
Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Groups and Setting
- Male control (MCo, n = 18)
- Male exercised (MEx, n = 18)
- Female control (FCo, n = 18)
- Female exercised (FEx, n = 18).
2.3. Exercise Training Protocol—Rat Model of Physiological Cardiac Hypertrophy
2.4. Echocardiography
2.5. In Vivo Right Atrial Electrophysiology
2.6. Quantitative Real-Time Polymerase Chain Reaction
2.7. Histology
2.8. Statistical Analysis
3. Results
3.1. Cardiac Morphological Alterations
3.2. In Vivo Electrophysiology
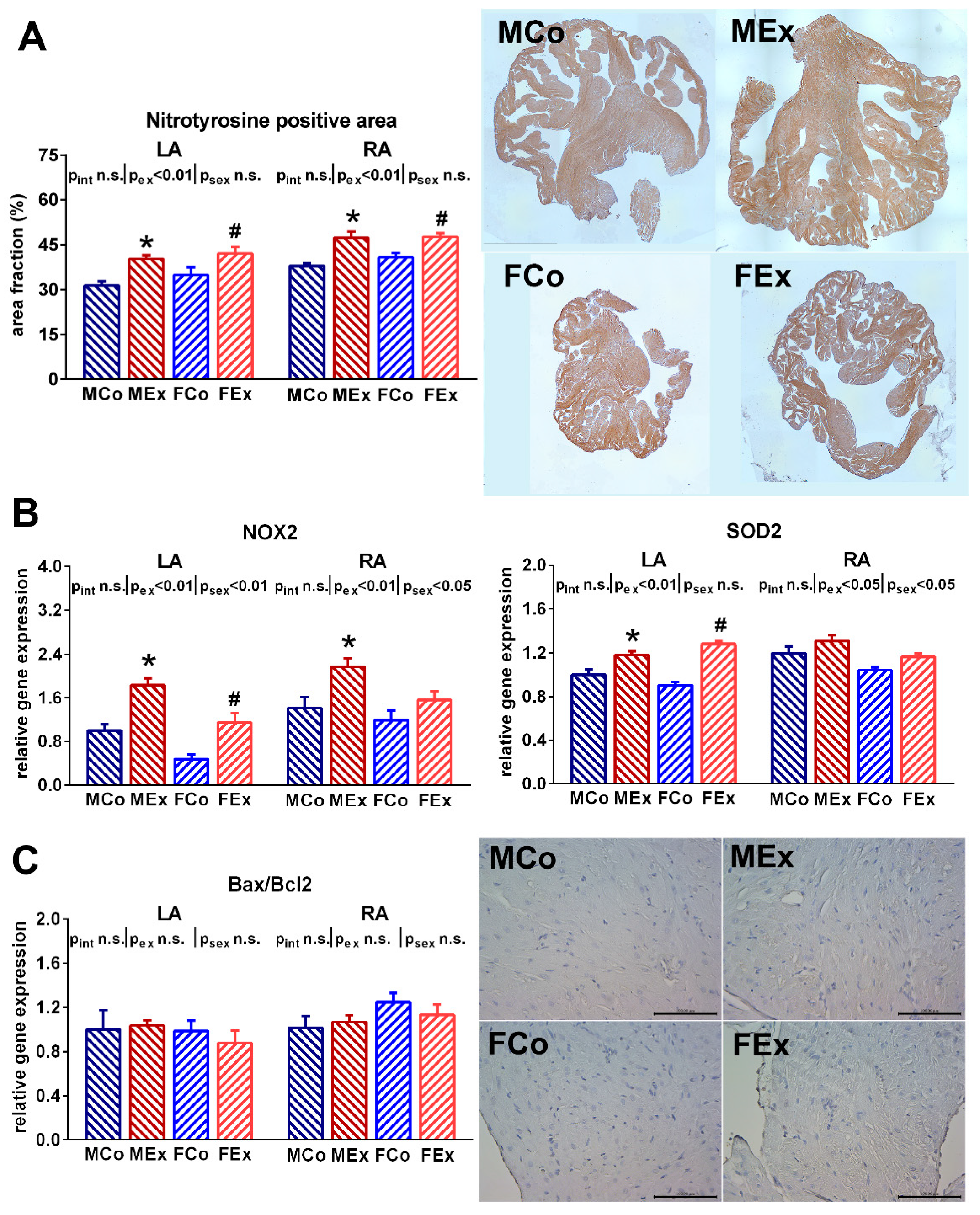
3.3. Atrial Oxidative Stress and Tissue Remodelling
3.4. Molecular Alterations Related to Electrical Remodeling
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- La Gerche, A.; Prior, D.L. Exercise--is it possible to have too much of a good thing? Heart Lung Circ. 2007, 16 (Suppl. 3), S102–S104. [Google Scholar] [CrossRef]
- Merghani, A.; Malhotra, A.; Sharma, S. The U-shaped relationship between exercise and cardiac morbidity. Trends Cardiovasc. Med. 2016, 26, 232–240. [Google Scholar] [CrossRef]
- Sharma, S.; Merghani, A.; Mont, L. Exercise and the heart: The good, the bad, and the ugly. Eur. Hear. J. 2015, 36, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-C.; Pate, R.R.; Lavie, C.J.; Sui, X.; Church, T.S.; Blair, S.N. Leisure-Time Running Reduces All-Cause and Cardiovascular Mortality Risk. J. Am. Coll. Cardiol. 2014, 64, 472–481. [Google Scholar] [CrossRef] [Green Version]
- Oláh, A.; Mátyás, C.; Kellermayer, D.; Ruppert, M.; Barta, B.A.; Sayour, A.A.; Török, M.; Koncsos, G.; Giricz, Z.; Ferdinandy, P.; et al. Sex Differences in Morphological and Functional Aspects of Exercise-Induced Cardiac Hypertrophy in a Rat Model. Front. Physiol. 2019, 10, 889. [Google Scholar] [CrossRef] [Green Version]
- D’Ascenzi, F.; Anselmi, F.; Focardi, M.; Mondillo, S. Atrial Enlargement in the Athlete’s Heart: Assessment of Atrial Function May Help Distinguish Adaptive from Pathologic Remodeling. J. Am. Soc. Echocardiogr. 2018, 31, 148–157. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.H.; Patil, H.R.; Lavie, C.J.; Magalski, A.; Vogel, R.A.; McCullough, P.A. Potential Adverse Cardiovascular Effects from Excessive Endurance Exercise. Mayo Clin. Proc. 2012, 87, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Oláh, A.; Németh, B.T.; Matyas, C.; Horvath, E.M.; Hidi, L.; Birtalan, E.; Kellermayer, D.; Ruppert, M.; Merkely, G.; Szabo, G.; et al. Cardiac effects of acute exhaustive exercise in a rat model. Int. J. Cardiol. 2015, 182, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Estes, N.A.M., 3rd; Madias, C. Atrial Fibrillation in Athletes: A Lesson in the Virtue of Moderation. JACC Clin. Electrophysiol. 2017, 3, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Gomar, F.; Perez-Quilis, C.; Lippi, G.; Cervellin, G.; Leischik, R.; Löllgen, H.; Serrano-Ostáriz, E.; Lucia, A. Atrial fibrillation in highly trained endurance athletes—Description of a syndrome. Int. J. Cardiol. 2017, 226, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Guasch, E.; Mont, L.; Sitges, M. Mechanisms of atrial fibrillation in athletes: What we know and what we do not know. Neth. Hear. J. 2018, 26, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finocchiaro, G.; Dhutia, H.; D’Silva, A.; Malhotra, A.; Steriotis, A.; Millar, L.; Prakash, K.; Narain, R.; Papadakis, M.; Sharma, R.; et al. Effect of Sex and Sporting Discipline on LV Adaptation to Exercise. JACC Cardiovasc. Imaging 2017, 10, 965–972. [Google Scholar] [CrossRef]
- Nattel, S. Physical activity and atrial fibrillation risk: it’s complicated; and sex is critical. Eur. Hear. J. 2020, 41, 1487–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovits, T.; Oláh, A.; Lux, Á.; Németh, B.T.; Hidi, L.; Birtalan, E.; Kellermayer, D.; Matyas, C.; Szabó, G.; Merkely, B. Rat model of exercise-induced cardiac hypertrophy: Hemodynamic characterization using left ventricular pressure-volume analysis. Am. J. Physiol. Circ. Physiol. 2013, 305, H124–H134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oláh, A.; Kellermayer, D.; Mátyás, C.; Németh, B.T.; Lux, Á.; Szabó, L.; Török, M.; Ruppert, M.; Meltzer, A.; Sayour, A.A.; et al. Complete Reversion of Cardiac Functional Adaptation Induced by Exercise Training. Med. Sci. Sports Exerc. 2017, 49, 420–429. [Google Scholar] [CrossRef]
- Oláh, A.; Kovács, A.; Lux, Á.; Tokodi, M.; Braun, S.; Lakatos, B.K.; Mátyás, C.; Kellermayer, D.; Ruppert, M.; Sayour, A.A.; et al. Characterization of the dynamic changes in left ventricular morphology and function induced by exercise training and detraining. Int. J. Cardiol. 2019, 277, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foryst-Ludwig, A.; Kintscher, U. Sex differences in exercise-induced cardiac hypertrophy. Pflügers Archiv Eur. J. Physiol. 2013, 465, 731–737. [Google Scholar] [CrossRef]
- Guasch, E.; Benito, B.; Qi, X.; Cifelli, C.; Naud, P.; Shi, Y.; Mighiu, A.; Tardif, J.C.; Tadevosyan, A.; Chen, Y.; et al. Atrial fibrillation promotion by endurance exercise: Demonstration and mechanistic exploration in an animal model. J. Am. Coll. Cardiol. 2013, 62, 68–77. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, A.; Riegler, L.; Cocchia, R.; Scarafile, R.; Salerno, G.; Gravino, R.; Golia, E.; Vriz, O.; Citro, R.; Limongelli, G.; et al. Left atrial volume index in highly trained athletes. Am. Heart J. 2010, 159, 1155–1161. [Google Scholar] [CrossRef]
- Pelliccia, A.; Maron, B.J.; Di Paolo, F.M.; Biffi, A.; Quattrini, F.M.; Pisicchio, C.; Roselli, A.; Caselli, S.; Culasso, F. Prevalence and Clinical Significance of Left Atrial Remodeling in Competitive Athletes. J. Am. Coll. Cardiol. 2005, 46, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Mosén, H.; Steding-Ehrenborg, K. Atrial remodelling is less pronounced in female endurance-trained athletes compared with that in male athletes. Scand. Cardiovasc. J. 2013, 48, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, V.R.; Kannan, S.; Sadhaasivam, K.; Gounder, S.S.; Davidson, C.J.; Boeheme, C.; Hoidal, J.R.; Wang, L.; Rajasekaran, N.S. Acute exercise stress activates Nrf2/ARE signaling and promotes antioxidant mechanisms in the myocardium. Free. Radic. Biol. Med. 2012, 52, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Korantzopoulos, P.; Letsas, K.P.; Tse, G.; Fragakis, N.; Goudis, C.A.; Liu, T. Inflammation and atrial fibrillation: A comprehensive review. J. Arrhythmia 2018, 34, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Lou, H.; Singal, P.K. Oxidative Stress and Apoptosis in Heart Dysfunction. Herz 2002, 27, 662–668. [Google Scholar] [CrossRef]
- de Bold, A.J.; Bruneau, B.G.; de Kuroski Bold, M.L. Mechanical and neuroendocrine regulation of the endocrine heart. Cardiovasc. Res. 1996, 31, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Aschar-Sobbi, R.; Izaddoustdar, F.; Korogyi, A.S.; Wang, Q.; Farman, G.P.; Yang, F.; Yang, W.; Dorian, D.; Simpson, J.A.; Tuomi, J.M.; et al. Increased atrial arrhythmia susceptibility induced by intense endurance exercise in mice requires TNFα. Nat. Commun. 2015, 6, 6018. [Google Scholar] [CrossRef]
- Bernecker, C.; Scherr, J.; Schinner, S.; Braun, S.; Scherbaum, W.A.; Halle, M. Evidence for an exercise induced increase of TNF-α and IL-6 in marathon runners. Scand. J. Med. Sci. Sports 2011, 23, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Gal, P.; Marrouche, N.F. Magnetic resonance imaging of atrial fibrosis: Redefining atrial fibrillation to a syndrome. Eur. Hear. J. 2017, 38, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Benito, B.; Gay-Jordi, G.; Serrano-Mollar, A.; Guasch, E.; Shi, Y.; Tardif, J.-C.; Brugada, J.; Nattel, S.; Mont, L. Cardiac Arrhythmogenic Remodeling in a Rat Model of Long-Term Intensive Exercise Training. Circulation 2011, 123, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Stein, R.; Medeiros, C.M.; A Rosito, G.; I Zimerman, L.; Ribeiro, J.P. Intrinsic sinus and atrioventricular node electrophysiologic adaptations in endurance athletes. J. Am. Coll. Cardiol. 2002, 39, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Such, L.; Rodriguez, A.; Alberola, A.; Lopez, L.; Ruiz, R.; Artal, L.; Pons, I.; Pons, M.L.; García, C.; Chorro, F.J. Intrinsic changes on automatism, conduction, and refractoriness by exercise in isolated rabbit heart. J. Appl. Physiol. 2002, 92, 225–229. [Google Scholar] [CrossRef]
- Besnier, F.; Labrunée, M.; Pathak, A.; Traon, A.P.-L.; Galès, C.; Sénard, J.-M.; Guiraud, T. Exercise training-induced modification in autonomic nervous system: An update for cardiac patients. Ann. Phys. Rehabil. Med. 2017, 60, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Wehrens, X.H.; Chiang, D.Y.; Li, N. Chronic exercise: A contributing factor to atrial fibrillation? J. Am. Coll. Cardiol. 2013, 62, 78–80. [Google Scholar] [CrossRef] [Green Version]
- Kamalvand, K.; Tan, K.; Lloyd, G.; Gill, J.; Bucknall, C.; Sulke, N. Alterations in atrial electrophysiology associated with chronic atrial fibrillation in man. Eur. Hear. J. 1999, 20, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.; Caballero, R.; Gómez, R.; Delpón, E. IKur/Kv1.5 channel blockers for the treatment of atrial fibrillation. Expert Opin. Investig. Drugs 2009, 18, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Goudis, C.A.; Korantzopoulos, P.; Ntalas, I.V.; Kallergis, E.M.; Liu, T.; Ketikoglou, D.G. Diabetes mellitus and atrial fibrillation: Pathophysiological mechanisms and potential upstream therapies. Int. J. Cardiol. 2015, 184, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yu, T.; Yang, H.; Peng, Z. Exhaustive Exercise-Induced Cardiac Conduction System Injury and Changes of cTnT and Cx43. Int. J. Sports Med. 2014, 36, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.R.; Wachtell, K.; Abdulla, J. The Relationship Between Physical Activity and Risk of Atrial Fibrillation-A Systematic Review and Meta-Analysis. J. Atr. Fibrillation 2013, 5, 789. [Google Scholar] [PubMed]
- Colombo, C.S.S.S.; Finocchiaro, G. The Female Athlete’s Heart: Facts and Fallacies. Curr. Treat. Options Cardiovasc. Med. 2018, 20, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dworatzek, E.; Mahmoodzadeh, S.; Schriever, C.; Kusumoto, K.; Kramer, L.; Santos, G.; Fliegner, D.; Leung, Y.-K.; Ho, S.-M.; Zimmermann, W.-H.; et al. Sex-specific regulation of collagen I and III expression by 17β-Estradiol in cardiac fibroblasts: Role of estrogen receptors. Cardiovasc. Res. 2019, 115, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.D.; Lehmann, M.H.; Ritchie, R.H.; Gwathmey, J.K.; Green, G.E.; Schiebinger, R.J. Androgen Receptors Mediate Hypertrophy in Cardiac Myocytes. Circulation 1998, 98, 256–261. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MCo (n = 12) | MEx (n = 12) | FCo (n = 12) | FEx (n = 12) | pint | pex | psex | |
|---|---|---|---|---|---|---|---|
| Heart rate (beat/min) | 375 ± 7 | 355 ± 9 | 367 ± 5 | 353 ± 7 | n.s. | <0.05 | n.s. |
| P length (ms) | 16.3 ± 0.3 | 17.1 ± 0.3 | 15.9 ± 0.5 | 18.4 ± 0.5 # | n.s. | <0.01 | n.s. |
| P amplitude (mV) | 0.12 ± 0.01 | 0.16 ± 0.01 * | 0.14 ± 0.01 | 0.19 ± 0.01 # | n.s. | <0.01 | <0.01 |
| PR length (ms) | 49.4 ± 0.9 | 51.9 ± 1.7 | 49.4 ± 1.1 | 53.7 ± 1.1 | n.s. | <0.05 | n.s. |
| QRS length (ms) | 18.2 ± 0.2 | 18.7 ± 0.3 | 18.7 ± 0.5 | 19.0 ± 0.6 | n.s. | n.s. | n.s. |
| QRS amplitude (mV) | 0.85 ± 0.05 | 1.05 ± 0.05 * | 1.02 ± 0.05 | 1.33 ± 0.06 # | n.s. | <0.01 | <0.01 |
| QT length (ms) | 65.4 ± 3.5 | 76.7 ± 1.9 * | 74.4 ± 2.5 | 80.9 ± 3.6 # | n.s. | <0.01 | <0.05 |
| T amplitude (mV) | 0.09 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.16 ± 0.02 | n.s. | <0.05 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oláh, A.; Barta, B.A.; Sayour, A.A.; Ruppert, M.; Virág-Tulassay, E.; Novák, J.; Varga, Z.V.; Ferdinandy, P.; Merkely, B.; Radovits, T. Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity. Antioxidants 2021, 10, 452. https://doi.org/10.3390/antiox10030452
Oláh A, Barta BA, Sayour AA, Ruppert M, Virág-Tulassay E, Novák J, Varga ZV, Ferdinandy P, Merkely B, Radovits T. Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity. Antioxidants. 2021; 10(3):452. https://doi.org/10.3390/antiox10030452
Chicago/Turabian StyleOláh, Attila, Bálint András Barta, Alex Ali Sayour, Mihály Ruppert, Eszter Virág-Tulassay, Julianna Novák, Zoltán V. Varga, Péter Ferdinandy, Béla Merkely, and Tamás Radovits. 2021. "Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity" Antioxidants 10, no. 3: 452. https://doi.org/10.3390/antiox10030452
APA StyleOláh, A., Barta, B. A., Sayour, A. A., Ruppert, M., Virág-Tulassay, E., Novák, J., Varga, Z. V., Ferdinandy, P., Merkely, B., & Radovits, T. (2021). Balanced Intense Exercise Training Induces Atrial Oxidative Stress Counterbalanced by the Antioxidant System and Atrial Hypertrophy That Is Not Associated with Pathological Remodeling or Arrhythmogenicity. Antioxidants, 10(3), 452. https://doi.org/10.3390/antiox10030452