
Perinatal Resveratrol Therapy to Dioxin-Exposed Dams Prevents the Programming of Hypertension in Adult Rat Offspring




Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.3. Gut Microbiota Compositions
2.4. Measurement of Cytokines in the Kidneys
2.5. Analysis of NO Pathway
2.6. Statistical Analysis
3. Results
3.1. Morphological Values and Blood Pressures
3.2. AHR Signaling Pathway
3.3. Cytokine Concentrations in the Kidneys
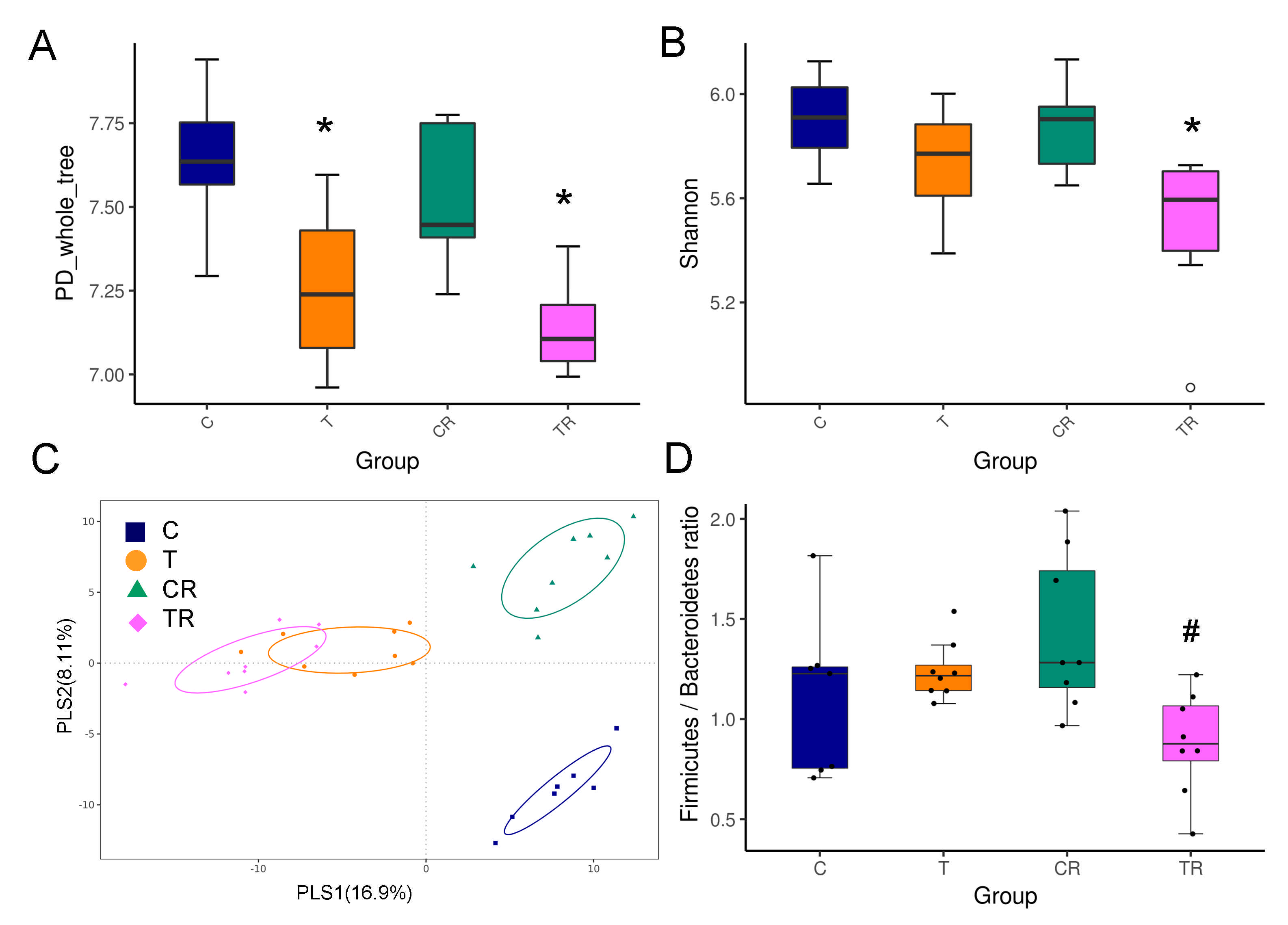
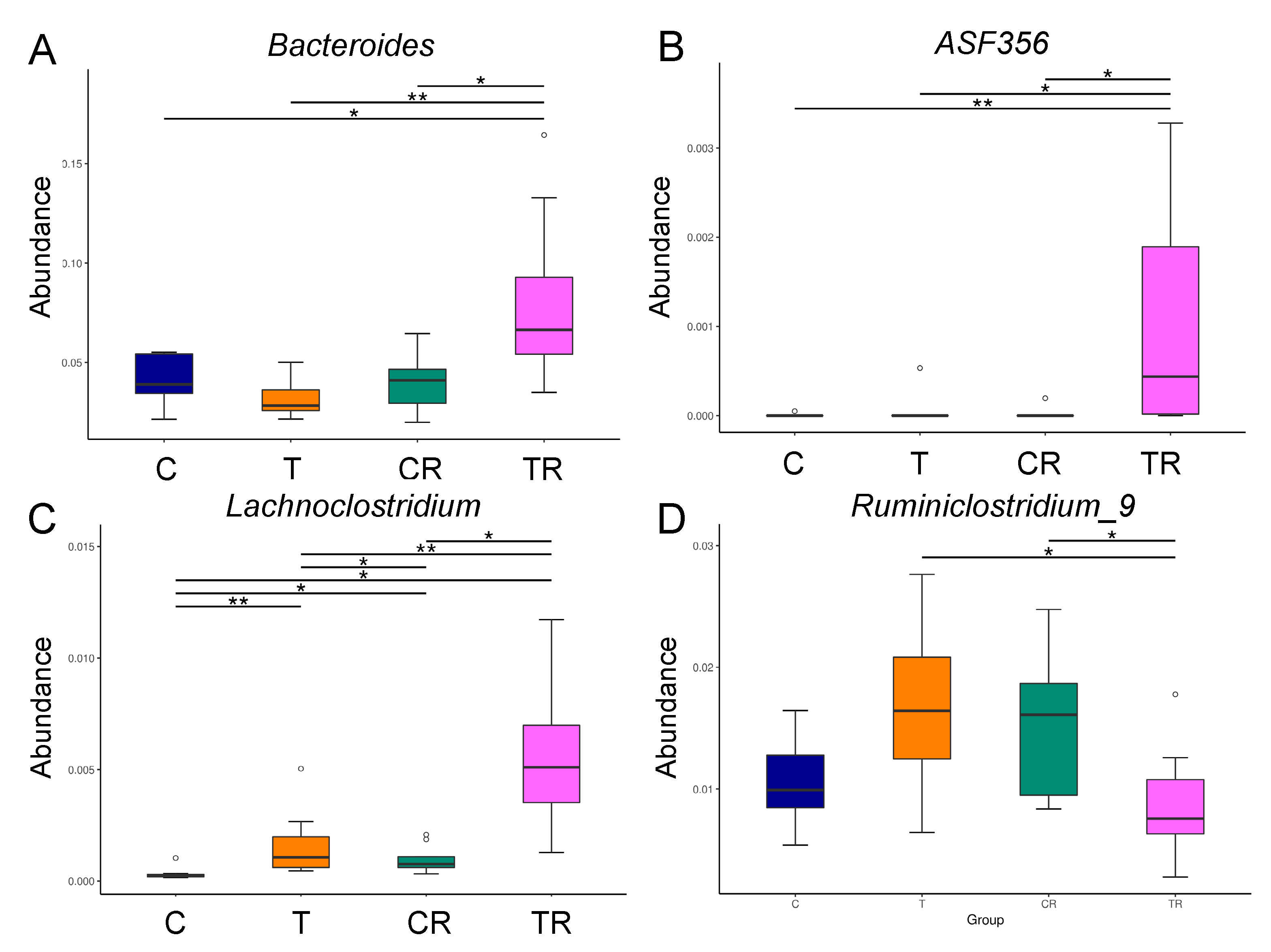
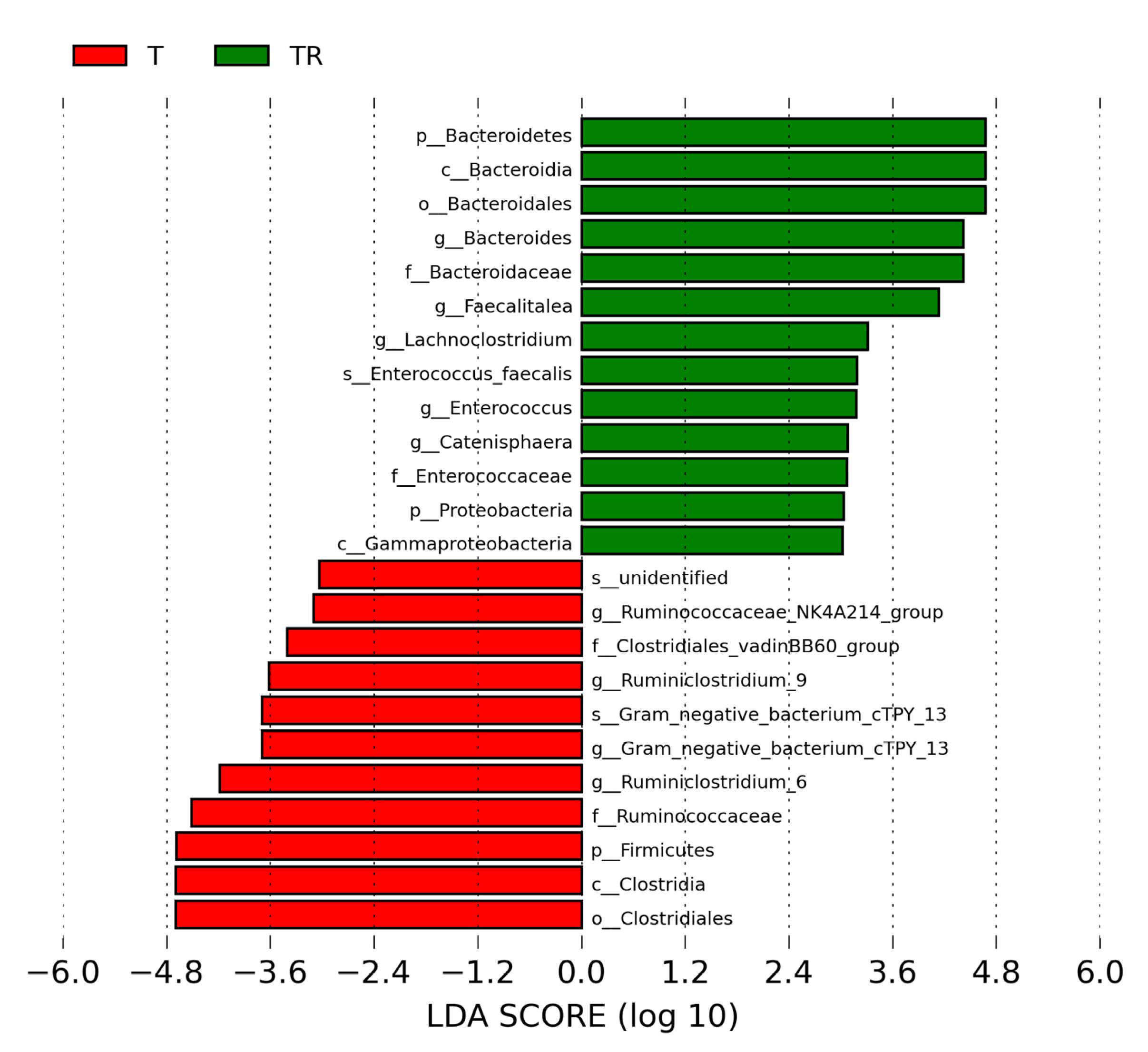
3.4. Gut Microbiota Composition
3.5. Plasma NO-Related Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Hypertension. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/hypertension (accessed on 28 July 2021).
- Godfrey, K.M.; Barker, D.J. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M. The birth and future health of DOHaD. J. Dev. Orig. Health Dis. 2015, 6, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Joles, J.A. Reprogramming: A preventive strategy in hypertension focusing on the kidney. Int. J. Mol. Sci. 2015, 17, E23. [Google Scholar] [CrossRef] [Green Version]
- Nüsken, E.; Dötsch, J.; Weber, L.T.; Nüsken, K. Developmental Programming of Renal Function and Re-Programming Approaches. Front. Pediatr. 2018, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Paixão, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 144. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Bertram, J.F.; Brenner, B.M.; Fall, C.; Hoy, W.E.; Ozanne, S.E.; Vikse, B.E. Effect of fetal and child health on kidney development and long-term risk of hypertension and kidney disease. Lancet 2013, 382, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Animal Models for DOHaD Research: Focus on Hypertension of Developmental Origins. Biomedicines 2021, 9, 623. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, M.J.; Bolger, P.M.; Jose, P.A. The developing kidney and environmental toxins. Pediatrics 2004, 113, 1084–1091. [Google Scholar]
- Dopico, M.; Gómez, A. Review of the current state and main sources of dioxins around the world. J. Air Waste Manag. Assoc. 2015, 65, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- NIOSH Pocket Guide to Chemical Hazards. 2020. Available online: https://www.cdc.gov/niosh/npg/ (accessed on 25 August 2021).
- Ahmed, R.G. Perinatal TCDD exposure alters developmental neuroendocrine system. Food Chem. Toxicol. 2011, 49, 1276–1284. [Google Scholar] [CrossRef]
- Aragon, A.C.; Goens, M.B.; Carbett, E.; Walker, M.K. Perinatal 2,3,7,8-tetrachlorodibenzo-p-dioxin exposure sensitizes offspring to angiotensin II-induced hypertension. Cardiovasc. Toxicol. 2008, 8, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Chan, J.Y.H.; Yu, H.R.; Lee, W.C.; Wu, K.L.H.; Chang-Chien, G.P.; Lin, S.; Hou, C.Y.; Tain, Y.L. Targeting on Gut Microbiota-Derived Metabolite Trimethylamine to Protect Adult Male Rat Offspring against Hypertension Programmed by Combined Maternal High-Fructose Intake and Dioxin Exposure. Int. J. Mol. Sci. 2020, 21, 5488. [Google Scholar] [CrossRef]
- Amakura, Y.; Tsutsumi, T.; Sasaki, K.; Nakamura, M.; Yoshida, T.; Maitani, T. Influence of food polyphenols on aryl hydrocarbon receptor-signaling pathway estimated by in vitro bioassay. Phytochemistry 2008, 69, 3117–3130. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial action of resveratrol: How and why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Daiber, A.; Förstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Tain, Y.L. Preventive Aspects of Early Resveratrol Supplementation in Cardiovascular and Kidney Disease of Developmental Origins. Int. J. Mol. Sci. 2021, 22, 4210. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Yang, H.W.; Tain, Y.L. Perinatal Resveratrol Therapy Prevents Hypertension Programmed by Maternal Chronic Kidney Disease in Adult Male Offspring: Implications of the Gut Microbiome and Their Metabolites. Biomedicines 2020, 8, 567. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Reyes, L.M.; Morton, J.S.; Fung, D.; Schneider, J.; Davidge, S.T. Effect of resveratrol on metabolic and cardiovascular function in male and female adult offspring exposed to prenatal hypoxia and a high-fat diet. J. Physiol. 2016, 594, 1465–1482. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Regulation of Nitric Oxide Production in the Developmental Programming of Hypertension and Kidney Disease. Int. J. Mol. Sci. 2019, 20, 681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W. Aryl hydrocarbon receptor (AHR) functions: Balancing opposing processes including inflammatory reactions. Biochem. Pharmacol. 2020, 178, 114093. [Google Scholar] [CrossRef] [PubMed]
- Alrafas, H.R.; Busbee, P.B.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol modulates the gut microbiota to prevent murine colitis development through induction of Tregs and suppression of Th17 cells. J. Leukoc. Biol. 2019, 106, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension 2001, 7, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Franczak, A.; Nynca, A.; Valdez, K.E.; Mizinga, K.M.; Petroff, B.K. Effects of acute and chronic exposure to the aryl hydrocarbon receptor agonist 2,3,7,8-tetrachlorodibenzo-p-dioxin on the transition to reproductive senescence in female Sprague-Dawley rats. Biol. Reprod. 2006, 74, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Maternal resveratrol therapy protected adult rat offspring against hypertension programmed by combined exposures to asymmetric dimethylarginine and trimethylamine-N-oxide. J. Nutr. Biochem. 2021, 93, 108630. [Google Scholar] [CrossRef]
- Wagner, B.D.; Grunwald, G.K.; Zerbe, G.O.; Mikulich-Gilbertson, S.K.; Robertson, C.E.; Zemanick, E.T.; Harris, J.K. On the use of diversity measures in longitudinal sequencing studies of microbial communities. Front. Microbiol. 2018, 9, 1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N. The role of endogenous aryl hydrocarbon receptor signaling in cardiovascular physiology. J. Cardiovasc. Dis. Res. 2011, 2, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Lin, Y.J.; Tain, Y.L. Maternal Exposure to Bisphenol A Combined with High-Fat Diet-Induced Programmed Hypertension in Adult Male Rat Offspring: Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 4382. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Lin, Y.J.; Lu, P.C.; Tain, Y.L. Maternal Resveratrol Therapy Protects Male Rat Offspring against Programmed Hypertension Induced by TCDD and Dexamethasone Exposures: Is It Relevant to Aryl Hydrocarbon Receptor? Int. J. Mol. Sci. 2018, 19, 2459. [Google Scholar] [CrossRef] [Green Version]
- Elijovich, F.; Laffer, C.L.; Sahinoz, M.; Pitzer, A.; Ferguson, J.F.; Kirabo, A. The Gut Microbiome, Inflammation, and Salt-Sensitive Hypertension. Curr. Hypertens. Rep. 2020, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Hase, K. Commensal microbiota regulates T cell fate decision in the gut. Semin. Immunopathol. 2015, 37, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zuo, K.; Li, J.; Xu, Q.; Hu, C.; Gao, Y.; Chen, M.; Hu, R.; Liu, Y.; Chi, H.; Yin, Q.; et al. Dysbiotic gut microbes may contribute to hypertension by limiting vitamin D production. Clin. Cardiol. 2019, 42, 710–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.M.; Yang, M.X.; Wu, Q.F.; Chen, J.; Deng, S.F.; Chen, L.; Wei, D.N.; Liang, F.R. Improvement of intestinal flora: Accompany with the antihypertensive effect of electroacupuncture on stage 1 hypertension. Chin. Med. 2021, 16, 7. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward | Reverse |
|---|---|---|---|
| AHR | AM902286.1 | GTCCTCAGCAGGAACGAAAG | CCAGGGAAGTCCAACTGTGT |
| AHRR | AY367561.1 | CAGCAACATGGCTTCTTTCA | TGAAGCACTGCATTCCAGAC |
| ARNT | NM_012780.3 | GTCTCCCTCCCAGATGATGA | GCTGGTAGCCAACAGTAGCC |
| CYP1A1 | NM_012540.3 | GCACTCTGGACAAACACCTG | ATATCCACCTTCTCGCCTGG |
| R18S | M11188.1 | GCCGCGGTAATTCCAGCTCCA | CCCGCCCGCTCCCAAGATC |
| Groups | C | T | CR | TR |
|---|---|---|---|---|
| N = 7 | N = 8 | N = 8 | N = 8 | |
| Body weight (BW) (g) | 415 ± 20 | 332 ± 7 * | 416 ± 25 | 355 ± 15 * |
| Left kidney weight (g) | 1.86 ± 0.1 | 1.73 ± 0.05 | 1.69 ± 0.1 | 1.78 ± 0.07 |
| Left kidney weight/100 g BW | 0.45 ± 0.02 | 0.52 ± 0.01 * | 0.41 ± 0.01 | 0.50 ± 0.03 |
| Systolic blood pressure (mmHg) | 129 ±1 | 142 ± 2 * | 130 ± 0 | 133 ± 0 # |
| Diastolic blood pressure (mmHg) | 80 ± 2 | 87 ± 3 * | 86 ± 2 | 85 ± 1 |
| Mean arterial pressure (mmHg) | 97 ± 1 | 105 ± 2 * | 100 ± 1 | 101 ± 1 |
| Groups | C | T | CR | TR |
|---|---|---|---|---|
| N = 7 | N = 8 | N = 8 | N = 8 | |
| IL-2 (pg/g protein) | 4283 ± 653 | 9075 ± 1270 * | 6391 ± 1037 | 5620 ± 317 |
| IL-6 (pg/g protein) | 6864 ± 1198 | 14,948 ± 1805 * | 10,724 ± 1858 | 5373 ± 557 # |
| IL-10 (pg/g protein) | 2040 ± 1072 | 10,390 ± 5561 | 562 ± 279 | 3136 ± 961 |
| IL-17A (pg/g protein) | 5529 ± 870 | 11,332 ± 1411 * | 8151 ± 1152 | 7293 ± 303 # |
| IL-17F (pg/g protein) | 2293 ± 336 | 3262 ± 388 | 2394 ± 179 | 3095 ± 187 |
| IL-22 (pg/g protein) | 1979 ± 273 | 3208 ± 404 * | 2332 ± 218 | 2838 ± 141 |
| IFN-γ (pg/g protein) | 3725 ± 586 | 8545 ± 1128 * | 5668 ± 982 | 4829 ± 337 # |
| Groups | C | T | CR | TR |
|---|---|---|---|---|
| N = 7 | N = 8 | N = 8 | N = 8 | |
| l-Citrulline (μM) | 71.8 ± 3.5 | 65 ± 3.8 | 73.9 ± 2.3 | 63.5 ± 5.3 |
| l-arginine (μM) | 134.9 ± 5.7 | 149.6 ± 7.9 | 189.3 ± 9.3 * | 159.9 ± 5.7 |
| ADMA (μM) | 1.56 ± 0.06 | 1.6 ± 0.08 | 1.37 ± 0.07 | 1.59 ± 0.08 |
| SDMA (μM) | 1.35 ± 0.03 | 1.69 ± 0.08 | 1.28 ± 0.09 * | 1.32 ± 0.03 |
| l-arginine-to-ADMA ratio (μM/μM) | 87.5 ± 6 | 96.3 ± 8.4 | 141.2 ± 11.1 * | 102.4 ± 6.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-N.; Hung, C.-H.; Hou, C.-Y.; Chang, C.-I.; Tain, Y.-L. Perinatal Resveratrol Therapy to Dioxin-Exposed Dams Prevents the Programming of Hypertension in Adult Rat Offspring. Antioxidants 2021, 10, 1393. https://doi.org/10.3390/antiox10091393
Hsu C-N, Hung C-H, Hou C-Y, Chang C-I, Tain Y-L. Perinatal Resveratrol Therapy to Dioxin-Exposed Dams Prevents the Programming of Hypertension in Adult Rat Offspring. Antioxidants. 2021; 10(9):1393. https://doi.org/10.3390/antiox10091393
Chicago/Turabian StyleHsu, Chien-Ning, Chih-Hsing Hung, Chih-Yao Hou, Chi-I. Chang, and You-Lin Tain. 2021. "Perinatal Resveratrol Therapy to Dioxin-Exposed Dams Prevents the Programming of Hypertension in Adult Rat Offspring" Antioxidants 10, no. 9: 1393. https://doi.org/10.3390/antiox10091393
APA StyleHsu, C. -N., Hung, C. -H., Hou, C. -Y., Chang, C. -I., & Tain, Y. -L. (2021). Perinatal Resveratrol Therapy to Dioxin-Exposed Dams Prevents the Programming of Hypertension in Adult Rat Offspring. Antioxidants, 10(9), 1393. https://doi.org/10.3390/antiox10091393