
Nutraceutical Supplementation Ameliorates Visual Function, Retinal Degeneration, and Redox Status in rd10 Mice

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Methods
2.1. Animals and Treatment
2.2. ERG Recordings
2.3. Retinal Histology
2.4. Microscopy and Quantification
2.5. Determination of Antioxidant and Oxidant Markers
2.6. Gene Expression Analysis
2.7. Statistical Analysis
3. Results
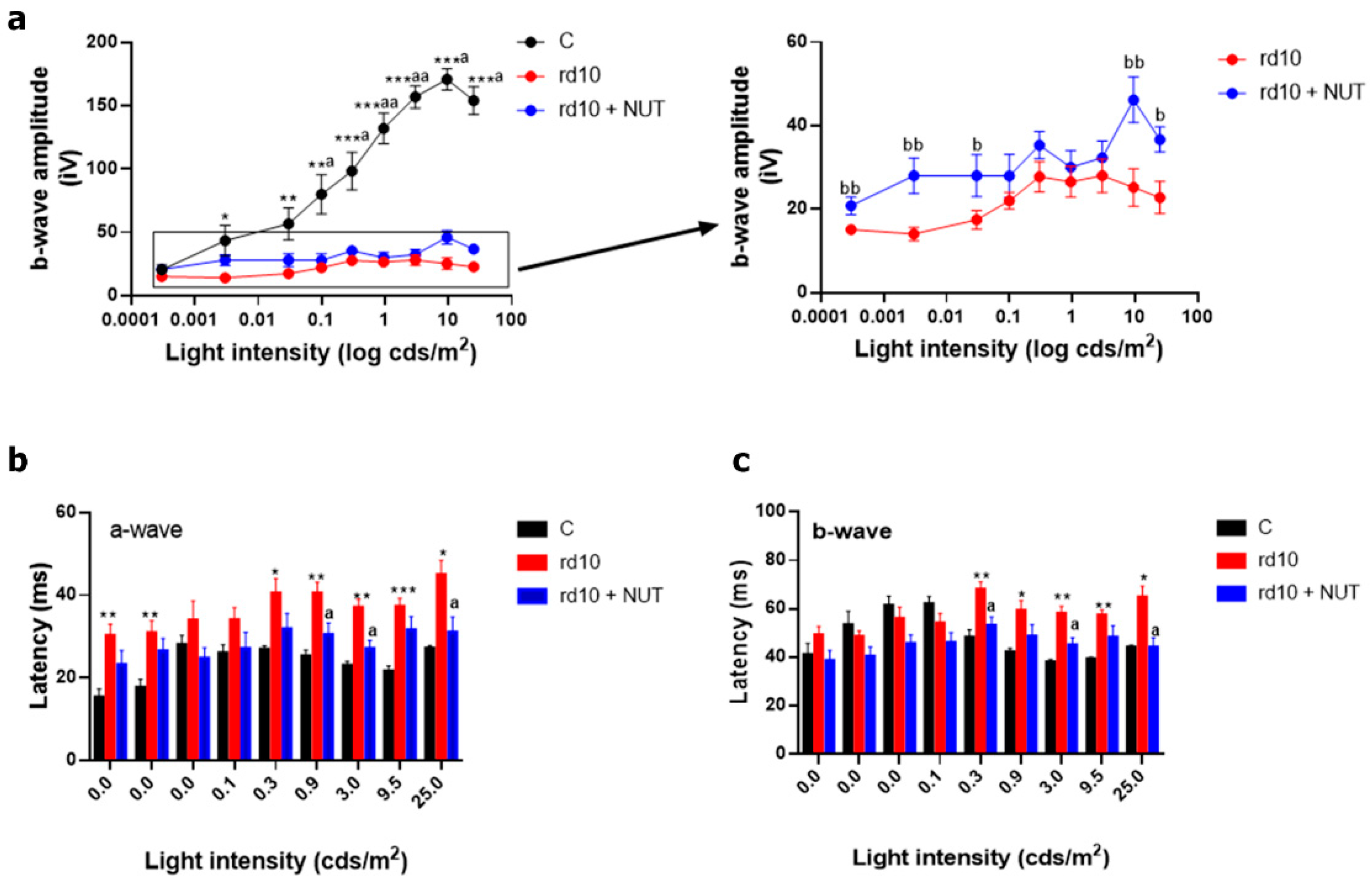
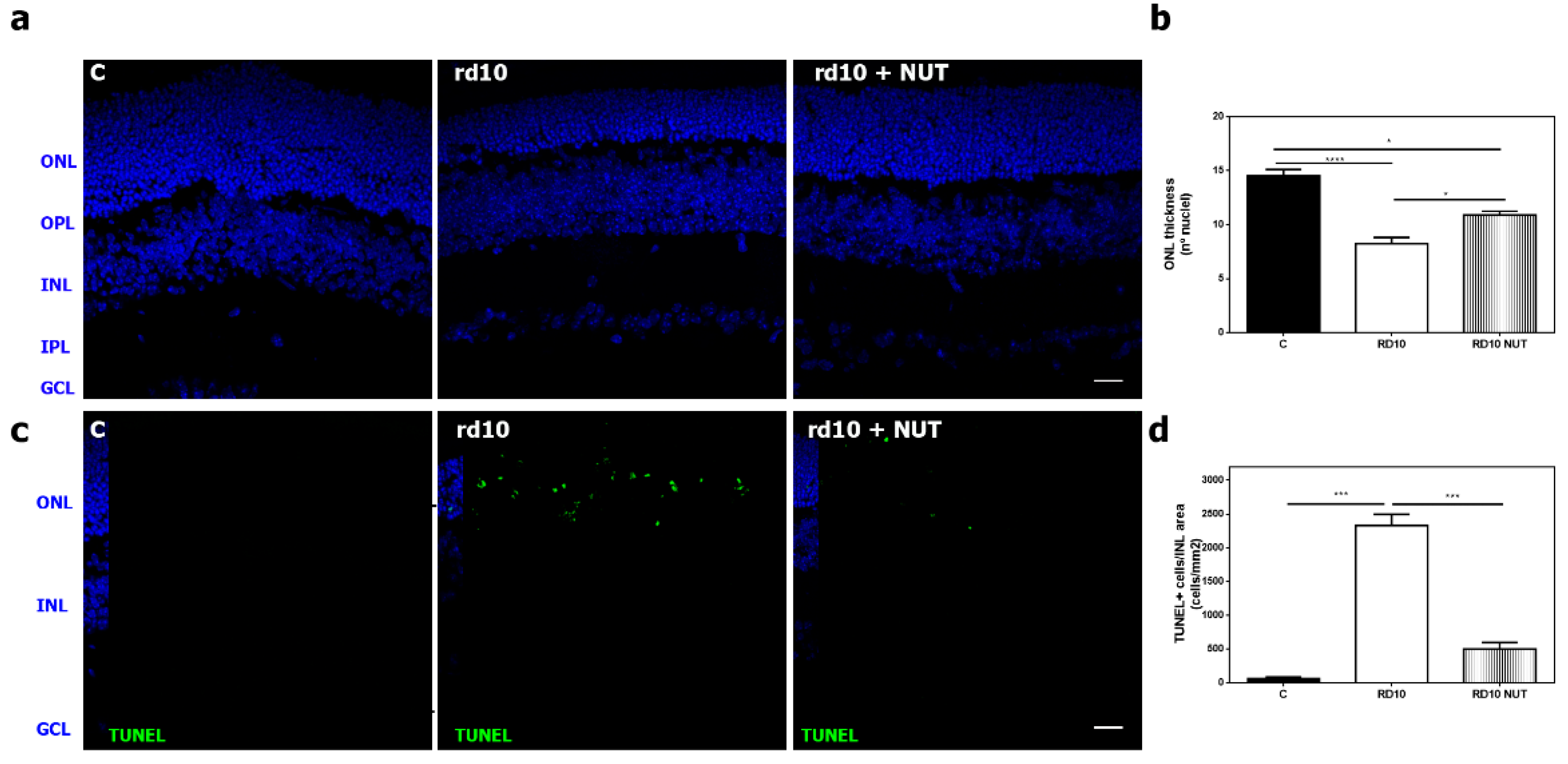
3.1. Oral Administration of Antioxidant Nutraceuticals Ameliorated Retinal Degeneration in rd10 Mice at PD18
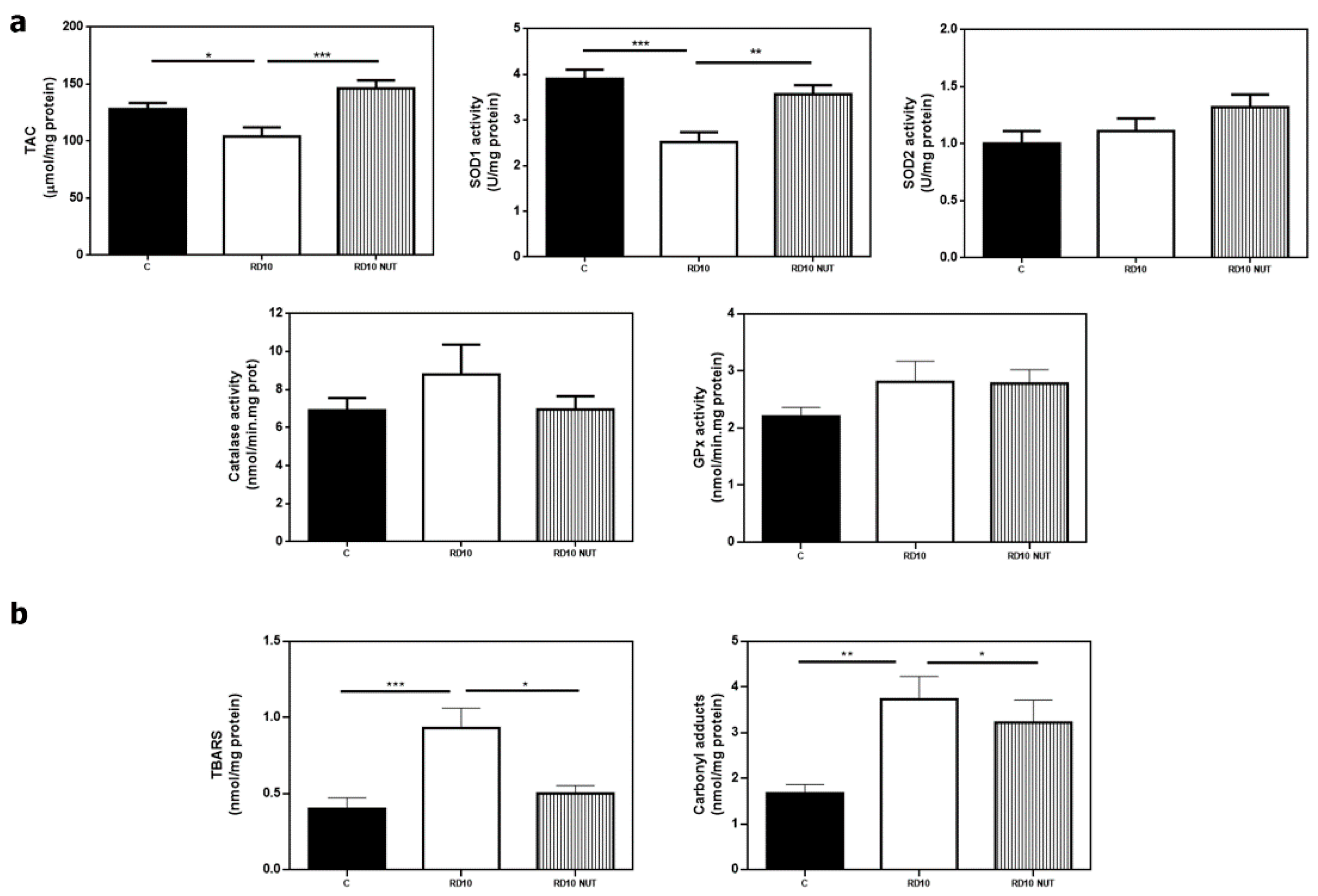
3.2. Oral Administration of Antioxidant Nutraceuticals Partially Restored Retinal Redox Status in rd10 Mice at P18
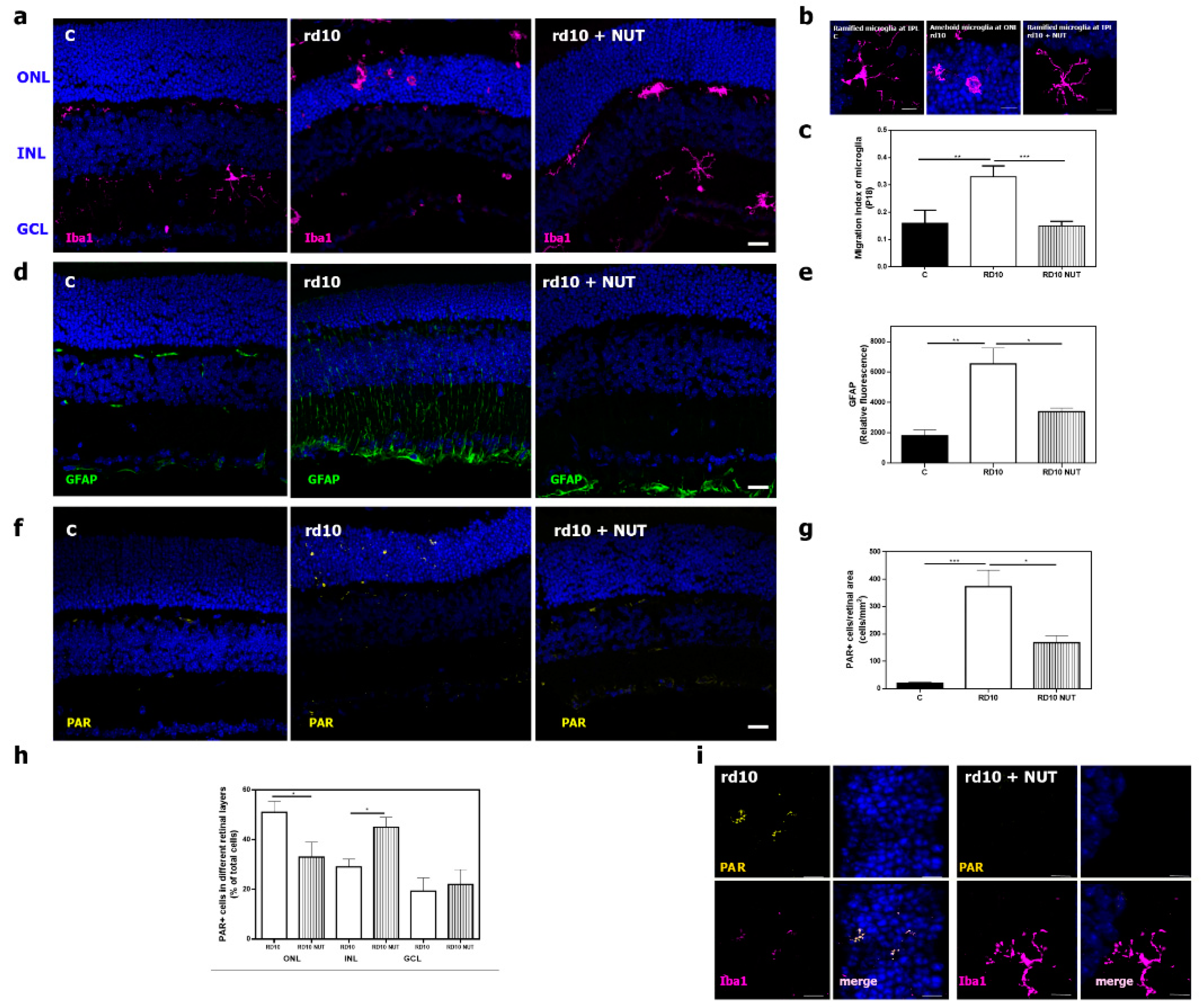
3.3. Oral Administration of Antioxidant Nutraceuticals Reduced Retinal Inflammation in rd10 Mice at PD18
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rivas, M.A.; Vecino, E. Animal models and different therapies for treatment of retinitis pigmentosa. Histol. Histopathol. 2009, 24, 1295–1322. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Betteridge, D.J. What is oxidative stress? Metab. Clin. Exp. 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Sickel, W. Electrical and metabolic manifestations of receptor and higher-order neuron activity in vertebrate retina. In Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1972; Volume 24, pp. 101–118. [Google Scholar] [CrossRef]
- Usui, S.; Oveson, B.C.; Lee, S.Y.; Jo, Y.J.; Yoshida, T.; Miki, A.; Miki, K.; Iwase, T.; Lu, L.; Campochiaro, P.A. NADPH oxidase plays a central role in cone cell death in retinitis pigmentosa. J. Neurochem. 2009, 110, 1028–1037. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, K.M.; Carvalho, L.S.; Rizzi, M.; Powell, K.; Holthaus, S.M.; Azam, S.A.; Duran, Y.; Ribeiro, J.; Luhmann, U.F.; Bainbridge, J.W.; et al. Gene therapy restores vision in rd1 mice after removal of a confounding mutation in Gpr179. Nat. Commun. 2015, 6, 6006. [Google Scholar] [CrossRef]
- Shen, J.; Yang, X.; Dong, A.; Petters, R.M.; Peng, Y.W.; Wong, F.; Campochiaro, P.A. Oxidative damage is a potential cause of cone cell death in retinitis pigmentosa. J. Cell. Physiol. 2005, 203, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Ikeda, Y.; Yoshida, N.; Notomi, S.; Hisatomi, T.; Oka, S.; De Luca, G.; Yonemitsu, Y.; Bignami, M.; Nakabeppu, Y.; et al. MutT homolog-1 attenuates oxidative DNA damage and delays photoreceptor cell death in inherited retinal degeneration. Am. J. Pathol. 2012, 181, 1378–1386. [Google Scholar] [CrossRef]
- Martínez-Fernández de la Cámara, C.; Hernández-Pinto, A.M.; Olivares-González, L.; Cuevas-Martín, C.; Sánchez-Aragó, M.; Hervás, D.; Salom, D.; Cuezva, J.M.; de la Rosa, E.J.; Millán, J.M.; et al. Adalimumab reduces photoreceptor cell death in a mouse model of retinal degeneration. Sci. Rep. 2015, 5, 11764. [Google Scholar] [CrossRef]
- Olivares-González, L.; Martínez-Fernández de la Cámara, C.; Hervás, D.; Millán, J.M.; Rodrigo, R. HIF-1α stabilization reduces retinal degeneration in a mouse model of retinitis pigmentosa. FASEB J. 2018, 32, 2438–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Cámara, C.M.-F.; Sequedo, M.D.; Gómez-Pinedo, U.; Jaijo, T.; Aller, E.; García-Tárraga, P.; García-Verdugo, J.M.; Millán, J.M.; Rodrigo, R. Phosphodiesterase inhibition induces retinal degeneration, oxidative stress and inflammation in cone-enriched cultures of porcine retina. Exp. Eye Res. 2013, 111, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Campochiaro, P.A.; Strauss, R.W.; Lu, L.; Hafiz, G.; Wolfson, Y.; Shah, S.M.; Sophie, R.; Mir, T.A.; Scholl, H.P. Is there excess oxidative stress and damage in eyes of patients with retinitis pigmentosa? Antioxid. Redox Signal. 2015, 23, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Olivares-González, L.; Velasco, S. Retinal inflammation, cell death and inherited retinal dystrophies. Int. J. Mol. Sci. 2021, 22, 2096. [Google Scholar] [CrossRef]
- Valero-Vello, M.; Peris-Martínez, C. Searching for the antioxidant, anti-inflammatory, and neuroprotective potential of natural food and nutritional supplements for ocular health in the mediterranean population. Foods 2021, 10, 1231. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report No. 8. Arch. Ophthalmol. 2001, 119, 1417–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfonso-Muñoz, E.A.; Burggraaf-Sánchez de Las Matas, R. Role of oral antioxidant supplementation in the current management of diabetic retinopathy. Int. J. Mol. Sci. 2021, 22, 4020. [Google Scholar] [CrossRef] [PubMed]
- Piano, I.; D’Antongiovanni, V.; Testai, L.; Calderone, V.; Gargini, C. A nutraceutical strategy to slowing down the progression of cone death in an animal model of retinitis pigmentosa. Front. Neurosci. 2019, 13, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, J.K.W.; Wolf, S.A.; Pompoes, I.M.; Joussen, A.M.; Lam, W.C. Potential effects of nutraceuticals in retinopathy of prematurity. Life 2021, 11, 79. [Google Scholar] [CrossRef]
- Murakami, Y.; Nakabeppu, Y. Oxidative stress and microglial response in retinitis pigmentosa. Int. J. Mol. Sci. 2020, 21, 7170. [Google Scholar] [CrossRef]
- Cuervo, M.; Abete, I.; Baladia, E. Ingestas Dietéticas de Referencia (IDR) Para la Población Española, 1st ed.; EUNSA, Ed.; EUNSA: Barañain, Spain, 2010. (In Spanish) [Google Scholar]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Cámara, C.M.-F.; Olivares-González, L.; Hervás, D.; Salom, D.; Millán, J.M.; Rodrigo, R. Infliximab reduces Zaprinast-induced retinal degeneration in cultures of porcine retina. J. Neuroinflamm. 2014, 11, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, B.; Hawes, N.L.; Pardue, M.T.; German, A.M.; Hurd, R.E.; Davisson, M.T.; Nusinowitz, S.; Rengarajan, K.; Boyd, A.P.; Sidney, S.S.; et al. Two mouse retinal degenerations caused by missense mutations in the beta-subunit of rod cGMP phosphodiesterase gene. Vis. Res. 2007, 47, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Olivares-González, L.; Velasco, S.; Millán, J.M.; Rodrigo, R. Intravitreal administration of adalimumab delays retinal degeneration in rd10 mice. FASEB J. 2020, 34, 13839–13861. [Google Scholar] [CrossRef] [PubMed]
- Trouillet, A.; Dubus, E.; Dégardin, J.; Estivalet, A.; Ivkovic, I.; Godefroy, D.; García-Ayuso, D.; Simonutti, M.; Sahly, I.; Sahel, J.A.; et al. Cone degeneration is triggered by the absence of USH1 proteins but prevented by antioxidant treatments. Sci. Rep. 2018, 8, 1968. [Google Scholar] [CrossRef]
- Fernández-Sánchez, L.; Lax, P.; Esquiva, G.; Martín-Nieto, J.; Pinilla, I.; Cuenca, N. Safranal, a saffron constituent, attenuates retinal degeneration in P23H rats. PLoS ONE 2012, 7, e43074. [Google Scholar] [CrossRef] [Green Version]
- Jayakody, S.A.; Gonzalez-Cordero, A.; Ali, R.R.; Pearson, R.A. Cellular strategies for retinal repair by photoreceptor replacement. Prog. Retin. Eye Res. 2015, 46, 31–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, E.Y.; Clemons, T.E.; Agrón, E.; Sperduto, R.D.; Sangiovanni, J.P.; Kurinij, N.; Davis, M.D. Long-term effects of vitamins C and E, β-carotene, and zinc on age-related macular degeneration: AREDS report No. 35. Ophthalmology 2013, 120, 1604–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, E.Y.; Clemons, T.E.; Sangiovanni, J.P.; Danis, R.P.; Ferris, F.L.; Elman, M.J.; Antoszyk, A.N.; Ruby, A.J.; Orth, D.; Bressler, S.B.; et al. Secondary analyses of the effects of lutein/zeaxanthin on age-related macular degeneration progression: AREDS2 report No. 3. JAMA Ophthalmol. 2014, 132, 142–149. [Google Scholar] [CrossRef]
- Evans, J. Antioxidant supplements to prevent or slow down the progression of AMD: A systematic review and meta-analysis. Eye 2008, 22, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Camelo, S.; Latil, M. Beyond AREDS formulations, what is next for intermediate age-related macular degeneration (iAMD) treatment? Potential benefits of antioxidant and anti-inflammatory apocarotenoids as neuroprotectors. Oxid. Med. Cell. Longev. 2020, 2020, 4984927. [Google Scholar] [CrossRef]
- Wong, P.; Markey, M.; Rapp, C.M.; Darrow, R.M.; Ziesel, A.; Organisciak, D.T. Enhancing the efficacy of AREDS antioxidants in light-induced retinal degeneration. Mol. Vis. 2017, 23, 718–739. [Google Scholar]
- Saari, J.C. Vitamin A and vision. Sub-Cell. Biochem. 2016, 81, 231–259. [Google Scholar] [CrossRef]
- Bavik, C.; Henry, S.H.; Zhang, Y.; Mitts, K.; McGinn, T.; Budzynski, E.; Pashko, A.; Lieu, K.L.; Zhong, S.; Blumberg, B.; et al. Visual cycle modulation as an approach toward preservation of retinal integrity. PLoS ONE 2015, 10, e0124940. [Google Scholar] [CrossRef] [Green Version]
- Klaver, C.C.W.; Thiadens, A. Vitamin A for children with retinitis pigmentosa: An unresolved mystery. JAMA Ophthalmol. 2018, 136, 496–497. [Google Scholar] [CrossRef] [PubMed]
- Berson, E.L.; Rosner, B.; Sandberg, M.A.; Hayes, K.C.; Nicholson, B.W.; Weigel-DiFranco, C.; Willett, W. A randomized trial of vitamin A and vitamin E supplementation for retinitis pigmentosa. Arch. Ophthalmol. 1993, 111, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Radu, R.A.; Yuan, Q.; Hu, J.; Peng, J.H.; Lloyd, M.; Nusinowitz, S.; Bok, D.; Travis, G.H. Accelerated accumulation of lipofuscin pigments in the RPE of a mouse model for ABCA4-mediated retinal dystrophies following Vitamin A supplementation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3821–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi-Okai, K.; Nagino, H.; Yamada, K.; Okai, Y. Antioxidant and prooxidant activities of B group vitamins in lipid peroxidation. J. UOEH 2006, 28, 359–368. [Google Scholar] [CrossRef]
- Hsu, C.C.; Cheng, C.H.; Hsu, C.L.; Lee, W.J.; Huang, S.C.; Huang, Y.C. Role of vitamin B6 status on antioxidant defenses, glutathione, and related enzyme activities in mice with homocysteine-induced oxidative stress. Food Nutr. Res. 2015, 59, 25702. [Google Scholar] [CrossRef] [Green Version]
- Bahmani, F.; Karamali, M.; Shakeri, H.; Asemi, Z. The effects of folate supplementation on inflammatory factors and biomarkers of oxidative stress in overweight and obese women with polycystic ovary syndrome: A randomized, double-blind, placebo-controlled clinical trial. Clin. Endocrinol. 2014, 81, 582–587. [Google Scholar] [CrossRef]
- Christen, W.G.; Glynn, R.J.; Chew, E.Y.; Albert, C.M.; Manson, J.E. Folic acid, pyridoxine, and cyanocobalamin combination treatment and age-related macular degeneration in women: The women’s antioxidant and folic acid cardiovascular study. Arch. Intern. Med. 2009, 169, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Merle, B.M.; Silver, R.E.; Rosner, B.; Seddon, J.M. Dietary folate, B vitamins, genetic susceptibility and progression to advanced nonexudative age-related macular degeneration with geographic atrophy: A prospective cohort study. Am. J. Clin. Nutr. 2016, 103, 1135–1144. [Google Scholar] [CrossRef]
- Ibuki, M.; Lee, D. Rice bran and vitamin B6 suppress pathological neovascularization in a murine model of age-related macular degeneration as novel hif inhibitors. Int. J. Mol. Sci. 2020, 21, 8940. [Google Scholar] [CrossRef] [PubMed]
- Locri, F.; Cammalleri, M.; Dal Monte, M. Protective efficacy of a dietary supplement based on forskolin, homotaurine, spearmint extract, and group B vitamins in a mouse model of optic nerve injury. Nutrients 2019, 11, 2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammalleri, M.; Dal Monte, M.; Amato, R.; Bagnoli, P.; Rusciano, D. A dietary combination of forskolin with homotaurine, spearmint and B vitamins protects injured retinal ganglion cells in a rodent model of hypertensive glaucoma. Nutrients 2020, 12, 1189. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.W.; Li, Q.; Zhang, J.Z.; Zhang, Y.M.; Liu, Y.; Yang, K.H. The protective roles of folic acid in preventing diabetic retinopathy are potentially associated with suppressions on angiogenesis, inflammation, and oxidative stress. Ophthalmic Res. 2019, 62, 80–92. [Google Scholar] [CrossRef]
- Stelling, M.P.; Motta, J.M.; Mashid, M.; Johnson, W.E.; Pavão, M.S. Metal ions and the extracellular matrix in tumor migration. FEBS J. 2019, 286, 2950–2964. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Bang, E.S.; Lee, J.H.; Lee, J.D.; Kang, D.R.; Hong, J.; Lee, J.M. Serum concentrations of trace elements zinc, copper, selenium, and manganese in critically Ill patients. Biol. Trace Element Res. 2019, 188, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Salmonowicz, B.; Krzystek-Korpacka, M.; Noczyńska, A. Trace elements, magnesium, and the efficacy of antioxidant systems in children with type 1 diabetes mellitus and in their siblings. Adv. Clin. Exp. Med. 2014, 23, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Semeraro, F.; Gambicordi, E.; Cancarini, A.; Morescalchi, F.; Costagliola, C.; Russo, A. Treatment of exudative age-related macular degeneration with aflibercept combined with pranoprofen eye drops or nutraceutical support with omega-3: A randomized trial. Br. J. Clin. Pharmacol. 2019, 85, 908–913. [Google Scholar] [CrossRef]
- Bell, S.G.; Vallee, B.L. The metallothionein/thionein system: An oxidoreductive metabolic zinc link. Chembiochem 2009, 10, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.R.; Stillman, M.J. Metal-dependent protein folding: Metallation of metallothionein. J. Inorg. Biochem. 2006, 100, 2101–2107. [Google Scholar] [CrossRef]
- Kamińska, A.; Romano, G.L. Influence of trace elements on neurodegenerative diseases of the eye—The glaucoma model. Int. J. Mol. Sci. 2021, 22, 4323. [Google Scholar] [CrossRef] [PubMed]
- Ugarte, M.; Osborne, N.N. Recent advances in the understanding of the role of zinc in ocular tissues. Metallomics 2014, 6, 189–200. [Google Scholar] [CrossRef]
- Tan, J.S.; Wang, J.J.; Flood, V.; Rochtchina, E.; Smith, W.; Mitchell, P. Dietary antioxidants and the long-term incidence of age-related macular degeneration: The blue mountains eye study. Ophthalmology 2008, 115, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Duntas, L.H. Selenium and inflammation: Underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009, 41, 443–447. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, I.S.; Murphy, M.E. Type-2 copper-containing enzymes. Cell. Mol. Life Sci. 2007, 64, 2887–2899. [Google Scholar] [CrossRef]
- Ugarte, M.; Osborne, N.N.; Brown, L.A.; Bishop, P.N. Iron, zinc, and copper in retinal physiology and disease. Surv. Ophthalmol. 2013, 58, 585–609. [Google Scholar] [CrossRef]
- Zampatti, S.; Ricci, F.; Cusumano, A.; Marsella, L.T.; Novelli, G.; Giardina, E. Review of nutrient actions on age-related macular degeneration. Nutr. Res. 2014, 34, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B.; López-Pozo, M.; Stewart, J.J. Zeaxanthin and lutein: Photoprotectors, anti-inflammatories, and brain food. Molecules 2020, 25, 3607. [Google Scholar] [CrossRef]
- Sandberg, M.A.; Johnson, E.J.; Berson, E.L. The relationship of macular pigment optical density to serum lutein in retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1086–1091. [Google Scholar] [CrossRef]
- Yu, M.; Yan, W.; Beight, C. Lutein and zeaxanthin isomers reduce photoreceptor degeneration in the Pde6b (rd10) mouse model of retinitis pigmentosa. BioMed Res. Int. 2018, 2018, 4374087. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Yan, W.; Beight, C. Lutein and zeaxanthin isomers protect against light-induced retinopathy via decreasing oxidative and endoplasmic reticulum stress in BALB/cJ mice. Nutrients 2018, 10, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozawa, Y.; Sasaki, M.; Takahashi, N.; Kamoshita, M.; Miyake, S.; Tsubota, K. Neuroprotective effects of lutein in the retina. Curr. Pharm. Des. 2012, 18, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Shahzad, T.; Malik, A.; Shariati, M.A.; Laishevtcev, A.; Plygun, S.; Heydari, M.; et al. Xanthophyll: Health benefits and therapeutic insights. Life Sci. 2020, 240, 117104. [Google Scholar] [CrossRef]
- Berson, E.L.; Rosner, B.; Sandberg, M.A.; Weigel-DiFranco, C.; Brockhurst, R.J.; Hayes, K.C.; Johnson, E.J.; Anderson, E.J.; Johnson, C.A.; Gaudio, A.R.; et al. Clinical trial of lutein in patients with retinitis pigmentosa receiving vitamin A. Arch. Ophthalmol. 2010, 128, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, H.; Melia, M.; Dagnelie, G. Lutein supplementation in retinitis pigmentosa: PC-based vision assessment in a randomized double-masked placebo-controlled clinical trial [NCT00029289]. BMC Ophthalmol. 2006, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotenstreich, Y.; Belkin, M.; Sadetzki, S.; Chetrit, A.; Ferman-Attar, G.; Sher, I.; Harari, A.; Shaish, A.; Harats, D. Treatment with 9-cis β-carotene-rich powder in patients with retinitis pigmentosa: A randomized crossover trial. JAMA Ophthalmol. 2013, 131, 985–992. [Google Scholar] [CrossRef] [Green Version]
- Gombart, A.F.; Pierre, A.; Maggini, S. A review of micronutrients and the immune system-working in harmony to reduce the risk of infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef] [Green Version]
- Gunal, M.Y.; Sakul, A.A. Protective effect of lutein/zeaxanthin isomers in traumatic brain injury in mice. Neurotox. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Vellozo, N.S.; Pereira-Marques, S.T.; Cabral-Piccin, M.P.; Filardy, A.A.; Ribeiro-Gomes, F.L.; Rigoni, T.S.; DosReis, G.A.; Lopes, M.F. All-trans retinoic acid promotes an M1- to M2-phenotype shift and inhibits macrophage-mediated immunity to leishmania major. Front. Immunol. 2017, 8, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.M.; Teixeira, F.M.E. Impact of retinoic acid on immune cells and inflammatory diseases. Mediat. Inflamm. 2018, 2018, 3067126. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.; Peto, T.; Lengyel, I.; Emri, E. Zinc nutrition and inflammation in the aging retina. Mol. Nutr. Food Res. 2019, 63, e1801049. [Google Scholar] [CrossRef] [PubMed]
- Smailhodzic, D.; van Asten, F.; Blom, A.M.; Mohlin, F.C.; den Hollander, A.I.; van de Ven, J.P.; van Huet, R.A.; Groenewoud, J.M.; Tian, Y.; Berendschot, T.T.; et al. Zinc supplementation inhibits complement activation in age-related macular degeneration. PLoS ONE 2014, 9, e112682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana-Gálvez, J.; Villela-Castrejón, J. Synergistic combinations of curcumin, sulforaphane, and dihydrocaffeic acid against human colon cancer cells. Int. J. Mol. Sci. 2020, 21, 3108. [Google Scholar] [CrossRef]
- Veurink, G.; Perry, G.; Singh, S.K. Role of antioxidants and a nutrient rich diet in Alzheimer’s disease. Open Biol. 2020, 10, 200084. [Google Scholar] [CrossRef]
- Zhang, L.; McClements, D.J.; Wei, Z.; Wang, G.; Liu, X.; Liu, F. Delivery of synergistic polyphenol combinations using biopolymer-based systems: Advances in physicochemical properties, stability and bioavailability. Crit. Rev. Food Sci. Nutr. 2019, 60, 2083–2097. [Google Scholar] [CrossRef]
- Shi, C.; Wang, P.; Airen, S.; Brown, C.; Liu, Z.; Townsend, J.H.; Wang, J.; Jiang, H. Nutritional and medical food therapies for diabetic retinopathy. Eye Vis. 2020, 7, 1–16. [Google Scholar] [CrossRef]
- Leena, M.M.; Silvia, M.G.; Vinitha, K.; Moses, J.A.; Anandharamakrishnan, C. Synergistic potential of nutraceuticals: Mechanisms and prospects for futuristic medicine. Food Funct. 2020, 11, 9317–9337. [Google Scholar] [CrossRef]
- Gargini, C.; Terzibasi, E.; Mazzoni, F.; Strettoi, E. Retinal organization in the retinal degeneration 10 (rd10) mutant mouse: A morphological and ERG study. J. Comp. Neurol. 2007, 500, 222–238. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yin, Z.; Gao, L.; Sun, D.; Hu, X.; Xue, L.; Dai, J.; Zeng, Y.; Chen, S.; Pan, B.; et al. Curcumin delays retinal degeneration by regulating microglia activation in the retina of rd1 mice. Cell. Physiol. Biochem. 2017, 44, 479–493. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, K. Vitamins and mineral supplements for retinitis pigmentosa. J. Ophthalmol. 2019, 2019, 8524607. [Google Scholar] [CrossRef]
- Ramírez-Lamelas, D.T.; Benlloch-Navarro, S.; López-Pedrajas, R.; Gimeno-Hernández, R.; Olivar, T.; Silvestre, D.; Miranda, M. Lipoic acid and progesterone alone or in combination ameliorate retinal degeneration in an experimental model of hereditary retinal degeneration. Front. Pharmacol. 2018, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Robles-Rivera, R.R.; Castellanos-González, J.A.; Olvera-Montaño, C.; Flores-Martin, R.A.; López-Contreras, A.K.; Arevalo-Simental, D.E.; Cardona-Muñoz, E.G. Adjuvant therapies in diabetic retinopathy as an early approach to delay its progression: The importance of oxidative stress and inflammation. Oxid. Med. Cell. Longev. 2020, 2020, 3096470. [Google Scholar] [CrossRef]
- Tokarz, P.; Kaarniranta, K.; Blasiak, J. Role of antioxidant enzymes and small molecular weight antioxidants in the pathogenesis of age-related macular degeneration (AMD). Biogerontology 2013, 14, 461–482. [Google Scholar] [CrossRef] [Green Version]
- Ahmadinejad, F.; Geir Møller, S.; Hashemzadeh-Chaleshtori, M.; Bidkhori, G.; Jami, M.S. Molecular mechanisms behind free radical scavengers function against oxidative stress. Antioxidants 2017, 6, 51. [Google Scholar] [CrossRef]
- Skourtis, G.; Krontira, A.; Ntaoula, S.; Ferlemi, A.V.; Zeliou, K.; Georgakopoulos, C.; Margarity, G.M.; Lamari, N.F.; Pharmakakis, N. Protective antioxidant effects of saffron extract on retinas of streptozotocin-induced diabetic rats. Rom. J. Ophthalmol. 2020, 64, 394–403. [Google Scholar] [CrossRef] [PubMed]
- King, R.E.; Kent, K.D.; Bomser, J.A. Resveratrol reduces oxidation and proliferation of human retinal pigment epithelial cells via extracellular signal-regulated kinase inhibition. Chemico-Biol. Interact. 2005, 151, 143–149. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Z.Z.; Cheng, Y.L.; Lin, W.; Qu, C. Resveratrol protects against oxidative damage of retinal pigment epithelium cells by modulating SOD/MDA activity and activating Bcl-2 expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 378–388. [Google Scholar] [CrossRef]
- Delmas, D.; Cornebise, C.; Courtaut, F.; Xiao, J. New highlights of resveratrol: A review of properties against ocular diseases. Int. J. Mol. Sci. 2021, 22, 1295. [Google Scholar] [CrossRef]
- Yu, C.C.; Nandrot, E.F.; Dun, Y.; Finnemann, S.C. Dietary antioxidants prevent age-related retinal pigment epithelium actin damage and blindness in mice lacking αvβ5 integrin. Free Radic. Biol. Med. 2012, 52, 660–670. [Google Scholar] [CrossRef] [Green Version]
- Stence, N.; Waite, M.; Dailey, M.E. Dynamics of microglial activation: A confocal time-lapse analysis in hippocampal slices. Glia 2001, 33, 256–266. [Google Scholar] [CrossRef]
- Michelucci, A.; Heurtaux, T.; Grandbarbe, L.; Morga, E.; Heuschling, P. Characterization of the microglial phenotype under specific pro-inflammatory and anti-inflammatory conditions: Effects of oligomeric and fibrillar amyloid-beta. J. Neuroimmunol. 2009, 210, 3–12. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, J.Y.; Kim, J.Y.; Park, J.; Lee, W.T.; Lee, J.E. M2 phenotype microglia-derived cytokine stimulates proliferation and neuronal differentiation of endogenous stem cells in ischemic brain. Exp. Neurobiol. 2017, 26, 33–41. [Google Scholar] [CrossRef]
- Chhor, V.; Le Charpentier, T.; Lebon, S.; Oré, M.V.; Celador, I.L.; Josserand, J.; Degos, V.; Jacotot, E.; Hagberg, H.; Sävman, K.; et al. Characterization of phenotype markers and neuronotoxic potential of polarised primary microglia in vitro. Brain Behav. Immun. 2013, 32, 70–85. [Google Scholar] [CrossRef]
- Peña-Altamira, E.; Petralla, S.; Massenzio, F.; Virgili, M.; Bolognesi, M.L.; Monti, B. Nutritional and pharmacological strategies to regulate microglial polarization in cognitive aging and Alzheimer’s disease. Front. Aging Neurosci. 2017, 9, 175. [Google Scholar] [CrossRef]
- Ebert, S.; Weigelt, K.; Walczak, Y.; Drobnik, W.; Mauerer, R.; Hume, D.A.; Weber, B.H.; Langmann, T. Docosahexaenoic acid attenuates microglial activation and delays early retinal degeneration. J. Neurochem. 2009, 110, 1863–1875. [Google Scholar] [CrossRef]
- Wang, W.; Tam, K.C.; Ng, T.C.; Goit, R.K.; Chan, K.L.S.; Lo, A.C.Y. Long-term lutein administration attenuates retinal inflammation and functional deficits in early diabetic retinopathy using the Ins2(Akita/+) mice. BMJ Open Diabetes Res. Care 2020, 8, e001519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, Y.; Luo, Y.; Du, Y.; Zhang, X.; Fu, J. Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/ TLR4/ NF-κB pathways in BV2 cells. Mol. Immunol. 2019, 116, 29–37. [Google Scholar] [CrossRef]
- Fernández-Albarral, J.A.; Ramírez, A.I.; de Hoz, R.; López-Villarín, N.; Salobrar-García, E. Neuroprotective and anti-inflammatory effects of a hydrophilic saffron extract in a model of glaucoma. Int. J. Mol. Sci. 2019, 20, 4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosado, M.M.; Bennici, E.; Novelli, F.; Pioli, C. Beyond DNA repair, the immunological role of PARP-1 and its siblings. Immunology 2013, 139, 428–437. [Google Scholar] [CrossRef]
- Sinha, A.; Katyal, S.; Kauppinen, T.M. PARP-DNA trapping ability of PARP inhibitors jeopardizes astrocyte viability: Implications for CNS disease therapeutics. Neuropharmacology 2021, 187, 108502. [Google Scholar] [CrossRef]
- Sahaboglu, A.; Barth, M.; Secer, E.; Amo, E.M.; Urtti, A.; Arsenijevic, Y.; Zrenner, E.; Paquet-Durand, F. Olaparib significantly delays photoreceptor loss in a model for hereditary retinal degeneration. Sci. Rep. 2016, 6, 39537. [Google Scholar] [CrossRef] [Green Version]
- Carullo, G.; Federico, S.; Relitti, N. Retinitis pigmentosa and retinal degenerations: Deciphering pathways and targets for drug discovery and development. ACS Chem. Neurosci. 2020, 11, 2173–2191. [Google Scholar] [CrossRef]
- Raghunatha, P.; Vosoughi, A.; Kauppinen, T.M. Microglial NMDA receptors drive pro-inflammatory responses via PARP-1/TRMP2 signaling. Glia 2020, 68, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Correani, V.; Martire, S.; Mignogna, G.; Caruso, L.B.; Tempera, I.; Giorgi, A.; Grieco, M.; Mosca, L.; Schininà, M.E.; Maras, B.; et al. Poly(ADP-ribosylated) proteins in β-amyloid peptide-stimulated microglial cells. Biochem. Pharmacol. 2019, 167, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, M.; Zyma, M.; Robaszkiewicz, A. The role of PARP1 in monocyte and macrophage commitment and specification: Future Perspectives and limitations for the treatment of monocyte and macrophage relevant diseases with PARP inhibitors. Cell 2020, 9, 2040. [Google Scholar] [CrossRef]
- Connor, W.E.; Duell, P.B.; Kean, R.; Wang, Y. The prime role of HDL to transport lutein into the retina: Evidence from HDL-deficient WHAM chicks having a mutant ABCA1 transporter. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4226–4231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, B.; Natoli, S.; Liew, G.; Flood, V.M. Lutein and zeaxanthin-food sources, bioavailability and dietary variety in age-related macular degeneration protection. Nutrients 2017, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Norkus, E.P.; Norkus, K.L.; Dharmarajan, T.S.; Schierle, J.; Schalch, W. Serum lutein response is greater from free lutein than from esterified lutein during 4 weeks of supplementation in healthy adults. J. Am. Coll. Nutr. 2010, 29, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Beck, M.; Elliott, J.; Etheve, S.; Roberts, R.; Schalch, W. Effects of formulation on the bioavailability of lutein and zeaxanthin: A randomized, double-blind, cross-over, comparative, single-dose study in healthy subjects. Eur. J. Nutr. 2013, 52, 1381–1391. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; McPartlin, J.; Goggins, M.; Weir, D.G.; Scott, J.M. Unmetabolized folic acid in serum: Acute studies in subjects consuming fortified food and supplements. Am. J. Clin. Nutr. 1997, 65, 1790–1795. [Google Scholar] [CrossRef]
- Milman, N. Intestinal absorption of folic acid—New physiologic & molecular aspects. Indian J. Med Res. 2012, 136, 725–728. [Google Scholar]
- Naderi, N.; House, J.D. Recent developments in folate nutrition. Adv. Food Nutr. Res. 2018, 83, 195–213. [Google Scholar] [CrossRef]
- Reboul, E. Absorption of vitamin A and carotenoids by the enterocyte: Focus on transport proteins. Nutrients 2013, 5, 3563–3581. [Google Scholar] [CrossRef] [Green Version]
- Perusek, L.; Maeda, T. Vitamin A derivatives as treatment options for retinal degenerative diseases. Nutrients 2013, 5, 2646–2666. [Google Scholar] [CrossRef] [Green Version]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [Green Version]
- Maares, M.; Haase, H. A guide to human zinc absorption: General overview and recent advances of in vitro intestinal models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [Green Version]
- Zerounian, N.R.; Redekosky, C.; Malpe, R.; Linder, M.C. Regulation of copper absorption by copper availability in the Caco-2 cell intestinal model. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G739–G747. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients | Components | Composition |
|---|---|---|
| Macronutrients | Crude protein | 14.3% |
| Fat | 4.0% | |
| Carbohydrate | 48.0% | |
| Crude fiber | 4.1% | |
| Neutral Detergent fiber | 18.0% | |
| Ash | 4.7% | |
| Minerals | Calcium | 0.7% |
| Phosphorus | 0.6% | |
| Sodium | 0.1% | |
| Potassium | 0.6% | |
| Chloride | 0.3% | |
| Magnesium | 0.2% | |
| Zinc | 70 mg/kg | |
| Manganese | 100 mg/kg | |
| Copper | 15 mg/kg | |
| Iodine | 6 mg/kg | |
| Iron | 175 mg/kg | |
| Selenium | 0.23 mg/kg | |
| Vitamins | Vitamin A | 6.0 IU/g |
| Vitamin D3 | 0.6 IU/g | |
| Vitamin E | 120 IU/kg | |
| Vitamin K3 | 20 mg/kg | |
| Vitamin B1 | 12 mg/kg | |
| Vitamin B2 | 6 mg/kg | |
| Niacin | 54 mg/kg | |
| Vitamin B6 | 10 mg/kg | |
| Pantothenic acid | 17 mg/kg | |
| Vitamin B12 | 0.03 mg/kg | |
| Biotin | 0.26 mg/kg | |
| Folate (folic acid) | 2 mg/kg | |
| Choline | 1030 mg/kg |
| Gene | C Relative Expression (Mean ± SEM) | rd10 Relative Expression (Mean ± SEM) | rd10 + NUT Relative Expression (Mean ± SEM) |
|---|---|---|---|
| TNFα | 1.0 ± 0.0 | 40.8 ± 8.0 *** | 5.8 ± 1.4 * |
| TNFR1 | 1.0 ± 0.1 | 2.3 ± 0.4 * | 0.9 ± 0.2 bbb |
| IL-6 | 1.0 ± 0.0 | 7.8 ± 2.0 ** | 2.5 ± 0.6 bb |
| IL-1β | 1.0 ± 0.0 | 17.0 ± 3.0 *** | 3.6 ± 0.8 bb |
| IL-18 | 1.0 ± 0.1 | 0.4 ± 0.1 * | 0.1 ± 0.0 *** |
| GFAP | 1.0 ± 0.0 | 8.3 ± 1.0 *** | 2.6 ± 0.5 **b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivares-González, L.; Velasco, S.; Campillo, I.; Salom, D.; González-García, E.; Soriano del Castillo, J.M.; Rodrigo, R. Nutraceutical Supplementation Ameliorates Visual Function, Retinal Degeneration, and Redox Status in rd10 Mice. Antioxidants 2021, 10, 1033. https://doi.org/10.3390/antiox10071033
Olivares-González L, Velasco S, Campillo I, Salom D, González-García E, Soriano del Castillo JM, Rodrigo R. Nutraceutical Supplementation Ameliorates Visual Function, Retinal Degeneration, and Redox Status in rd10 Mice. Antioxidants. 2021; 10(7):1033. https://doi.org/10.3390/antiox10071033
Chicago/Turabian StyleOlivares-González, Lorena, Sheyla Velasco, Isabel Campillo, David Salom, Emilio González-García, José Miguel Soriano del Castillo, and Regina Rodrigo. 2021. "Nutraceutical Supplementation Ameliorates Visual Function, Retinal Degeneration, and Redox Status in rd10 Mice" Antioxidants 10, no. 7: 1033. https://doi.org/10.3390/antiox10071033
APA StyleOlivares-González, L., Velasco, S., Campillo, I., Salom, D., González-García, E., Soriano del Castillo, J. M., & Rodrigo, R. (2021). Nutraceutical Supplementation Ameliorates Visual Function, Retinal Degeneration, and Redox Status in rd10 Mice. Antioxidants, 10(7), 1033. https://doi.org/10.3390/antiox10071033