H.; Jafari, M.; Besharat, S. Physiological, biochemical and gene-expressional responses to water deficit in apple subjected to partial root-zone drying (PRD). Plant Physiol. Biochem. 2020, 148, 333–346. [Google Scholar] [CrossRef]
- Li, X.; Ahammed, G.J.; Zhang, X.N.; Zhang, L.; Yan, P.; Zhang, L.P.; Fu, J.Y.; & Han, W.Y. Melatonin-mediated regulation of anthocyanin biosynthesis and antioxidant defense confer tolerance to arsenic stress in Camellia sinensis L. J. Hazard. Mater. 2021, 403, 123922. [Google Scholar] [CrossRef] [PubMed]
- Torabian, S.; Zahedi, M.; Khoshgoftar, A.H. Effects of foliar spray of two kinds of zinc oxide on the growth and ion concentration of sunflower cultivars under salt stress. J. Plant Nutr. 2016, 39, 172–180. [Google Scholar] [CrossRef]
- Shanmugam, V.; Lo, J.C.; Yeh, K.C. Control of Zn uptake in Arabidopsis halleri: A balance between Zn and Fe. Front. Plant Sci. 2013, 4, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, A.U.; Hadi, F.; Nawaz, M.A.; Rahman, K. Potassium and zinc increase tolerance to salt stress in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2017, 116, 139–149. [Google Scholar] [CrossRef]
- Akanbi-Gada, M.A.; Ogunkunle, C.O.; Vishwakarma, V.; Viswanathan, K.; Fatoba, P.O. Phytotoxicity of nano-zinc oxide to tomato plant (Solanum lycopersicum L.): Zn uptake, stress enzymes response and influence on non-enzymatic antioxidants in fruits. Environ. Technol. Innov. 2019, 14, 100325. [Google Scholar] [CrossRef]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt stress induces physiochemical alterations in rice grain composition and quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Printz, B.; Lutts, S.; Hausman, J.F.; Sergeant, K. Copper trafficking in plants and its implication on cell wall dynamics. Front. Plant Sci. 2016, 7, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Koch, W. Dietary Polyphenols—Important non-nutrients in the prevention of chronic noncommunicable diseases. Nutrients 2019, 11, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- De Pascale, S.; Maggio, A.; Fogliano, V.; Ambrosino, P.; Ritieni, A. Irrigation with saline water improves carotenoids content and antioxidant activity of tomato. J. Hortic. Sci. Biotechnol. 2001, 76, 447–453. [Google Scholar] [CrossRef]
- Botía, P.; Navarro, J.M.; Cerdá, A.; Martínez, V. Yield and fruit quality of two melon cultivars irrigated with saline water at different stages of development. Eur. J. Agron. 2005, 23, 243–253. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Acronym | RT (min) | Q1 | Q3 | DP (V) | CE (eV) | CXP (V) |
|---|---|---|---|---|---|---|---|
| Catechin | CTC | 2.33 | 289.0 | 244.9 | −108 | −22 | −11 |
| Chlorogenic Acid | CGA | 2.44 | 353.0 | 191.0 | −61 | −24 | −9 |
| Epicatechin | ECTC | 2.63 | 289.0 | 244.9 | −108 | −22 | −11 |
| Quercetagetin 7-O-Glucoside | Q7G | 3.07 | 479.1 | 316.9 | −152 | −31 | −14 |
| p-Coumaric Acid | PCA | 3.09 | 163.0 | 119.0 | −65 | −18 | −11 |
| Trans-Ferulic Acid | TFRA | 3.28 | 193.0 | 134.0 | −62 | −20 | −8 |
| Rutin | RTN | 3.30 | 609.2 | 299.9 | −154 | −48 | −11 |
| Quercetin 3-O-Glucoside | QCT3G | 3.40 | 463.1 | 300.0 | −154 | −37 | −5 |
| Kaempferol 3-O-Rutinoside | KPF3R | 3.53 | 593.2 | 284.9 | −138 | −40 | −5 |
| Kaempferol 3-O-Glucoside | KPF3G | 3.62 | 447.1 | 284.1 | −202 | −39 | −11 |
| Kaempferol 7-O-Glucoside | KPF7G | 3.66 | 447.1 | 284.9 | −158 | −38 | −5 |
| Phloridzin | PDZ | 3.71 | 435.1 | 272.9 | −135 | −23 | −5 |
| Quercetin | QCT | 4.30 | 301.0 | 150.9 | −113 | −38 | −8 |
| Tiliroside | TSD | 4.41 | 593.2 | 284.9 | −138 | −40 | −5 |
| Phloretin | PHL | 4.64 | 273.0 | 167.0 | −103 | −38 | −11 |
| Catechin | CTC | 2.33 | 289.0 | 244.9 | −108 | −22 | −11 |
| Treatments (NaCl) | |||
|---|---|---|---|
| 0 mM | 100 mM | t-test | |
| Shoot Length Increment (mm) | 73 ± 15.4 | 56 ± 15.1 | ns |
| Fruit Weight (g) | 18.0 ± 5.88 | 16.1 ± 7.06 | ns |
| Leaf Proline (mg g−1 FW) | 0.28 ± 0.050 | 0.26 ± 0.030 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francini, A.; Sodini, M.; Vicario, G.; Raffaelli, A.; Gucci, R.; Caruso, G.; Sebastiani, L. Cations and Phenolic Compounds Concentrations in Fruits of Fig Plants Exposed to Moderate Levels of Salinity. Antioxidants 2021, 10, 1865. https://doi.org/10.3390/antiox10121865
Francini A, Sodini M, Vicario G, Raffaelli A, Gucci R, Caruso G, Sebastiani L. Cations and Phenolic Compounds Concentrations in Fruits of Fig Plants Exposed to Moderate Levels of Salinity. Antioxidants. 2021; 10(12):1865. https://doi.org/10.3390/antiox10121865
Chicago/Turabian StyleFrancini, Alessandra, Mirko Sodini, Giulia Vicario, Andrea Raffaelli, Riccardo Gucci, Giovanni Caruso, and Luca Sebastiani. 2021. "Cations and Phenolic Compounds Concentrations in Fruits of Fig Plants Exposed to Moderate Levels of Salinity" Antioxidants 10, no. 12: 1865. https://doi.org/10.3390/antiox10121865
APA StyleFrancini, A., Sodini, M., Vicario, G., Raffaelli, A., Gucci, R., Caruso, G., & Sebastiani, L. (2021). Cations and Phenolic Compounds Concentrations in Fruits of Fig Plants Exposed to Moderate Levels of Salinity. Antioxidants, 10(12), 1865. https://doi.org/10.3390/antiox10121865