
Treatment with Bixin-Loaded Polymeric Nanoparticles Prevents Cigarette Smoke-Induced Acute Lung Inflammation and Oxidative Stress in Mice

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nanoparticle Preparation
2.3. Bixin Content
2.4. Physical Characterization of Nanoparticles
2.5. Nanoparticle Thermal Stability
2.6. Animals
2.7. Experimental Procedure
2.8. Bronchoalveolar Lavage
2.9. Tissue Processing and Lung Homogenate Preparation
2.10. Myeloperoxidase (MPO) Activity
2.11. Malondialdehyde (MDA) Products and Nitrite Quantification
2.12. Superoxide Dismutase (SOD) and Catalase (CAT) Activity
2.13. Western Blotting
2.14. Enzyme-Linked Immunosorbent Assay (ELISA) to Detect TNF-α Levels
2.15. Gelatin Zymography
2.16. Immunohistochemistry
2.17. Statistical Analysis
3. Results
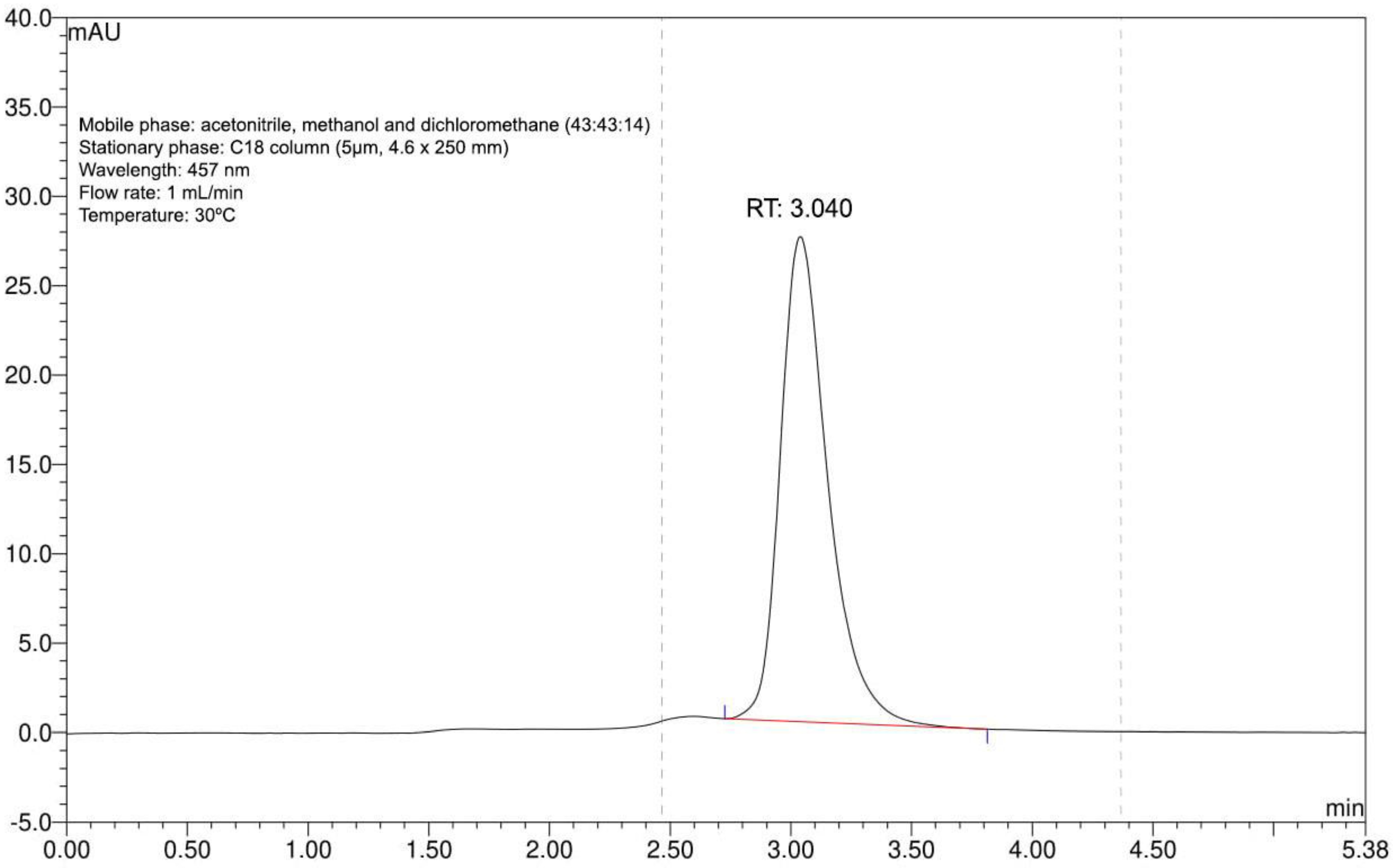
3.1. Bixin Extraction
3.2. Nanoparticle Characterization
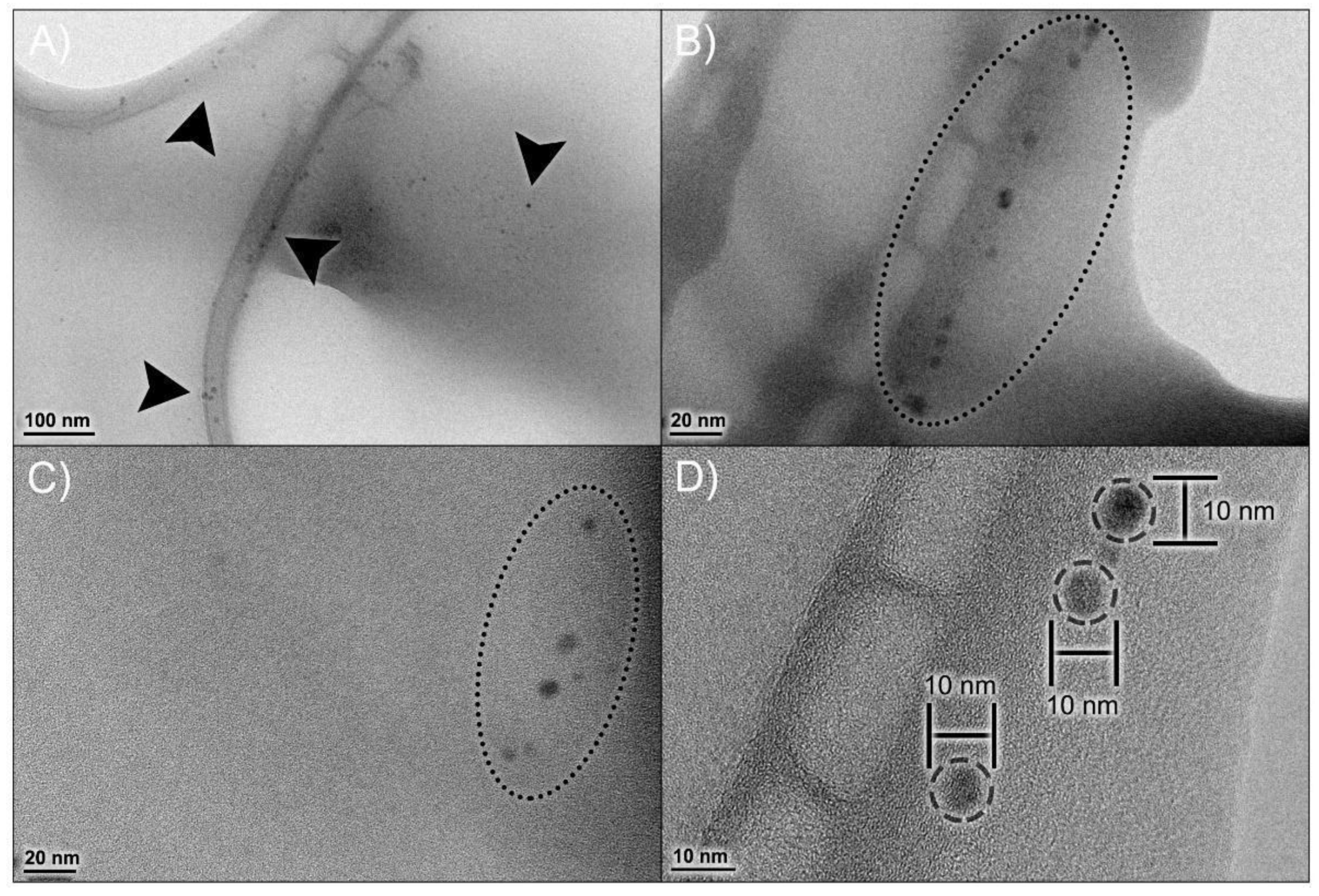
3.2.1. Size, Zeta Potential and External Morphology
3.2.2. Thermal Stability of Nanoparticles
3.3. npBX Prevents ALI Induced by CS Exposure
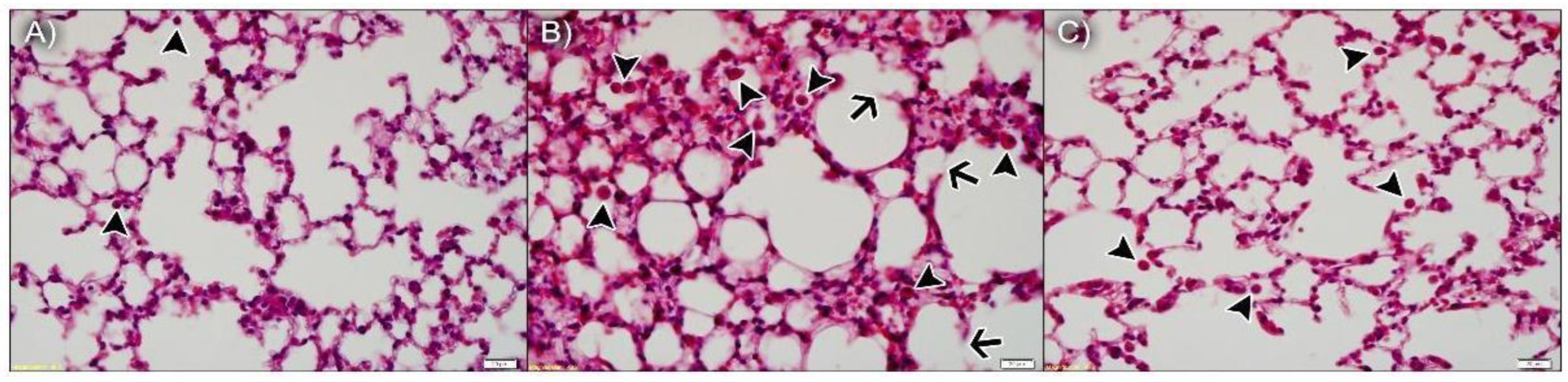
3.3.1. npBX Preserves Pulmonary Histoarchitecture
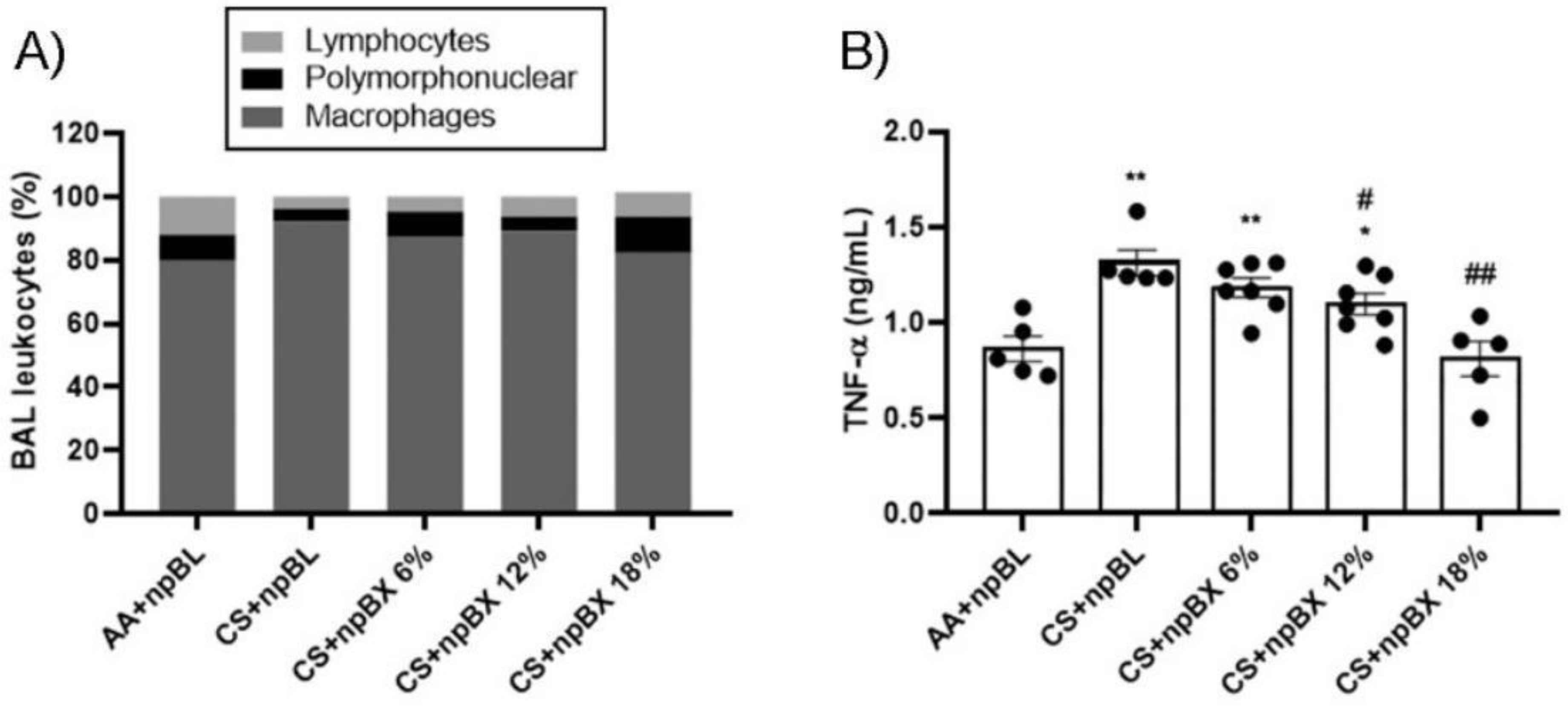
3.3.2. npBX Prevents the Increase in Leukocyte Numbers and TNF-α Levels in BALF from Mice Exposed to CS
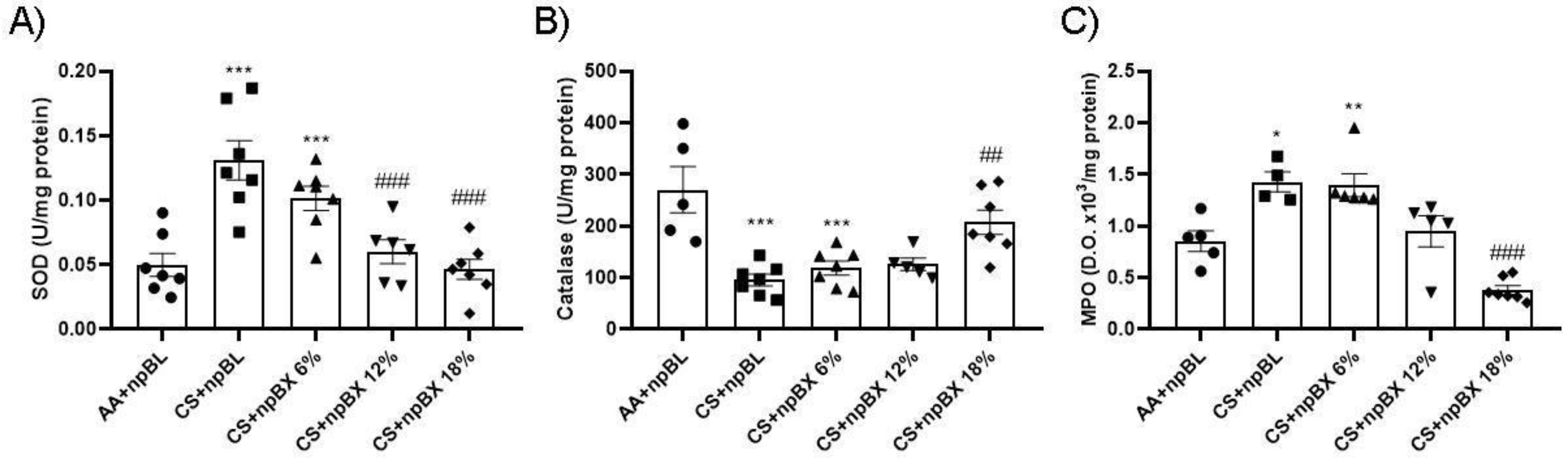
3.3.3. npBX Prevents Redox Imbalance Induced by CS Exposure in Mouse Lungs
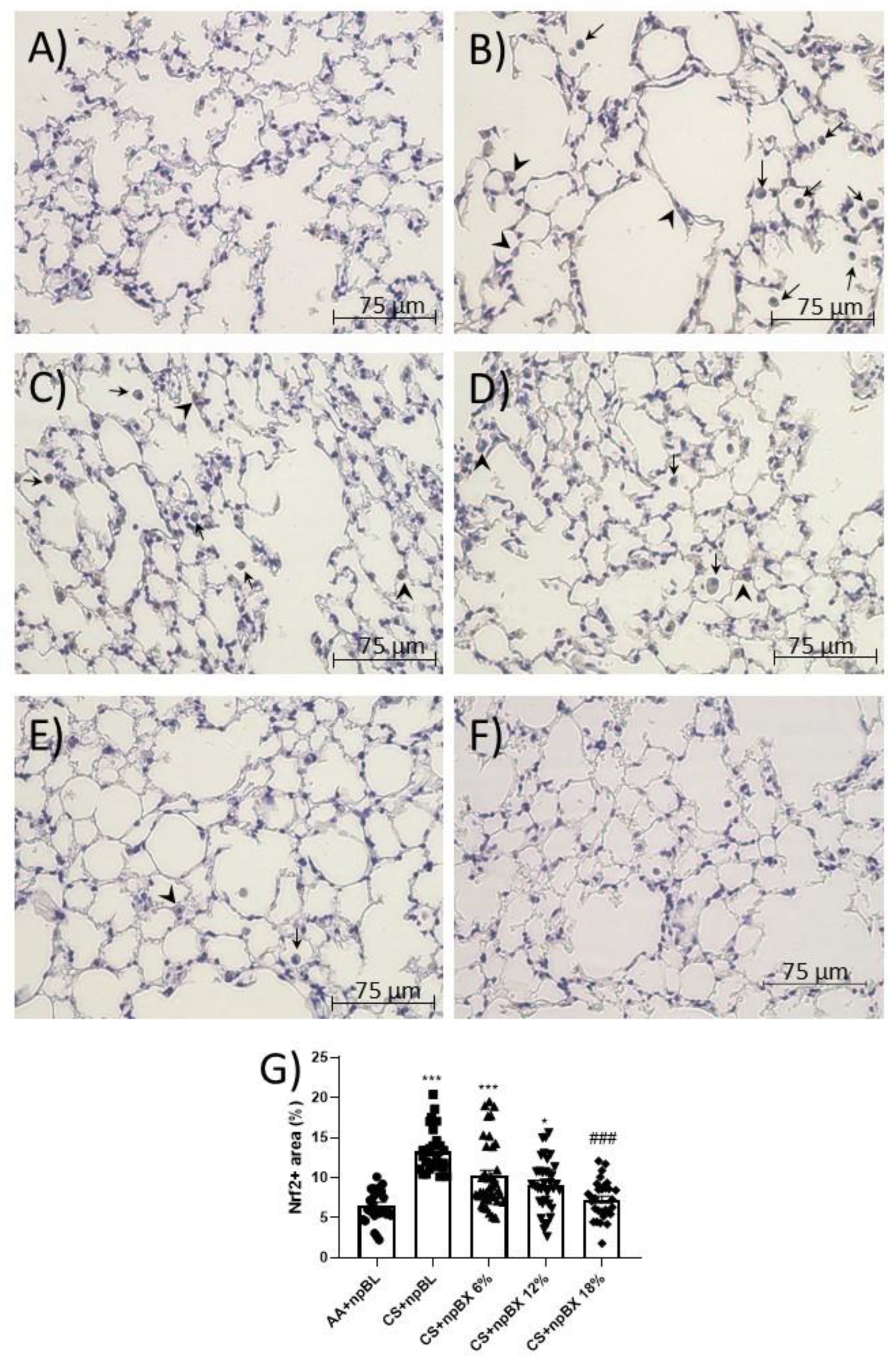
3.3.4. npBX Antioxidant Activity is Independent of Nrf2
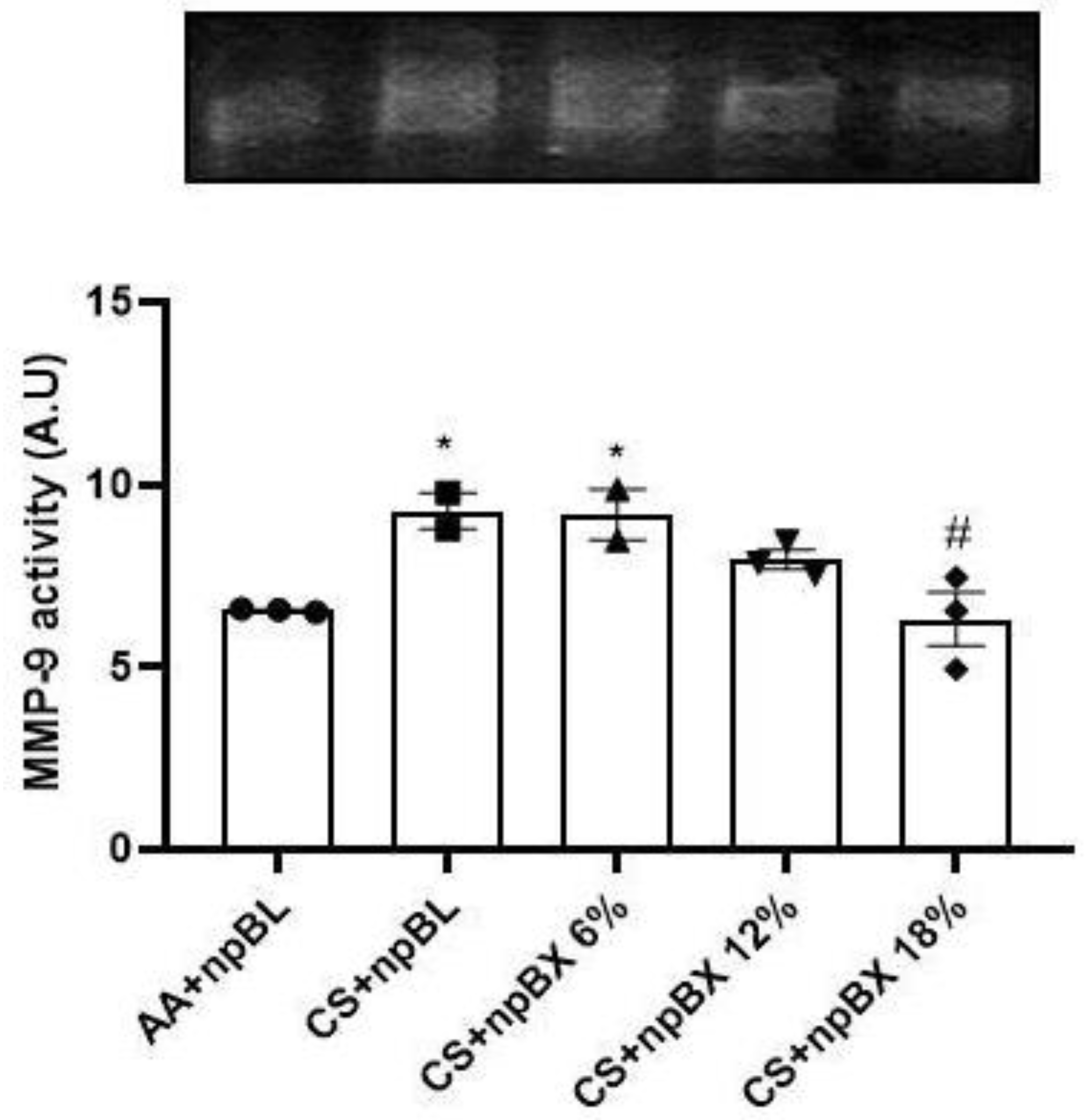
3.3.5. Bixin Nanoparticles Protected Lungs from MMP-9 Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brody, J.S.; Spira, A. State of the art. Chronic obstructive pulmonary disease, inflammation, and lung cancer. Proc. Am. Thorac. Soc. 2006, 3, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Musk, A.W.; de Klerk, N.H. History of tobacco and health. Respirology 2003, 8, 286–290. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 4 April 2020).
- Sapey, E.; Stockley, R.A. COPD exacerbations 2: Aetiology. Thorax 2006, 61, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.L.; Cosio, M.; Churg, A. Animal models of chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L1–L15. [Google Scholar] [CrossRef]
- Brode, S.K.; Ling, S.C.; Chapman, K.R. Alpha-1 antitrypsin deficiency: A commonly overlooked cause of lung disease. CMAJ 2012, 184, 1365–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurzem, J.R.; Rennard, S.I. Pathogenesis of COPD. Semin. Respir. Crit. Care Med. 2005, 26, 142–153. [Google Scholar] [CrossRef]
- Barnes, P.J. Oxidative stress-based therapeutics in COPD. Redox Biol. 2020, 33, 101544. [Google Scholar] [CrossRef]
- Barnes, P.J. New anti-inflammatory targets for chronic obstructive pulmonary disease. Nat. Rev. Drug Discov. 2013, 12, 543–559. [Google Scholar] [CrossRef]
- Kennedy-Feitosa, E.; Okuro, R.T.; Pinho Ribeiro, V.; Lanzetti, M.; Barroso, M.V.; Zin, W.A.; Porto, L.C.; Brito-Gitirana, L.; Valenca, S.S. Eucalyptol attenuates cigarette smoke-induced acute lung inflammation and oxidative stress in the mouse. Pulm. Pharmacol. Ther. 2016, 41, 11–18. [Google Scholar] [CrossRef]
- Lopes, A.A.; Ferreira, T.S.; Nesi, R.T.; Lanzetti, M.; Pires, K.M.; Silva, A.M.; Borges, R.M.; Silva, A.J.; Valenca, S.S.; Porto, L.C. Antioxidant action of propolis on mouse lungs exposed to short-term cigarette smoke. Bioorg. Med. Chem. 2013, 21, 7570–7577. [Google Scholar] [CrossRef] [Green Version]
- Moura, R.S.; Ferreira, T.S.; Lopes, A.A.; Pires, K.M.; Nesi, R.T.; Resende, A.C.; Souza, P.J.; Silva, A.J.; Borges, R.M.; Porto, L.C.; et al. Effects of Euterpe oleracea Mart. (ACAI) extract in acute lung inflammation induced by cigarette smoke in the mouse. Phytomedicine 2012, 19, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; Adcock, I.M. Oxidative stress and redox regulation of lung inflammation in COPD. Eur. Respir. J. 2006, 28, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Silva Bezerra, F.; Valença, S.S.; Lanzetti, M.; Pimenta, W.A.; Castro, P.; Gonçalves Koatz, V.L.; Porto, L.C. α-Tocopherol and ascorbic acid supplementation reduced acute lung inflammatory response by cigarette smoke in mouse. Nutrition 2006, 22, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Valenca, S.S.; Silva Bezerra, F.; Lopes, A.A.; Romana-Souza, B.; Marinho Cavalcante, M.C.; Lima, A.B.; Gonçalves Koatz, V.L.; Porto, L.C. Oxidative stress in mouse plasma and lungs induced by cigarette smoke and lipopolysaccharide. Environ. Res. 2008, 108, 199–204. [Google Scholar] [CrossRef]
- Ferreira, T.S.; Lanzetti, M.; Barroso, M.V.; Rueff-Barroso, C.R.; Benjamim, C.F.; de Brito-Gitirana, L.; Porto, L.C.; Valenca, S.S. Oxidative stress and inflammation are differentially affected by atorvastatin, pravastatin, rosuvastatin, and simvastatin on lungs from mice exposed to cigarette smoke. Inflammation 2014, 37, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- MacNee, W. Oxidants/Antioxidants and COPD. Chest 2000, 117, 303S–317S. [Google Scholar] [CrossRef]
- Pryor, W.A.; Stone, K. Oxidants in cigarette smoke. Radicals, hydrogen peroxide, peroxynitrate, and peroxynitrite. Ann. N. Y. Acad. Sci. 1993, 686, 12–27. [Google Scholar] [CrossRef]
- Helen, A.; Vijayammal, P.L. Effect of vitamin A supplementation on cigarette smoke-induced lipid peroxidation. Vet. Hum. Toxicol. 1997, 39, 18–21. [Google Scholar]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Et Biophys. Acta Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef]
- Lanzetti, M.; Bezerra, F.S.; Romana-Souza, B.; Brando-Lima, A.C.; Koatz, V.L.G.; Porto, L.C.; Valenca, S.S. Mate tea reduced acute lung inflammation in mice exposed to cigarette smoke. Nutrition 2008, 24, 375–381. [Google Scholar] [CrossRef]
- Taniguchi, A.; Tsuge, M.; Miyahara, N.; Tsukahara, H. Reactive Oxygen Species and Antioxidative Defense in Chronic Obstructive Pulmonary Disease. Antioxidants 2021, 10, 1537. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.M.; Soares, V.L.; de Oliveira, T.M.; Gesteira, A.S.; Otoni, W.C.; Costa, M.G. Isolation and purification of RNA from tissues rich in polyphenols, polysaccharides, and pigments of annatto (Bixa orellana L.). Mol. Biotechnol. 2007, 37, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Dequigiovanni, G.; Ramos, S.L.F.; Alves-Pereira, A.; Fabri, E.G.; Picanço-Rodrigues, D.; Clement, C.R.; Gepts, P.; Veasey, E.A. Highly structured genetic diversity of Bixa orellana var. urucurana, the wild ancestor of annatto, in Brazilian Amazonia. PLoS ONE 2018, 13, e0198593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, L.; Somacal, S.; Nichelle, S.M.; Rampelotto, C.; Robalo, S.S.; Roehrs, M.; Emanuelli, T. Short-Term Bixin Supplementation of Healthy Subjects Decreases the Susceptibility of LDL to Cu(2+)-Induced Oxidation Ex Vivo. J. Nutr. Metab. 2019, 2019, 9407069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, Y.; Phaniendra, A.; Periyasamy, L. Bixin Triggers Apoptosis of Human Hep3B Hepatocellular Carcinoma Cells: An Insight to Molecular and IN SILICO Approach. Nutr. Cancer 2018, 70, 971–983. [Google Scholar] [CrossRef]
- Pinzon-Garcia, A.D.; Orellano, L.A.A.; de Lazari, M.G.T.; Campos, P.P.; Cortes, M.E.; Sinisterra, R.D. Evidence of hypoglycemic, lipid-lowering and hepatoprotective effects of the Bixin and Bixin: Beta-CD inclusion compound in high-fat-fed obese mice. Biomed. Pharmacother. 2018, 106, 363–372. [Google Scholar] [CrossRef]
- Rojo de la Vega, M.; Krajisnik, A.; Zhang, D.D.; Wondrak, G.T. Targeting NRF2 for Improved Skin Barrier Function and Photoprotection: Focus on the Achiote-Derived Apocarotenoid Bixin. Nutrients 2017, 9, 1371. [Google Scholar] [CrossRef] [Green Version]
- Rojo de la Vega, M.; Zhang, D.D.; Wondrak, G.T. Topical Bixin Confers NRF2-Dependent Protection Against Photodamage and Hair Graying in Mouse Skin. Front. Pharmacol. 2018, 9, 287. [Google Scholar] [CrossRef] [Green Version]
- Somacal, S.; Figueiredo, C.G.; Quatrin, A.; Ruviaro, A.R.; Conte, L.; Augusti, P.R.; Roehrs, M.; Denardin, I.T.; Kasten, J.; da Veiga, M.L.; et al. The antiatherogenic effect of bixin in hypercholesterolemic rabbits is associated to the improvement of lipid profile and to its antioxidant and anti-inflammatory effects. Mol. Cell Biochem. 2015, 403, 243–253. [Google Scholar] [CrossRef]
- Tao, S.; Park, S.L.; Rojo de la Vega, M.; Zhang, D.D.; Wondrak, G.T. Systemic administration of the apocarotenoid bixin protects skin against solar UV-induced damage through activation of NRF2. Free Radic. Biol. Med. 2015, 89, 690–700. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; Rojo de la Vega, M.; Quijada, H.; Wondrak, G.T.; Wang, T.; Garcia, J.G.; Zhang, D.D. Bixin protects mice against ventilation-induced lung injury in an NRF2-dependent manner. Sci. Rep. 2016, 6, 18760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thresiamma, K.C.; George, J.; Kuttan, R. Protective effect of curcumin, ellagic acid and bixin on radiation induced toxicity. Indian J. Exp. Biol. 1996, 34, 845–847. [Google Scholar] [PubMed]
- Xu, Z.; Kong, X.Q. Bixin ameliorates high fat diet-induced cardiac injury in mice through inflammation and oxidative stress suppression. Biomed. Pharmacother. 2017, 89, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Zhang, H.; Zhang, J.; Li, B.; Zhang, Z.; Tao, S. Bixin protects against particle-induced long-term lung injury in an NRF2-dependent manner. Toxicol. Res. 2018, 7, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Chiste, R.C.; Mercadante, A.Z.; Gomes, A.; Fernandes, E.; Lima, J.L.; Bragagnolo, N. In vitro scavenging capacity of annatto seed extracts against reactive oxygen and nitrogen species. Food Chem. 2011, 127, 419–426. [Google Scholar] [CrossRef]
- Figueiredo-Junior, A.T.; dos Anjos, F.d.F.; Brito, F.d.C.d.M.; Viana, V.G.F.; Valença, S.S.; Lanzetti, M.; Finotelli, P.V. Bixin loaded on polymeric nanoparticles: Synthesis, characterization, and antioxidant applications in a biological system. Appl. Nanosci. 2021, 11, 63–78. [Google Scholar] [CrossRef]
- Fessi, H.; Puisieux, F.; Devissaguet, J.P.; Ammoury, N.; Benita, S. Nanocapsule formation by interfacial polymer deposition following solvent displacement. Int. J. Pharm. 1989, 55, R1–R4. [Google Scholar] [CrossRef]
- Silveira, N.; Longuinho, M.M.; Leitao, S.G.; Silva, R.S.; Lourenco, M.C.; Silva, P.E.; Pinto Mdo, C.; Abracado, L.G.; Finotelli, P.V. Synthesis and characterization of the antitubercular phenazine lapazine and development of PLGA and PCL nanoparticles for its entrapment. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 58, 458–466. [Google Scholar] [CrossRef]
- Raddatz-Mota, D. Chemical Characterization and Quantification of the Pigment Extraction Yield of Seven Mexican Accessions of Bixa Orellana Caracterizacion Quimica Y Cuantificacion del Rendimiento de Extraccion de Pigmento en Siete Accesiones Mexicanas de Bixa orellana. Rev. Mex. Ing. Química 2016, 15, 727–740. [Google Scholar] [CrossRef]
- Suzuki, K.; Ota, H.; Sasagawa, S.; Sakatani, T.; Fujikura, T. Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal. Biochem. 1983, 132, 345–352. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid Peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 421–431. [Google Scholar]
- Bannister, J.V.; Calabrese, L. Assays for Superoxide Dismutase. Methods Biochem. Anal. 1987, 32, 279–312. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Coker, M.S.; Forbes, L.V.; Plowman-Holmes, M.; Murdoch, D.R.; Winterbourn, C.C.; Kettle, A.J. Interactions of staphyloxanthin and enterobactin with myeloperoxidase and reactive chlorine species. Arch. Biochem. Biophys. 2018, 646, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Seagrave, J. Oxidative mechanisms in tobacco smoke-induced emphysema. J. Toxicol. Environ. Health Part A 2000, 61, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, L.; Li, Y.; Huang, X.; Gu, D.; Wei, B.; Su, D.; Jin, G. Ultrasmall superparamagnetic nanoparticles targeting E-selectin: Synthesis and effects in mice in vitro and in vivo. Int. J. Nanomed. 2019, 14, 4517–4528. [Google Scholar] [CrossRef] [Green Version]
- Panyam, J.; Labhasetwar, V. Biodegradable nanoparticles for drug and gene delivery to cells and tissue. Adv. Drug Deliv. Rev. 2003, 55, 329–347. [Google Scholar] [CrossRef]
- Cole, A.J.; Yang, V.C.; David, A.E. Cancer theranostics: The rise of targeted magnetic nanoparticles. Trends Biotechnol. 2011, 29, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Ali Khan, A.; Mudassir, J.; Mohtar, N.; Darwis, Y. Advanced drug delivery to the lymphatic system: Lipid-based nanoformulations. Int. J. Nanomed. 2013, 8, 2733–2744. [Google Scholar] [CrossRef] [Green Version]
- Trewyn, B.G.; Nieweg, J.A.; Zhao, Y.; Lin, V.S.Y. Biocompatible mesoporous silica nanoparticles with different morphologies for animal cell membrane penetration. Chem. Eng. J. 2008, 137, 23–29. [Google Scholar] [CrossRef]
- Barnes, P.J. Immunology of asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2008, 8, 183–192. [Google Scholar] [CrossRef]
- Churg, A.; Wang, R.D.; Tai, H.; Wang, X.; Xie, C.; Dai, J.; Shapiro, S.D.; Wright, J.L. Macrophage metalloelastase mediates acute cigarette smoke-induced inflammation via tumor necrosis factor-alpha release. Am. J. Respir. Crit. Care Med. 2003, 167, 1083–1089. [Google Scholar] [CrossRef]
- Pires, K.M.; Valenca, S.S.; Resende, A.C.; Porto, L.C.; Queiroz, E.F.; Moreira, D.D.; de Moura, R.S. Grape skin extract reduced pulmonary oxidative response in mice exposed to cigarette smoke. Med. Sci. Monit. 2011, 17, BR187–BR195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, T.; Glass, K.; Qiu, W.; Polverino, F.; Gupta, K.; Morrow, J.; Mancini, J.; Vuong, L.; Perrella, M.; Hersh, C.; et al. Haploinsufficiency of Hedgehog interacting protein causes increased emphysema induced by cigarette smoke through network rewiring. Genome Med. 2015, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Park, H.A.; Kwon, O.K.; Jang, Y.G.; Kim, J.Y.; Choi, B.K.; Lee, H.J.; Lee, S.; Paik, J.H.; Oh, S.R.; et al. Asiatic acid inhibits pulmonary inflammation induced by cigarette smoke. Int. Immunopharmacol. 2016, 39, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, A.G.; Dugour, A.V.; Sotomayor, V.; Clauzure, M.; Figueroa, J.M.; Santa-Coloma, T.A. N-acetyl cysteine reverts the proinflammatory state induced by cigarette smoke extract in lung Calu-3 cells. Redox Biol. 2018, 16, 294–302. [Google Scholar] [CrossRef]
- Szabó, C.; Ischiropoulos, H.; Radi, R. Peroxynitrite: Biochemistry, pathophysiology and development of therapeutics. Nat. Rev. Drug Discov. 2007, 6, 662–680. [Google Scholar] [CrossRef]
- Atkinson, J.J.; Senior, R.M. Matrix metalloproteinase-9 in lung remodeling. Am. J. Respir. Cell Mol. Biol. 2003, 28, 12–24. [Google Scholar] [CrossRef]
- Manoury, B.; Nenan, S.; Leclerc, O.; Guenon, I.; Boichot, E.; Planquois, J.-M.; Bertrand, C.P.; Lagente, V. The absence of reactive oxygen species production protects mice against bleomycin-induced pulmonary fibrosis. Respir. Res. 2005, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Churg, A.; Wang, R.D.; Tai, H.; Wang, X.; Xie, C.; Wright, J.L. Tumor necrosis factor-alpha drives 70% of cigarette smoke-induced emphysema in the mouse. Am. J. Respir. Crit. Care Med. 2004, 170, 492–498. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DLS | TEM | |||
|---|---|---|---|---|
| Medium Hydrodynamic Diameter (nm) | Polydisperssivity Index | Zeta Potential (mV) | Mean Diameter (nm) | |
| npBX | 23.57 ± 0.17 | 0.23 ± 0.01 | −31.73 ± 10.10 | 6.51 ± 0.34 |
| Group | Cell × 105 Mean ± SEM (%) | |||
|---|---|---|---|---|
| Total | Macrophages | Neutrophils | Lymphocyte | |
| AA + npBL | 3.08 ± 0.47 (100) | 2.51 ± 0.35 (79.78) | 0.23 ± 0.06 (8.11) | 0.34 ± 0.06 (12.11) |
| CS + npBL | 6.24 ± 0.88 (100) *** | 5.77 ± 0.80 (92.15) *** | 0.26 ± 0.05 (4.14) | 0.21 ± 0.03 (3.71) * |
| CS + npBX 6% | 4.94 ± 0.9 (100) * | 4.36 ± 0.76 (87.5) *** | 0.37 ± 0.09 (7.83) | 0.21 ± 0.05 (4.67) * |
| CS + npBX 12% | 3.84 ± 0.53 (100) ## | 3.44 ± 0.45 (89.19) ### $$ | 0.17 ± 0.04 (4.56) $ | 0.23 ± 0.04 (6.27) |
| CS + npBX 18% | 3.26 ± 0.7 (100) ### $ | 2.64 ± 0.48 (82.19) ### $$$ | 0.36 ± 0.15 (11.66) | 0.26 ± 0.07 (7.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo-Junior, A.T.; Valença, S.S.; Finotelli, P.V.; Anjos, F.d.F.d.; de Brito-Gitirana, L.; Takiya, C.M.; Lanzetti, M. Treatment with Bixin-Loaded Polymeric Nanoparticles Prevents Cigarette Smoke-Induced Acute Lung Inflammation and Oxidative Stress in Mice. Antioxidants 2022, 11, 1293. https://doi.org/10.3390/antiox11071293
Figueiredo-Junior AT, Valença SS, Finotelli PV, Anjos FdFd, de Brito-Gitirana L, Takiya CM, Lanzetti M. Treatment with Bixin-Loaded Polymeric Nanoparticles Prevents Cigarette Smoke-Induced Acute Lung Inflammation and Oxidative Stress in Mice. Antioxidants. 2022; 11(7):1293. https://doi.org/10.3390/antiox11071293
Chicago/Turabian StyleFigueiredo-Junior, Alexsandro Tavares, Samuel Santos Valença, Priscilla Vanessa Finotelli, Francisca de Fátima dos Anjos, Lycia de Brito-Gitirana, Christina Maeda Takiya, and Manuella Lanzetti. 2022. "Treatment with Bixin-Loaded Polymeric Nanoparticles Prevents Cigarette Smoke-Induced Acute Lung Inflammation and Oxidative Stress in Mice" Antioxidants 11, no. 7: 1293. https://doi.org/10.3390/antiox11071293