
High-Carbohydrate Diet Alleviates the Oxidative Stress, Inflammation and Apoptosis of Megalobrama amblycephala Following Dietary Exposure to Silver Nanoparticles


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Silver Nanoparticles Preparation
2.3. The Experimental Diets and Feeding Trial
2.4. Sample Collection
2.5. Analysis of Proximate Composition and Plasma and Liver Biochemical Indices
2.6. Analysis of Liver Histology and ATP and AMP Contents
2.7. Analysis of Western Blot (WB) and RT-PCR
2.8. Statistical Analyses
3. Results
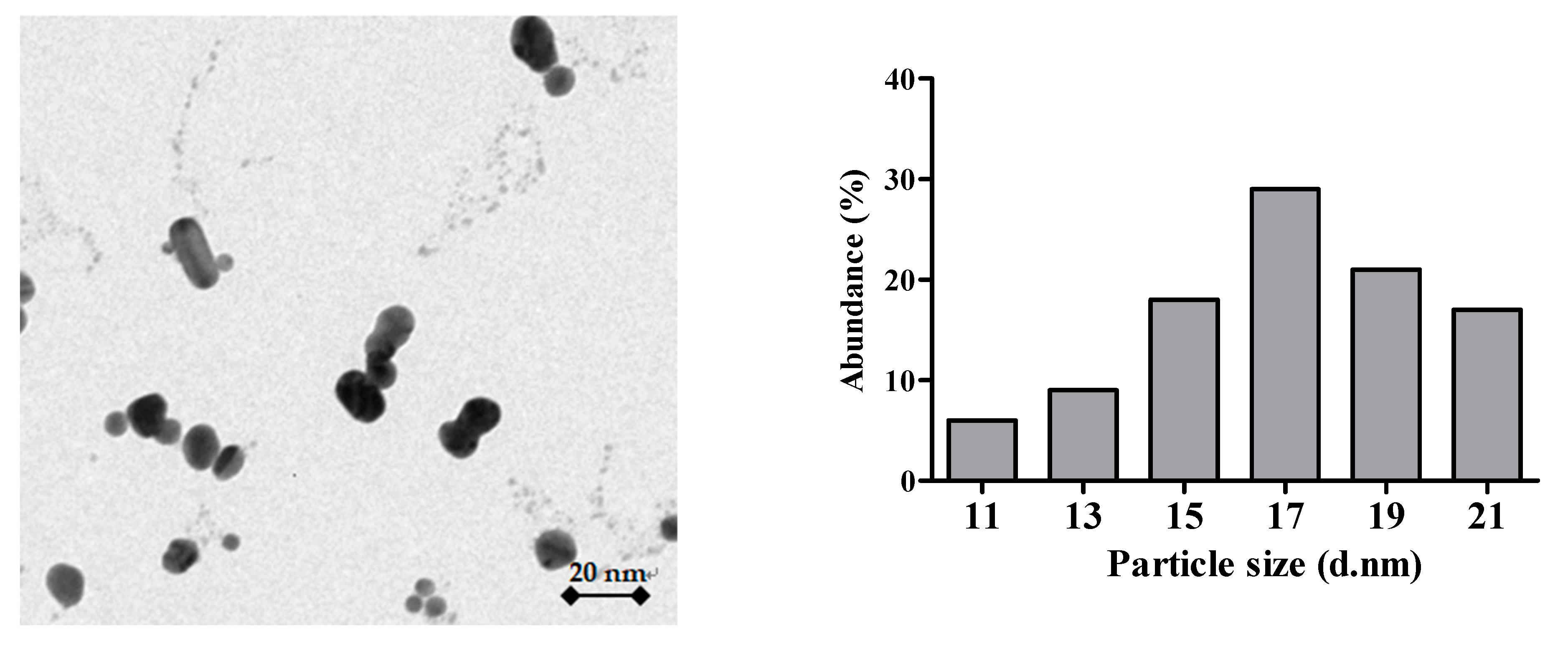
3.1. Characterization of the Ag-NPs
3.2. Growth Performance and Feed Utilization
3.3. Plasma and Liver Biochemistry Parameters
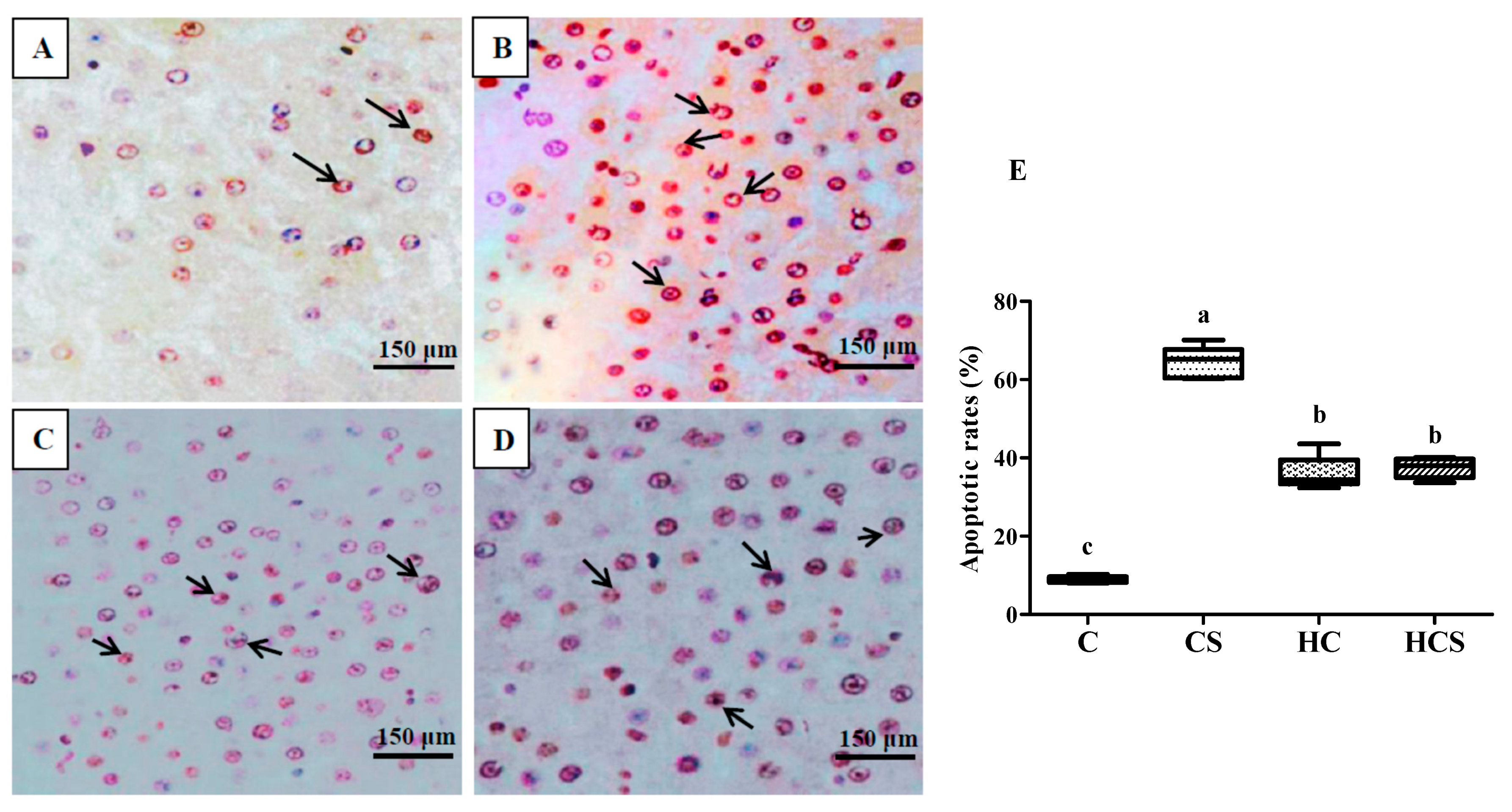
3.4. Liver Histological Analysis
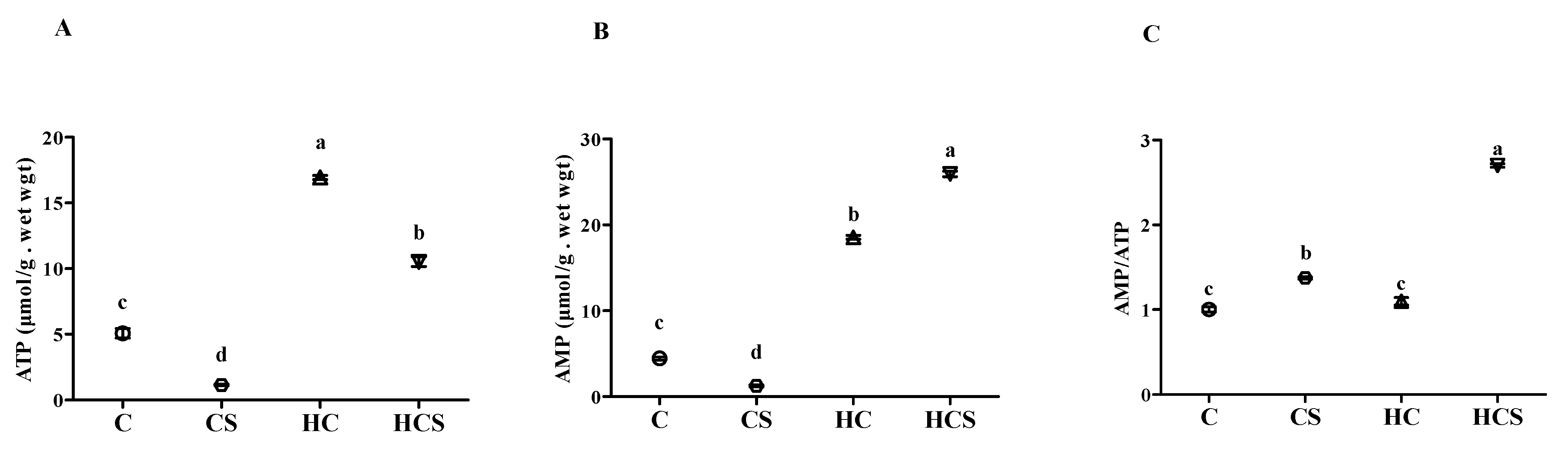
3.5. Liver ATP and AMP Contents
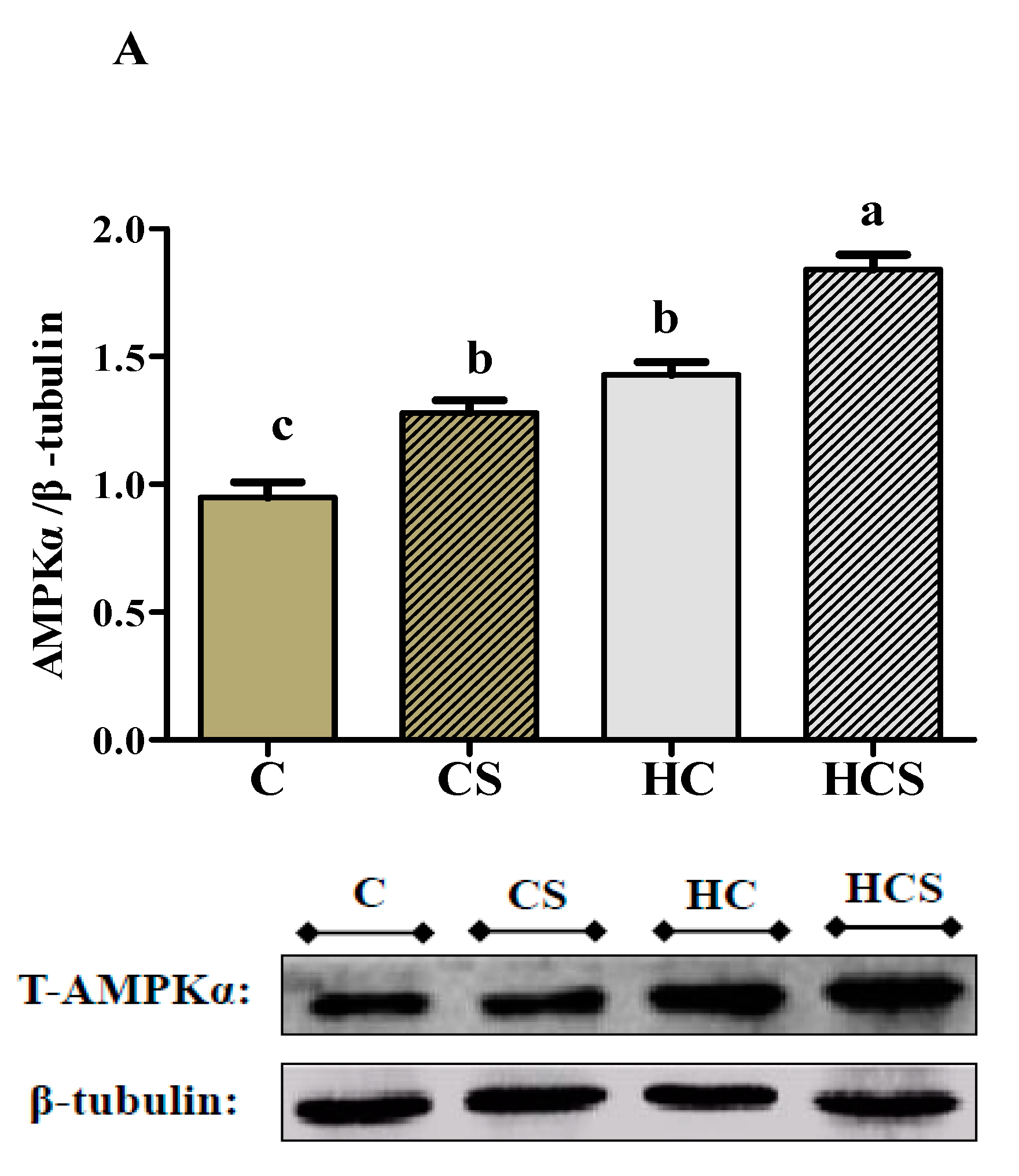
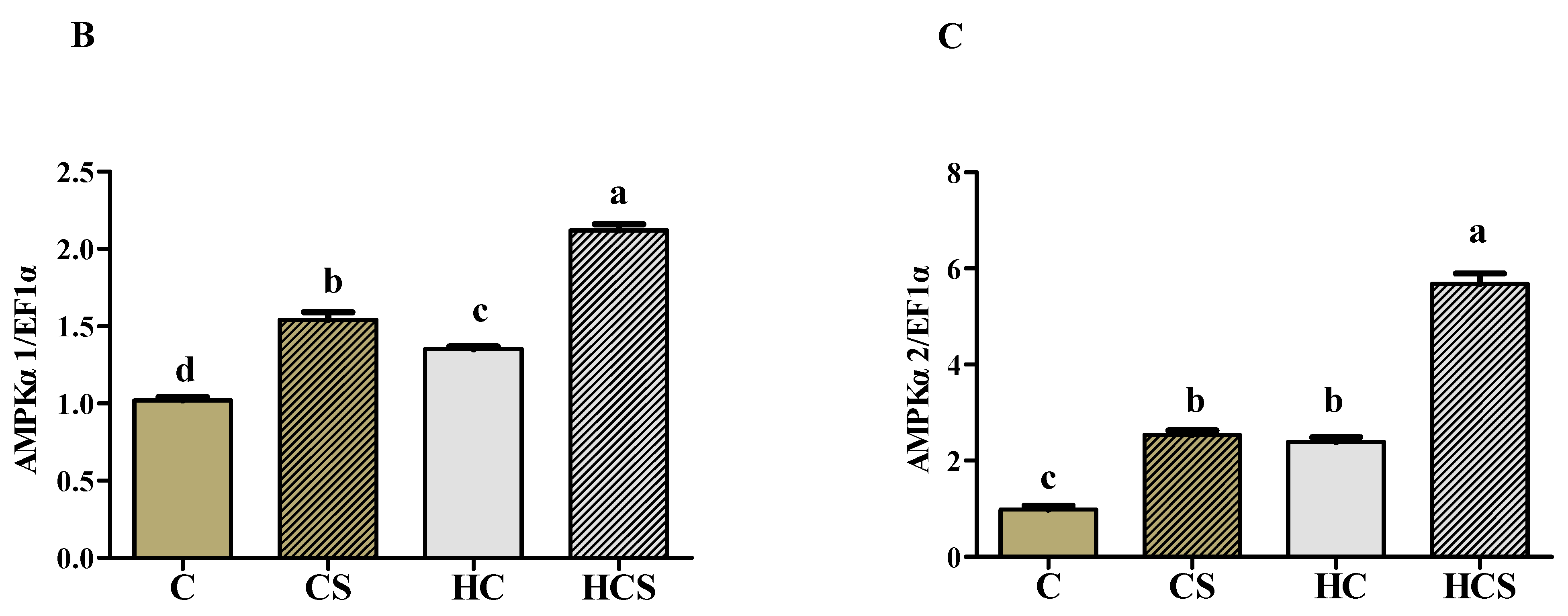
3.6. Protein Contents and Transcriptions of AMPKα
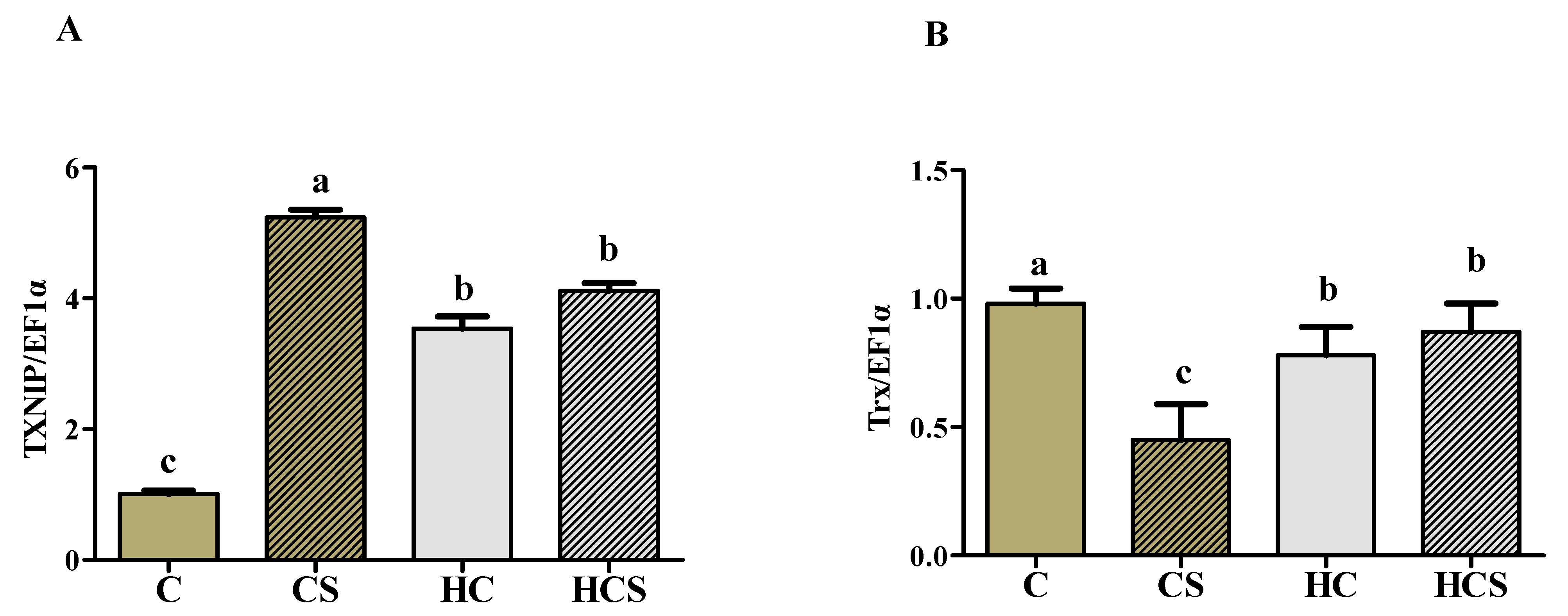
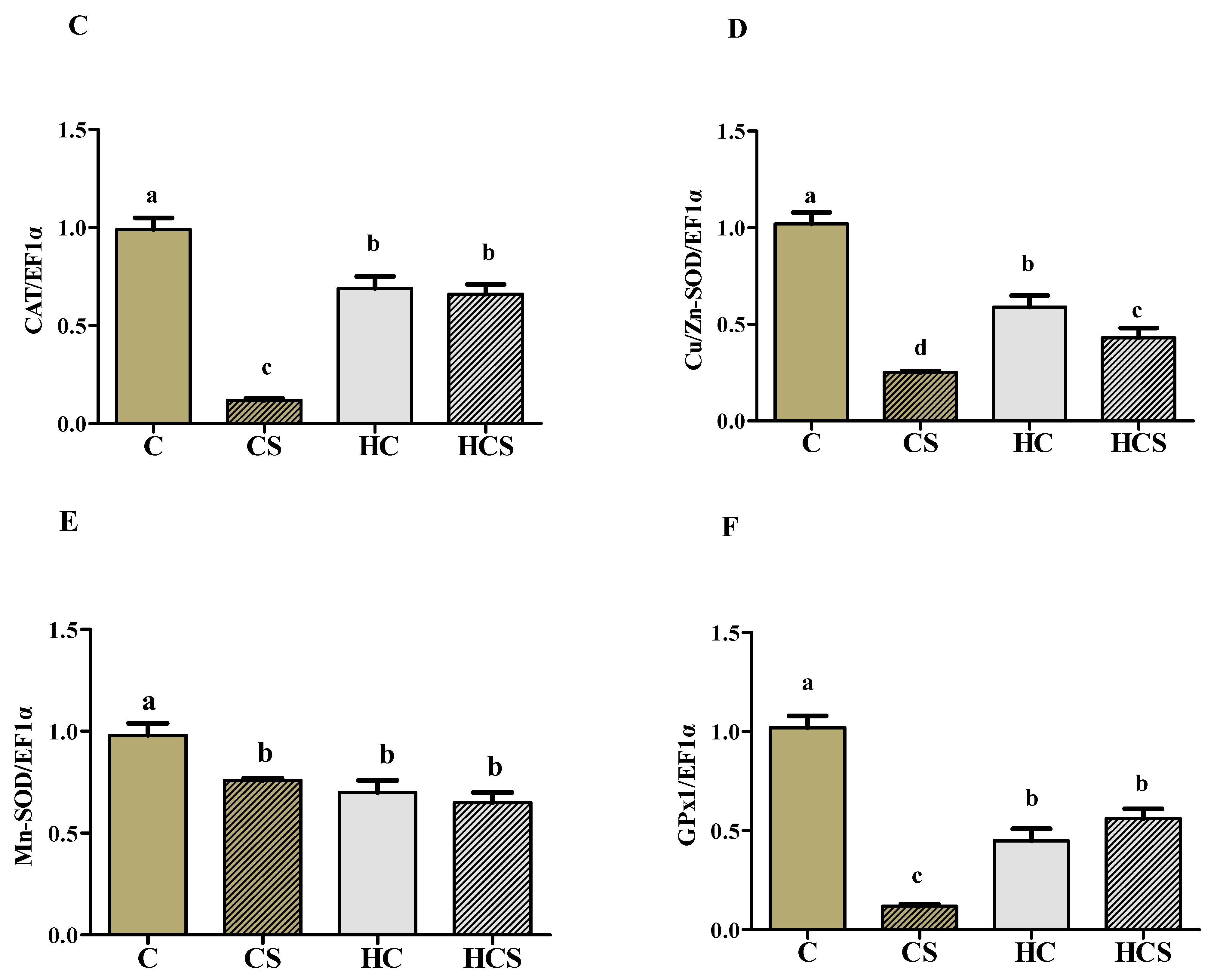
3.7. Transcriptions of Antioxidant-Related Genes
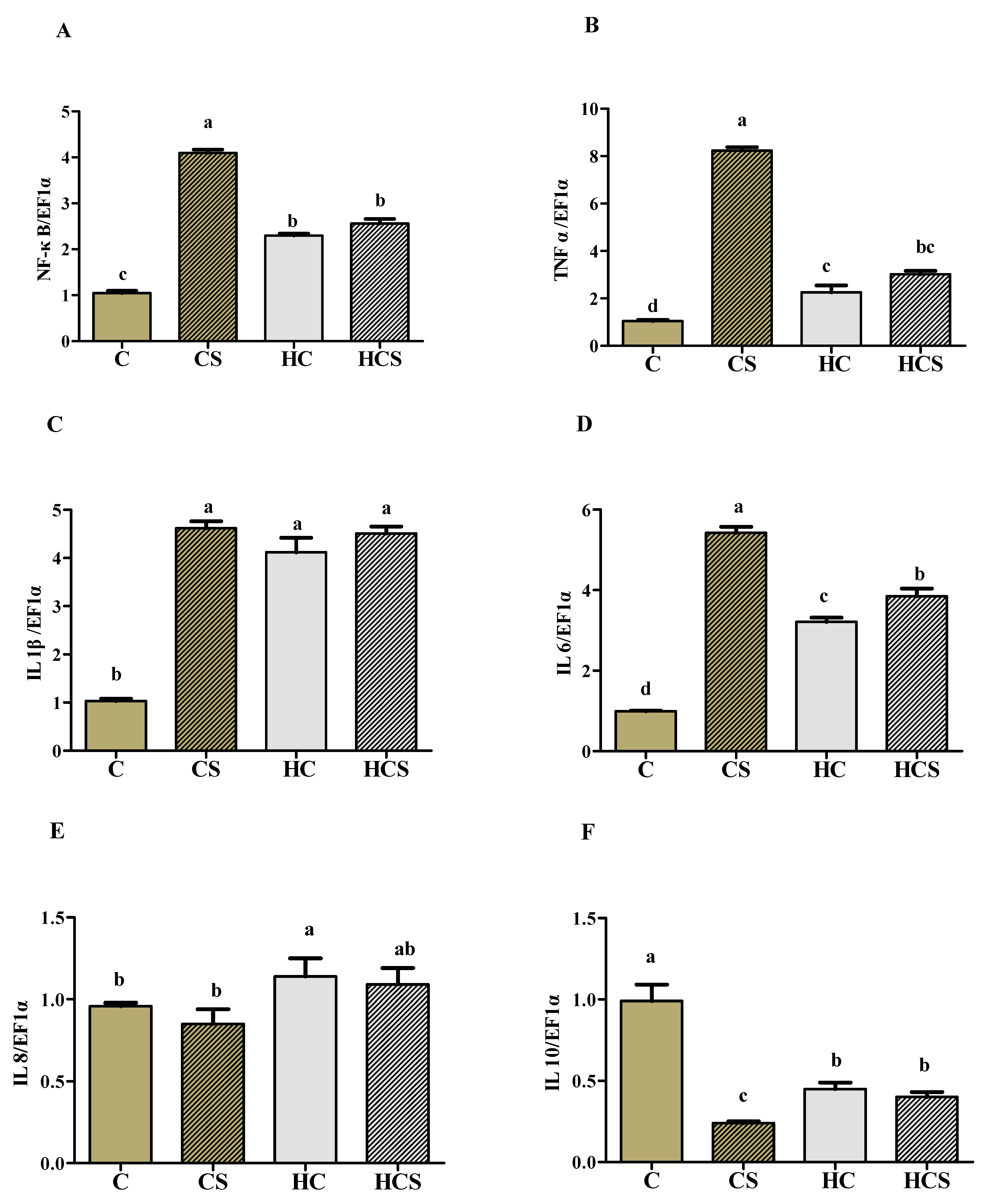
3.8. Transcriptions of Inflammation-Related Genes
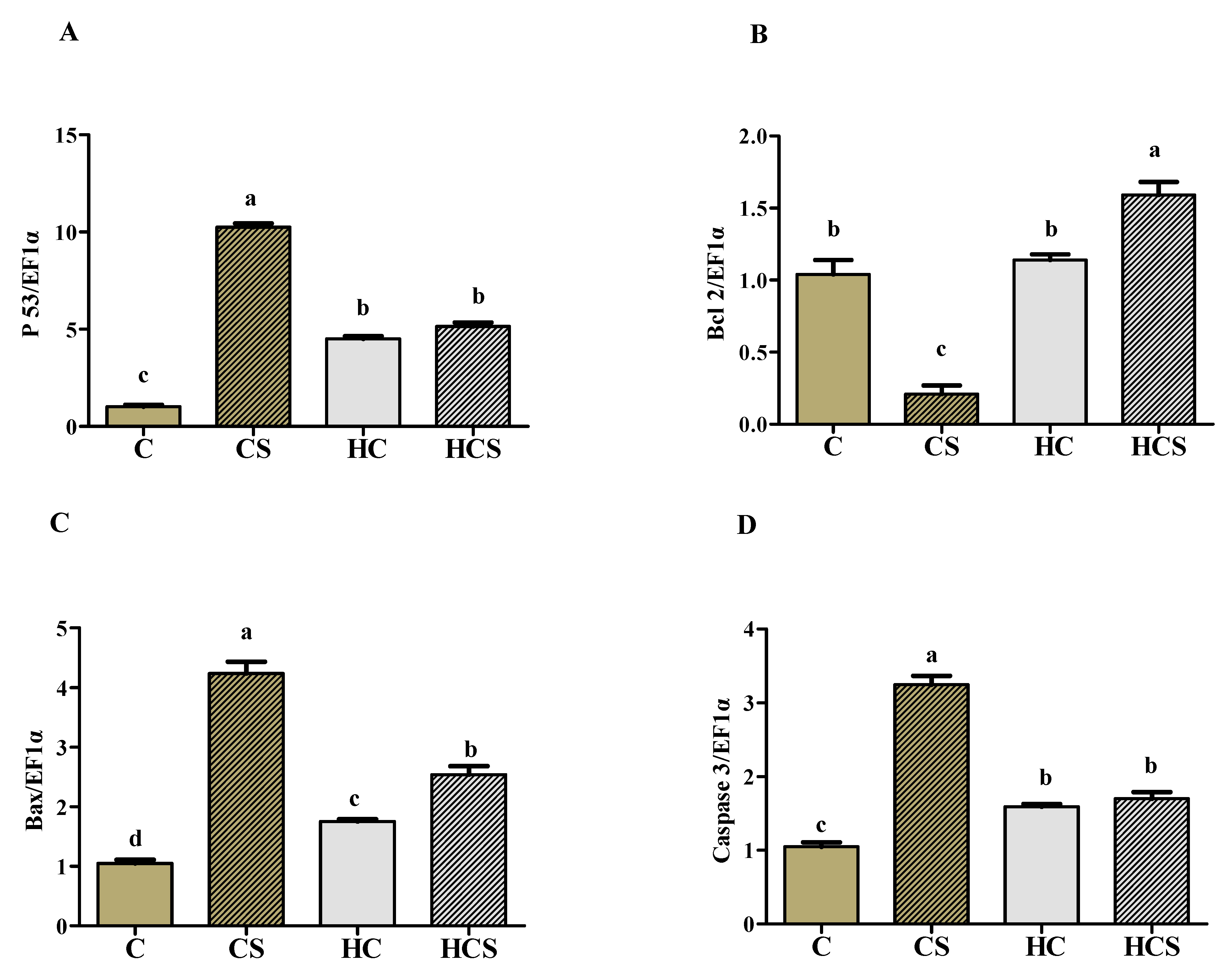
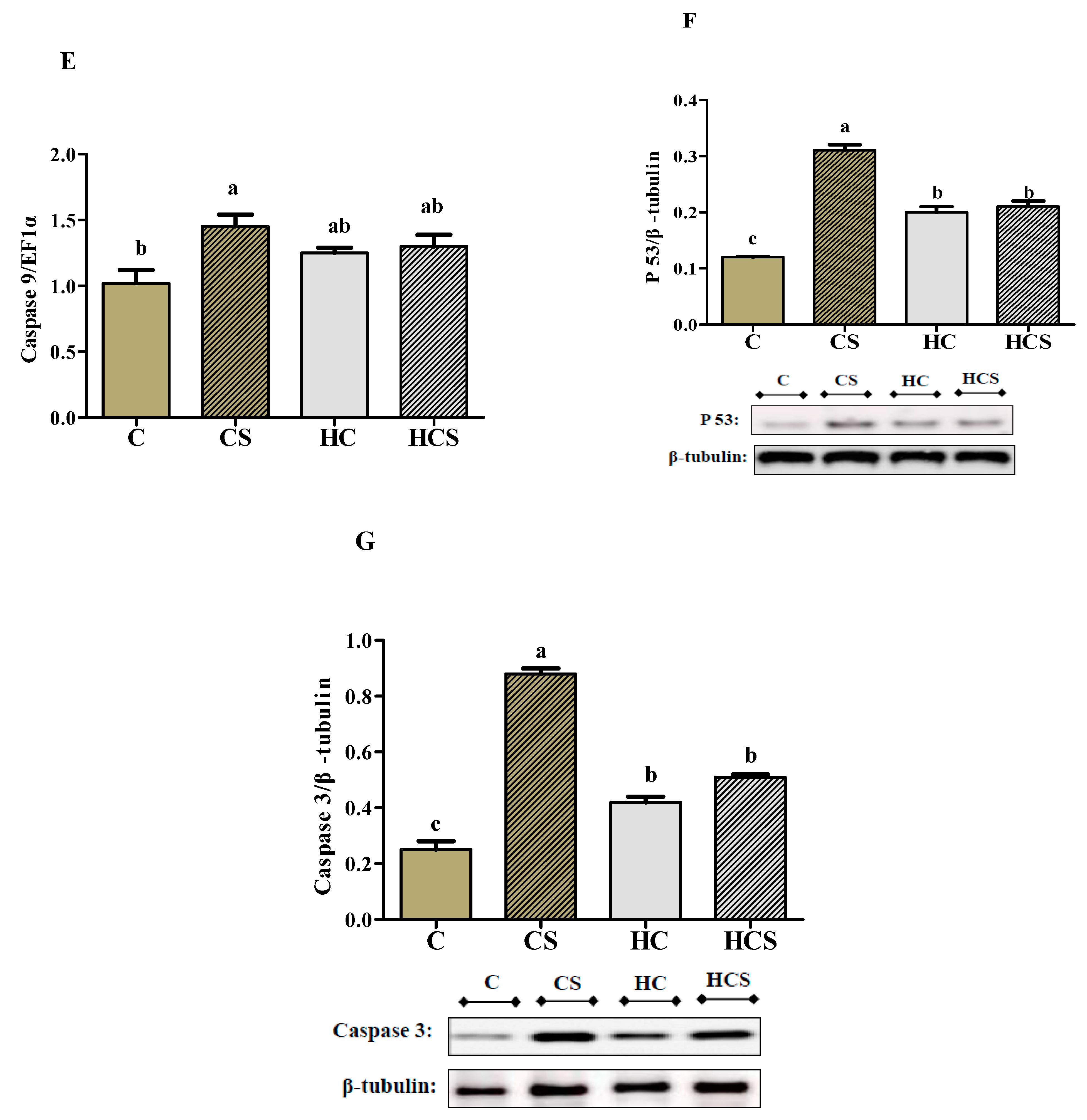
3.9. Transcriptions of Apoptosis-Related Genes and Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Du, J.; Tang, J.; Xu, S.; Ge, J.; Dong, Y.; Li, H.; Jin, M. A review on silver nanoparticles-induced ecotoxicity and the underlying toxicity mechanisms. Regul. Toxicol. Pharmacol. 2018, 98, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Q.F. Silver nanoparticles cause oxidative damage and histological changes in medaka (Oryzias latipes) after 14 days of exposure. Environ. Toxicol. Chem. 2013, 32, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.; Christian, P.; Urrea, J.A.; Roos, N.; Hassellöv, M.; Tollefsen, K.E.; Thomas, K.V. Effects of silver and gold nanoparticles on rainbow trout (Oncorhynchus mykiss) hepatocytes. Aquat. Toxicol. 2010, 96, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Asharani, P.V.; Wu, Y.L.; Gong, Z.Y.; Valiyaveettil, S. Toxicity of silver nanoparticles in zebrafish models. Nanotechnology 2008, 19, 255102. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, S.; Tripathy, D.; Mohanty, P.; Ghanim, H.; Syed, T.; Aljada, A.; Dandona, P. Differential effects of glucose and alcohol on reactive oxygen species generation and intranuclear nuclear factor-kappaB in mononuclear cells. Metabolism 2004, 53, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Chaudhuri, A.; Ghanim, H.; Mohanty, P. Proinflammatory effects of glucose and anti-inflammatory effect of insulin: Relevance to cardiovascular disease. Am. J. Cardiol. 2007, 99, 15B–26B. [Google Scholar] [CrossRef]
- Aljada, A.; Friedman, J.; Ghanim, H.; Mohanty, P.; Hofmeyer, D.; Chaudhuri, A.; Dandona, P. Glucose ingestion induces an increase in intranuclear nuclear factor kappa B, a fall in cellular inhibitor kappa B, and an increase in tumor necrosis factor alpha messenger RNA by mononuclear cells in healthy human subjects. Metabolism 2006, 55, 1177–1185. [Google Scholar] [CrossRef]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [Green Version]
- Keuper, M.; Blüher, M.; Schön, M.R.; Möller, P.; Dzyakanchuk, A.; Amrein, K.; Debatin, K.M.; Wabitsch, M.; Fischer-Posovszky, P. An inflammatory micro-environment promotes human adipocyte apoptosis. Mol. Cell. Endocrinol. 2011, 339, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, M.; Chu, S.H.; Jia, L.S.; Rose, P.; Sabapathy, K.; Schantz, J.T.; Cheung, N.S.; Spencer, J.P.E.; Armstrong, J.S. The pro-inflammatory oxidant hypochlorous acid induces Bax-dependent mitochondrial permeabilisation and cell death through AIF-/EndoG-dependent pathways. Cell. Signal. 2007, 19, 705–714. [Google Scholar] [CrossRef]
- Carling, D. AMPK signalling in health and disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef]
- Zheng, T.; Yang, X.; Li, W.; Wang, Q.; Chen, L.; Wu, D.; Bian, F.; Xing, S.; Jin, S. Salidroside attenuates high-fat diet-induced nonalcoholic fatty liver disease via AMPK-dependent TXNIP/NLRP3 pathway. Oxid. Med. Cell. Longev. 2018, 22, 8597897. [Google Scholar] [CrossRef]
- Li, W.; Qiu, X.; Jiang, H.; Zhi, Y.; Fu, J.; Liu, J. Ulinastatin inhibits the inflammation of LPS-induced acute lung injury in mice via regulation of AMPK/NF-κB pathway. Int. Immunopharmacol. 2015, 29, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.A.; Ford, B.M.; Block, K.; Kasinath, B.S.; Gorin, Y.; Choudhury, G.G.; Barnes, J.L.; Abboud, H.E. AMPK negatively regulates Nox4-dependent activation of p53 and epithelial cell apoptosis in diabetes. J. Biol. Chem. 2010, 285, 37503–37512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.S.; Qureshi, N.A.; Jabeen, M.; Wajid, F.; Sabri, S.; Shakir, M. The role of garlic oil in the amelioration of oxidative stress and tissue damage in rohu Labeo rohita treated with silver nanoparticles. Fish. Sci. 2020, 86, 255–269. [Google Scholar] [CrossRef]
- Govindasamy, R.; Rahuman, A.A. Histopathological studies and oxidative stress of synthesized silver nanoparticles in Mozambique tilapia (Oreochromis mossambicus). J. Environ. Sci. 2012, 24, 1091–1098. [Google Scholar] [CrossRef]
- Chi, H.P.; Yi, J.; Gu, M.B. Biomarker gene response in male Medaka (Oryzias latipes) chronically exposed to silver nanoparticle. Ecotox. Environ. Saf. 2012, 78, 239–245. [Google Scholar]
- Pantuck, E.J.; Pantuck, C.B.; Kappas, A.; Conney, A.H.; Anderson, K.E. Effects of protein and carbohydrate content of diet on drug conjugation. Clin. Pharm. 1991, 50, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Lu, K.L.; Liu, W.B.; Jiang, G.Z.; Xu, W.N. Effects of dietary lipid and carbohydrate and their interaction on growth performance and body composition of juvenile blunt snout bream, Megalobrama amblycephala. Isr. J. Aquacult. Bamidgeh. 2014, 66, 931. [Google Scholar]
- Clark, N.J.; Boyle, D.; Eynon, B.P.; Handy, R.D. Dietary exposure to silver nitrate compared to two forms of silver nanoparticles in rainbow trout: Bioaccumulation potential with minimal physiological effects. Environ. Sci. Nano 2019, 6, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1990; p. 1298. [Google Scholar]
- Habte-Tsion, H.M.; Ren, M.C.; Liu, B.; Ge, X.P.; Xie, J.; Chen, R. Threonine modulates immune response, antioxidant status and gene expressions of antioxidant enzymes and antioxidant immune-cytokine-related signaling molecules in juvenile blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2016, 51, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yang, Z.; Bo, L.; Zhao, Z.; Zhou, Q.; Sun, C. Cytotoxic effects, inflammatory response and apoptosis induction of cyclophosphamide in the peripheral blood leukocyte of blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2019, 93, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhao, X.; Ren, X.; Zhu, R.; Luo, Z.; Ren, B. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria-mediated apoptosis in zebrafish embryos. Aquat. Toxicol. 2016, 180, 56–70. [Google Scholar] [CrossRef]
- Lamprecht, W.; Trautschold, I. Adenosine-5-triphosphate. Determination with hexokinase and glucose-6-phosphate dehydrogenase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; Volume 4, pp. 2097–2110. [Google Scholar]
- Adam, H. Adenosine-5′-diphosphate and Adenosine-5′-monophosphate. Meth. Enzym. Anal. 1965, 4, 573–577. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2-ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gao, Z.X.; Luo, W.; Liu, H.; Zeng, C.; Liu, X.L.; Yi, S.K.; Wang, W.M. Transcriptome analysis and SSR/SNP markers information of the blunt snout bream (Megalobrama amblycephala). PLoS ONE 2012, 7, e42637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Liang, H.L.; Mokrani, A.; Ji, K.; Yu, H.; Ge, X.P.; Ren, M.C.; Xie, J.; Pan, L.K.; Sun, A. Dietary histidine affects intestinal antioxidant enzyme activities, antioxidant gene expressions and inflammatory factors in juvenile blunt snout bream (Megalobrama amblycephala). Aquacult. Nutr. 2018, 25, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional regulation of hepatic glucose metabolism in fish. Fish Physiol. Biochem. 2009, 35, 519–539. [Google Scholar] [CrossRef]
- Hemre, G.I.; Mommsen, T.P.; Krogdahl, A. Carbohydrates in fish nutrition, effects on growth, glucose metabolism and hepatic enzymes. Aquacult. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Kamalam, B.S.; Medale, F.; Panserat, S. Utilisation of dietary carbohydrates in farmed fishes, new insights on influencing factors, biological limitations and future strategies. Aquaculture 2017, 467, 3–27. [Google Scholar] [CrossRef]
- Shabrangharehdasht, M.; Mirvaghefi, A.; Farahmand, H. Effects of nanosilver on hematologic, histologic and molecular parameters of rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2020, 225, 105549. [Google Scholar] [CrossRef] [PubMed]
- Ramadi, K.B.; Mohamed, Y.A.; Al-Sbiei, A.; Almarzooqi, S.; Bashir, G.; Al Dhanhani, A.; Sarawathiamma, D.; Qadri, S.; Yasin, J.; Nemmar, A.; et al. Acute systemic exposure to silver-based nanoparticles induces hepatotoxicity and NLRP3-dependent inflammation. Nanotoxicology 2016, 10, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Pi, D.G.; Liu, Y.L.; Shi, H.F.; Li, S.; Odle, J.; Lin, X.; Zhu, H.L.; Chen, F.; Hou, Y.Q.; Leng, W.B. Dietary supplementation of aspartate enhances intestinal integrity and energy status in weanling piglets after lipopolysaccharide challenge. J. Nutr. Biochem. 2014, 25, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.Y.; Pan, M.Z.; Liu, J.H.; Yang, M.X.; Gu, Z.X.; Zhang, Y.; Liu, G.X.; Liu, D.; Zhang, W.B.; Mai, K.S. Chronic stress of high dietary carbohydrate level causes inflammation and influences glucose transport through SOCS3 in Japanese flounder Paralichthys olivaceus. Sci. Rep. 2018, 8, 7415. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.H.; Lin, Y.; Pan, W.J.; Huang, X.; Ge, X.P.; Ren, M.C.; Zhou, Q.L.; Liu, B. Identification of differentially expressed microRNAs associate with glucose metabolism in different organs of blunt snout bream (Megalobrama amblycephala). Int. J. Mol. Sci. 2017, 18, 1161. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Liu, W.B.; Shi, H.J.; Mi, H.F.; Li, X.F. Benfotiamine ameliorates high-carbohydrate diet-induced hepatic oxidative stress, inflammation and apoptosis in Megalobrama amblycephala. Aquac. Res. 2021. [Google Scholar] [CrossRef]
- Yildirim, N.C.; Danabas, S. Assessment of immunomodulatory biomarkers (Tnf-α, Il-1β and Il-6) in liver of Capoeta umbla for biomonitoring of pollution in uzuncayir dam lake (Tunceli, Turkey). Iran. J. Fish. Sci. 2014, 13, 653–666. [Google Scholar]
- Wang, B.; Feng, L.; Chen, G.F.; Jiang, W.D.; Liu, Y.; Kuang, S.Y.; Jiang, J.; Tang, L.; Wu, P.; Tang, W.N.; et al. Jian carp (Cyprinus carpio var. jian) intestinal immune responses, antioxidant status and tight junction protein mRNA expression are modulated via nrf2 and pkc in response to dietary arginine deficiency. Fish Shellfish Immun. 2016, 51, 116–124. [Google Scholar] [CrossRef]
- Hussain, S.M.; Hess, K.L.; Gearhart, J.M.; Geiss, K.T.; Schlager, J.J. In vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol. Vitr. 2005, 19, 975–983. [Google Scholar] [CrossRef]
- Coll Ferrer, M.C.; Eckmann, U.N.; Composto, R.J.; Eckmann, D.M. Hemocompatibility and biocompatibility of antibacterial biomimetic hybrid films. Toxicol. Appl. Pharmacol. 2013, 272, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Kong, H. The effect of Lycium barbarum polysaccharide on alcoholinduced oxidative stress in rats. Molecules 2010, 16, 2542–2550. [Google Scholar] [CrossRef]
- Allard, M.F.; Schönekess, B.O.; Henning, S.L.; English, D.R.; Lopaschuk, G.D. Contribution of oxidative metabolism and glycolysis to ATP production in hypertrophied hearts. Am. J. Physiol. 1994, 267, H742–H750. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.B.; Zhang, D.D.; Wang, K.Z.; Xia, S.L.; Li, X.F. Molecular characterization of AMP-activated protein kinase α2 from herbivorous fish Megalobrama amblycephala and responsiveness to glucose loading and dietary carbohydrate levels. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 208, 24–34. [Google Scholar] [CrossRef]
- Khan, A.A.; Alanazi, A.M.; Alsaif, N.; Al-Anazi, M.; Bhat, M.A. Potential cytotoxicity of silver nanoparticles: Stimulation of autophagy and mitochondrial dysfunction in cardiac cells. SaudiJ. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.B.; Zhang, D.D.; Cao, X.F.; Shi, H.J.; Li, X.F. Interactions between dietary carbohydrate and metformin, implications on energy sensing, insulin signaling pathway, glycolipid metabolism and glucose tolerance in blunt snout bream Megalobrama amblycephala. Aquaculture 2018, 483, 183–195. [Google Scholar] [CrossRef]
- Ren, Y.; Shi, Y.; Wang, Y.; Li, Y.; Wu, S.; Li, H.; Zhang, Y.; Duan, H. p38 MAPK pathway is involved in high glucose-induced thioredoxin interacting protein induction in mouse mesangial cells. FEBS Lett. 2010, 584, 3480–3485. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Rong, Y.; Zhang, M.; Wang, X.L.; LeMaire, S.A.; Coselli, J.S.; Zhang, Y.; Shen, Y.H. Up-regulation of thioredoxin interacting protein (Txnip) by p38 MAPK and FOXO1 contributes to the impaired thioredoxin activity and increased ROS in glucose-treated endothelial cells. Biochem. Biophys, Res. Commun. 2009, 381, 660–665. [Google Scholar] [CrossRef] [Green Version]
- Turturro, F.; Friday, E.; Welbourne, T. Hyperglycemia regulates thioredoxin-ROS activity through induction of thioredoxin-interacting protein (TXNIP) in metastatic breast cancer-derived cells MDA-MB-231. BMC Cancer 2007, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, N.; Reddy, M.A.; Guha, M.; Natarajan, R. High glucose-induced expression of proinflammatory cytokine and chemokine genes in monocytic cells. Diabetes 2003, 52, 1256–1264. [Google Scholar] [CrossRef] [Green Version]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar]
- Brown, D.M.; Donaldson, K.; Borm, P.J.; Schins, R.P.; Dehnhardt, M.; Gilmour, P.; Jimenez, L.A.; Stone, V. Calcium and ROS-mediated activation of transcription factors and TNF-alpha cytokine gene expression in macrophages exposed to ultrafine particles. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 286, L344–L353. [Google Scholar] [CrossRef]
- Nishanth, R.P.; Jyotsna, R.G.; Schlager, J.J.; Hussain, S.M.; Reddanna, P. Inflammatory responses of RAW 264.7 macrophages upon exposure to nanoparticles: Role of ROS-NFκB signaling pathway. Nanotoxicology 2011, 5, 502–516. [Google Scholar] [CrossRef]
- Salminen, A.; Hyttinen, J.M.T.; Kaarniranta, K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: Impact on healthspan and lifespan. J. Mol. Med. 2011, 89, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Green, C.J.; Pedersen, M.; Pedersen, B.K.; Scheele, C. Elevated NF-κB activation is conserved in human myocytes cultured from obese type 2 diabetic patients and attenuated by AMP-activated protein kinase. Diabetes 2011, 60, 2810–2819. [Google Scholar] [CrossRef] [Green Version]
- Guragain, D.; Gurung, P.; Chang, J.H.; Katila, N.; Chang, H.W.; Jeong, B.S.; Choi, D.Y.; Kim, J.A. AMPK is essential for IL-10 expression and for maintaining balance between inflammatory and cytoprotective signaling. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2020, 1864, 129631. [Google Scholar] [CrossRef]
- Martin, D.A.; Elkon, K.B. Mechanisms of apoptosis. Rheum. Dis. Clin. N. Am. 2004, 30, 441–454. [Google Scholar] [CrossRef]
- Strasser, A.; O’Connor, L.; Dixit, V.M. Apoptosis signaling. Annu. Rev. Biochem. 2000, 69, 217–245. [Google Scholar] [CrossRef]
- Kim, H.R.; Shin, D.Y.; Park, Y.J.; Park, C.W.; Oh, S.M.; Chung, K.H. Silver nanoparticles induce p53-mediated apoptosis in human bronchial epithelial (BEAS-2B) cells. J. Toxicol. Sci. 2014, 39, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.E.; Valentine, R.J.; Cacicedo, J.M.; Gauthier, M.S.; Ido, Y.; Ruderman, N.B. A novel inverse relationship between metformin-triggered AMPK-SIRT1 signaling and p53 protein abundance in high glucose-exposed HepG2 cells. Am. J. Physiol. Cell Physiol. 2012, 303, C4–C13. [Google Scholar] [CrossRef]
- Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell. 2005, 18, 283–293. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, I.J.; Yun, H.; Kang, I.; Choe, W.; Kim, S.S.; Ha, J. AMP-activated protein kinase antagonizes pro-apoptotic extracellular signal-regulated kinase activation by inducing dual-specificity protein phosphatases in response to glucose deprivation in HCT116 carcinoma. J. Biol. Chem. 2010, 285, 14617–14627. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet | High-Carbohydrate Diet | |
|---|---|---|
| Formulation(%) | ||
| Fish meal | 5.00 | 5.00 |
| Soybean meal | 30.00 | 30.00 |
| Rapeseed meal | 18.40 | 18.40 |
| Cottonseed meal | 15.00 | 15.00 |
| Soybean oil | 3.60 | 3.60 |
| Corn starch | 12.00 | 25.00 |
| Microcrystalline cellulose | 13.00 | 0.00 |
| Calcium biphosphate | 1.80 | 1.80 |
| Premix * | 1.20 | 1.20 |
| Proximate composition(% air-dry basis) | ||
| Moisture | 8.79 | 8.96 |
| Crude lipid | 4.97 | 5.25 |
| Ash | 7.00 | 6.96 |
| Crude protein | 31.71 | 31.94 |
| Crude fiber | 15.76 | 3.00 |
| Nitrogen-free extract † | 31.77 | 43.89 |
| Energy (MJ/kg) | 18.89 | 18.82 |
| Target Gene | Forward (5′-3) | Reverse (5′-3) | Accession Numbers or References |
|---|---|---|---|
| AMPKα 1 | AGTTGGACGAGAAGGAG | AGGGCATACAAAATCAC | ARF07712.1 |
| AMPKα 2 | ACAGCCCTAAGGCACGATG | TGGGTCGGGTAGTGTTGAG | KX061841 |
| TXNIP | CAGACTTGCTGTCCCCTAC | CTCCAGAACCAACTTATCG | MW582526 |
| Trx | TCACAATCGCCTTCAATC | CTCCCTTCTTACCCACAA | [29] |
| CAT | CAGTGCTCCTGATACCCAGC | TTCTGACACAGACGCTCTCG | [29] |
| Cu/Zn-SOD | AGTTGCCATGTGCACTTTTCT | AGGTGCTAGTCGAGTGTTAGG | KF479046.1 |
| Mn-SOD | AGCTGCACCACAGCAAGCAC | TCCTCCACCATTCGGTGACA | KF195932.1 |
| GPx1 | GAACGCCCACCCTCTGTTTG | GAACGCCCACCCTCTGTTTG | KF378713.1 |
| NF-κB | GAAGAAGGATGTGGGAGATG | TGTTGTCGTAGATGGGCTGAG | [30] |
| TNFα | TGGAGAGTGAACCAGGACCA | AGAGACCTGGCTGTAGACGA | KU976426.1 |
| IL 1β | ACGATAAGACCAGCACGACC | CTGTTTCCGTCTCTCAGCGT | [30] |
| IL 6 | CAGCAGAATGGGGGAGTTATC | CTCGCAGAGTCTTGACATCCTT | KJ755058.1 |
| IL 8 | CAGAGAGTCGACGCATTGGT | ATTCACGGTGCTTTGTTGGC | [31] |
| IL 10 | GTGTTTTCGGGTGCAAGTGG | ATGAACGAGATCCTGCGCTT | [31] |
| P 53 | CAGCAGGAGCCAATCCATCA | ACGTACTCCCCAGACCTGAA | [31] |
| Bcl 2 | CCAACTCATCAGGAAACAA | GGGTGCTGCGGGTAAC | EU490408.1 |
| Bax | ATCCAGCCAGCATCGT | CACTATCCCCAAGACCC | AF231015.1 |
| Caspase 3 | TCGTTCGTCTGTGTCCTGTTGAG | GCTGTGGAGAAGGCGTAGAGG | KY006115.1 |
| Caspase 9 | AATAAAGCACCGAGCG | GGGAGGAGGCCGATGAGCACTATCT | KM604705.1 |
| EF1α | CTTCTCAGGCTGACTGTGC | CCGCTAGCATTACCCTCC | X77689.1 |
| Parameters | Groups | p-Value | |||
|---|---|---|---|---|---|
| C | CS | HC | HCS | ||
| Initial weight (g) | 20.12 ± 0.07 | 20.42 ± 0.23 | 20.41 ± 0.21 | 20.22 ± 0.12 | >0.05 |
| Final weight (g) | 124.57 ± 5.18 a | 65.69 ± 8.41 d | 101.24 ± 4.11 b | 88.84 ± 4.31 c | <0.05 |
| WGR † (%) | 462.98 ± 10.20 a | 225.12 ± 9.57 d | 394.19 ± 8.31 b | 336.12 ± 5.61 c | <0.05 |
| SGR§(% day−1) | 3.12 ± 0.02 a | 1.95 ± 0.01 d | 2.81 ± 0.04 b | 2.64 ± 0.01 c | <0.05 |
| RFI || (% body weight d−1) | 3.35 ± 0.01 | 2.94 ± 0.03 | 3.12 ± 0.01 | 2.81 ± 0.03 | >0.05 |
| FCR | 1.35 ± 0.02 | 1.58 ± 0.01 | 1.30 ± 0.02 | 1.27 ± 0.02 | >0.05 |
| Parameters | Groups | p-Value | |||
|---|---|---|---|---|---|
| C | CS | HC | HCS | ||
| Plasma parameters | |||||
| AST (U/L) | 23.47 ± 0.12 c | 43.11 ± 0.11 a | 33.10 ± 0.21 b | 35.47 ± 0.35 b | <0.05 |
| ALT (U/L) | 2.01 ± 0.01 d | 6.24 ± 0.03 a | 3.45 ± 0.01 c | 4.85 ± 0.14 b | <0.05 |
| IL 1β (ng/L) | 1.21 ± 0.01 d | 7.11 ± 0.02 a | 2.31 ± 0.01 c | 5.34 ± 0.03 b | <0.05 |
| IL 6 (ng/L) | 21.24 ± 0.31 c | 45.72 ± 0.65 a | 33.54 ± 1.04 b | 34.78 ± 0.52 b | <0.05 |
| Liver parameters | |||||
| ROS (% control) | 1.02 ± 0.02 c | 7.14 ± 0.01 a | 4.25 ± 0.01 b | 4.64 ± 0.03 b | <0.05 |
| T-AOC (U/mg protein) | 1.45 ± 0.01 a | 0.35 ± 0.01 c | 0.54 ± 0.02 b | 0.55 ± 0.01 b | <0.05 |
| SOD (U/mg protein) | 12.36 ± 0.04 a | 6.35 ± 0.04 c | 10.19 ± 0.11 b | 9.22 ± 0.05 b | <0.05 |
| CAT (U/mg protein) | 20.21 ± 0.22 a | 4.49 ± 0.03 c | 11.34 ± 0.21 b | 10.18 ± 0.02 b | <0.05 |
| MDA (nmol/mg protein) | 2.34 ± 0.02 c | 10.29 ± 0.12 a | 8.21 ± 0.01 b | 8.61 ± 0.05 b | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Zhao, C.-Y.; Guan, J.-F.; Liu, X.-C.; Li, X.-F.; Xie, D.-Z.; Xu, C. High-Carbohydrate Diet Alleviates the Oxidative Stress, Inflammation and Apoptosis of Megalobrama amblycephala Following Dietary Exposure to Silver Nanoparticles. Antioxidants 2021, 10, 1343. https://doi.org/10.3390/antiox10091343
Chen F, Zhao C-Y, Guan J-F, Liu X-C, Li X-F, Xie D-Z, Xu C. High-Carbohydrate Diet Alleviates the Oxidative Stress, Inflammation and Apoptosis of Megalobrama amblycephala Following Dietary Exposure to Silver Nanoparticles. Antioxidants. 2021; 10(9):1343. https://doi.org/10.3390/antiox10091343
Chicago/Turabian StyleChen, Fang, Cai-Yuan Zhao, Jun-Feng Guan, Xiao-Cheng Liu, Xiang-Fei Li, Di-Zhi Xie, and Chao Xu. 2021. "High-Carbohydrate Diet Alleviates the Oxidative Stress, Inflammation and Apoptosis of Megalobrama amblycephala Following Dietary Exposure to Silver Nanoparticles" Antioxidants 10, no. 9: 1343. https://doi.org/10.3390/antiox10091343
APA StyleChen, F., Zhao, C.-Y., Guan, J.-F., Liu, X.-C., Li, X.-F., Xie, D.-Z., & Xu, C. (2021). High-Carbohydrate Diet Alleviates the Oxidative Stress, Inflammation and Apoptosis of Megalobrama amblycephala Following Dietary Exposure to Silver Nanoparticles. Antioxidants, 10(9), 1343. https://doi.org/10.3390/antiox10091343