
R- Is Superior to S-Form of α-Lipoic Acid in Anti-Inflammatory and Antioxidant Effects in Laying Hens




Abstract
1. Introduction
2. Materials and Methods
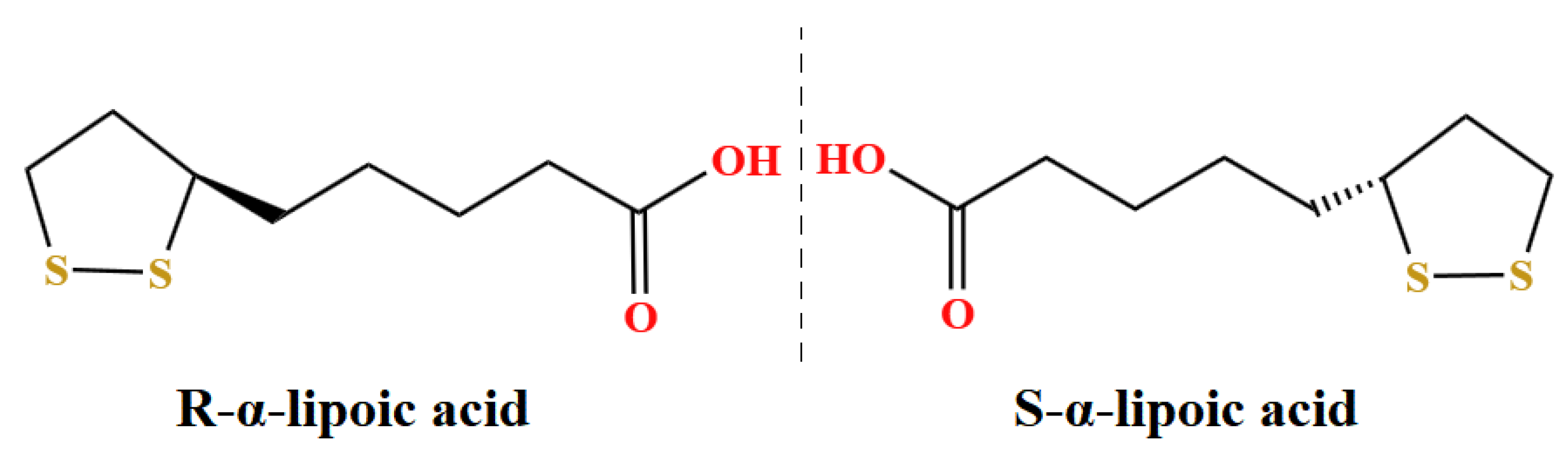
2.1. Materials
2.2. Experimental Design
2.3. Sample Collection
2.4. Indicator Test
2.5. Hematoxylin-Eosin Staining and Histopathology Analyses
2.6. Statistical Analysis
3. Results
3.1. Effect of S-LA and R-LA on Reproductive Performance of Laying Hens under Oxidative Stress
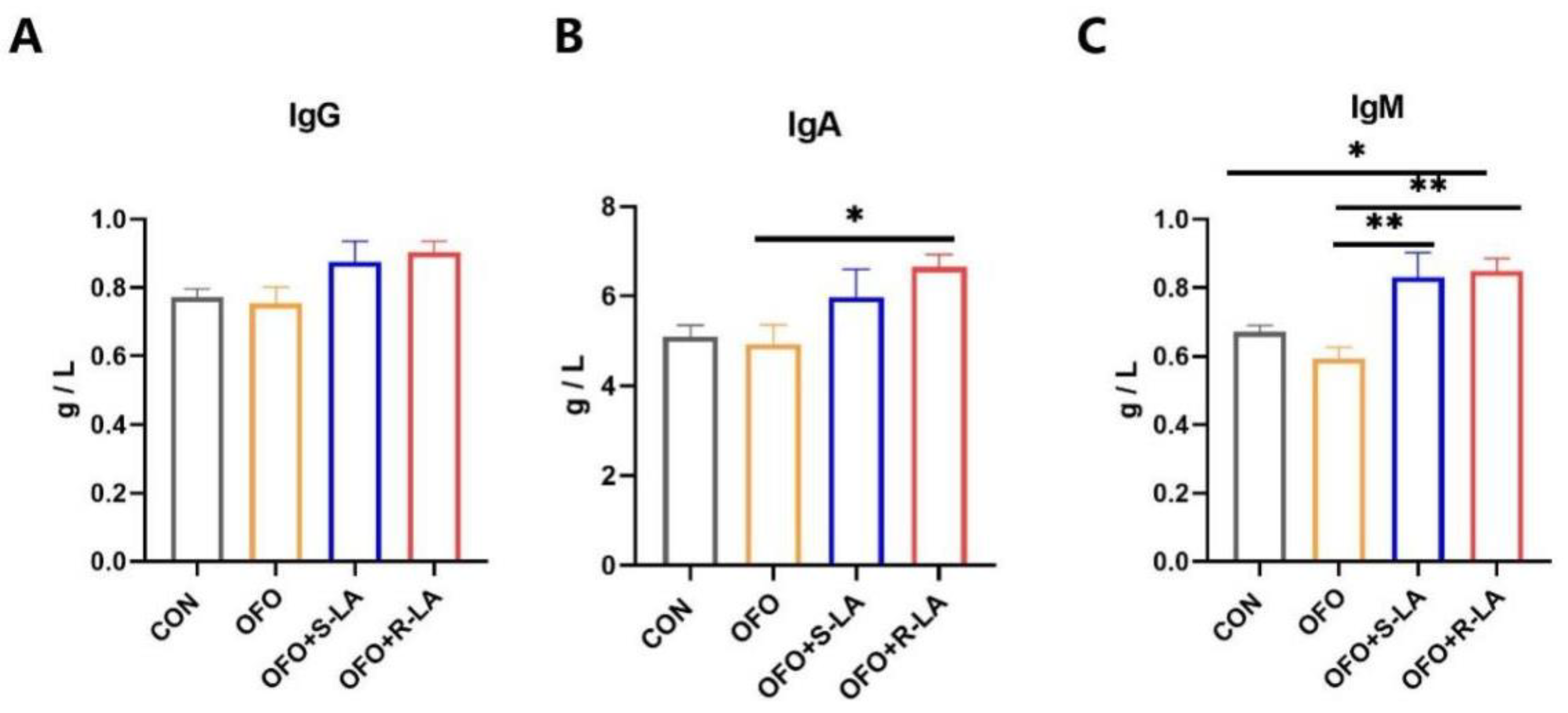
3.2. Effect of S-LA and R-LA on Immunoglobulin of Laying Hens under Oxidative Stress
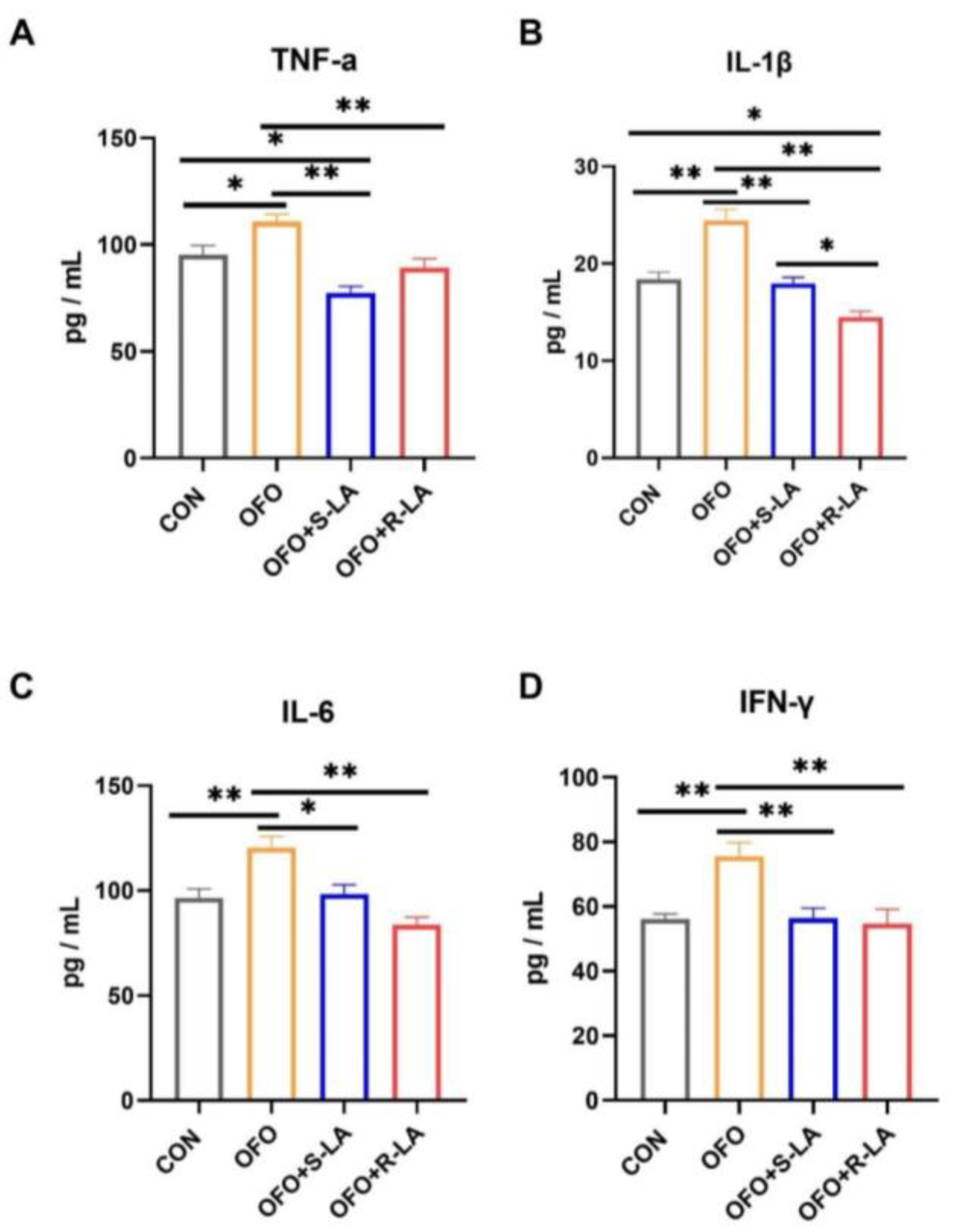
3.3. Effect of S-LA and R-LA on Serum Inflammatory Factors of Laying Hens under Oxidative Stress
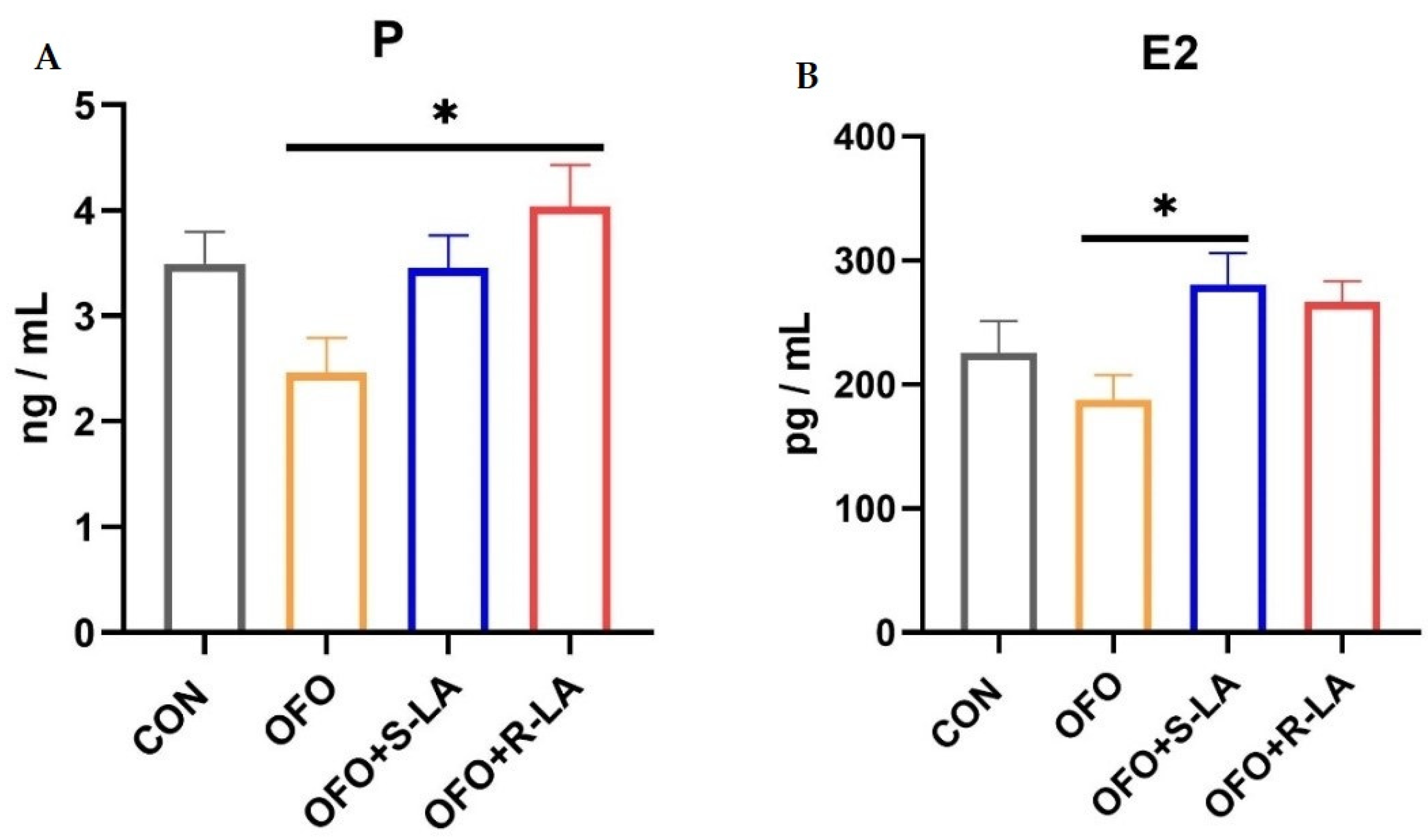
3.4. Effect of S-LA and R-LA on Serum Estrogen of Laying Hens under Oxidative Stress
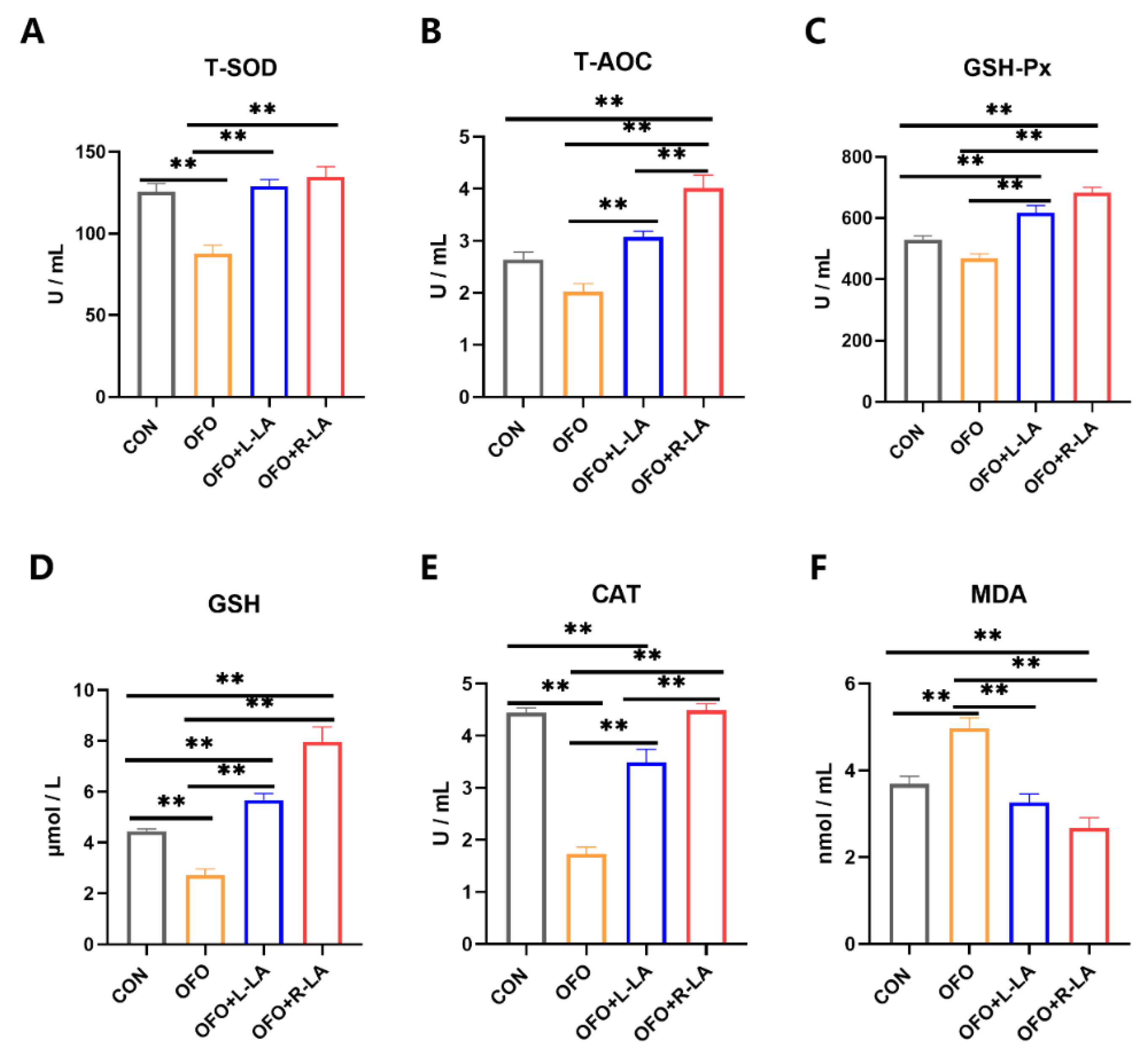
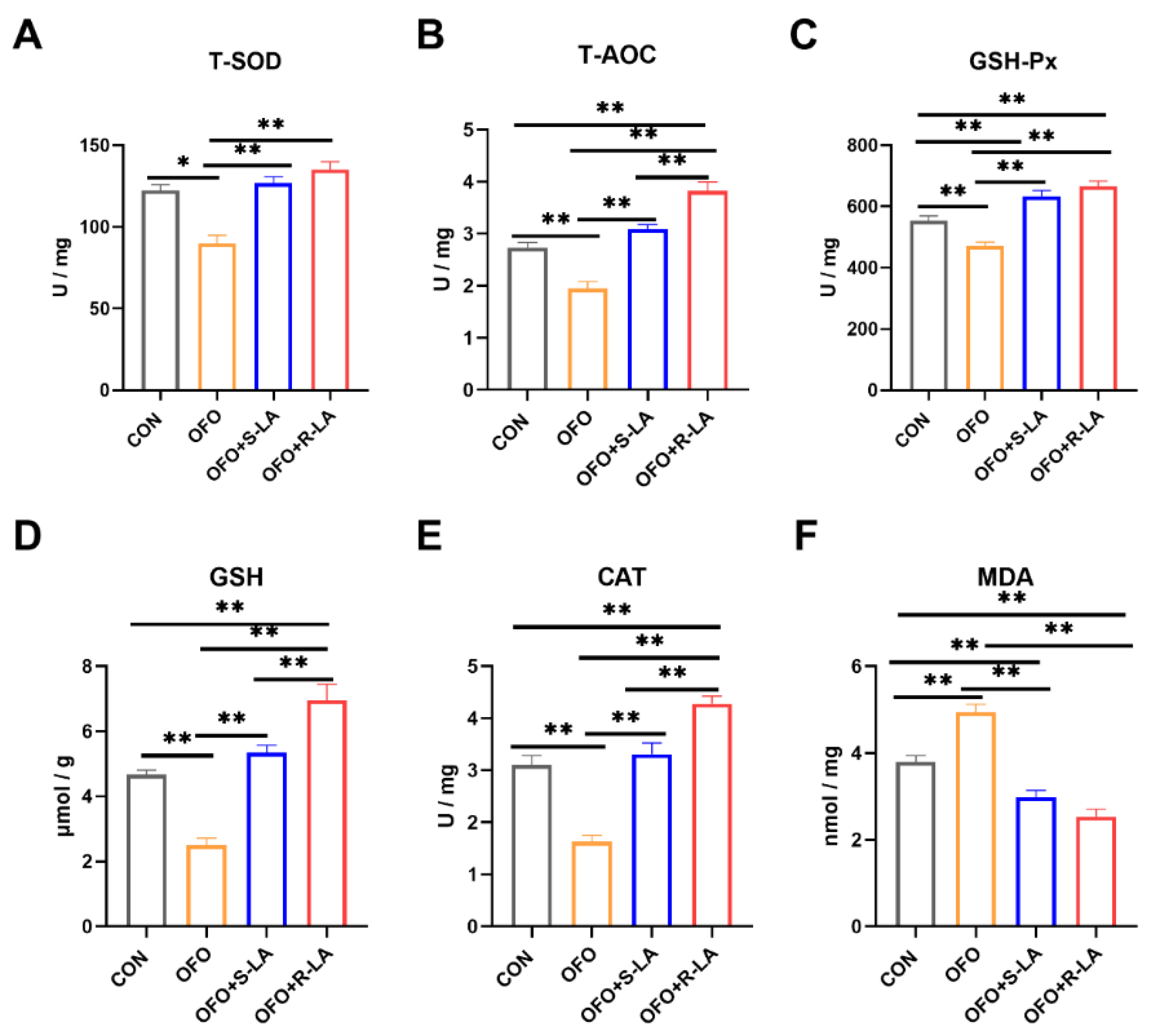
3.5. Effect of S-LA and R-LA on Serum Antioxidant of Laying Hens under Oxidative Stress
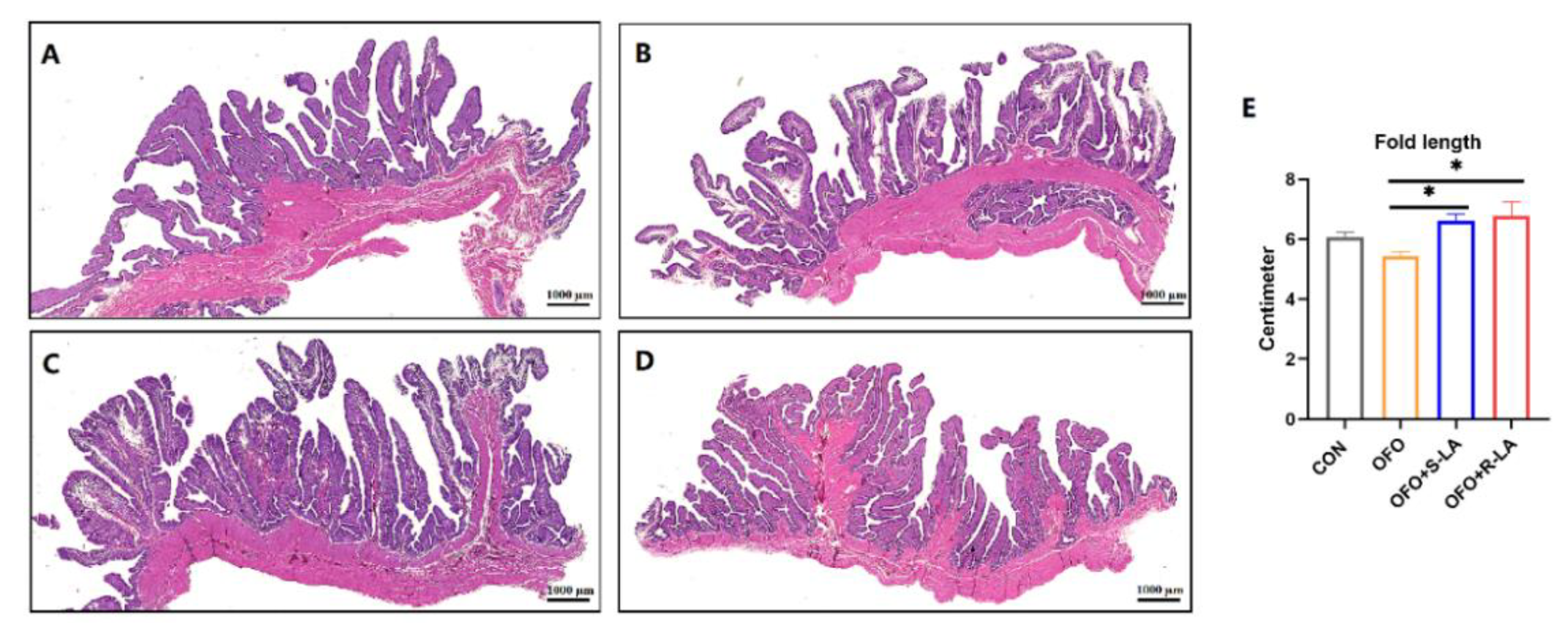
3.6. Effect of S-LA and R-LA on Oviduct Structure of Laying Hens under Oxidative Stress
3.7. Effect of S-LA and R-LA on Oviduct Antioxidant of Laying Hens under Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, M.; Sun, L.; Yang, R.; Wang, Z.; Qi, K. The optimization of fermentation conditions for producing cellulase of Bacillus amyloliquefaciens and its application to goose feed. R. Soc. Open Sci. 2017, 4, 171012. [Google Scholar] [CrossRef]
- Amevor, F.K.; Cui, Z.; Du, X.; Ning, Z.; Shu, G.; Jin, N.; Deng, X.; Tian, Y.; Zhang, Z.; Kang, X.; et al. Combination of quercetin and vitamin E supplementation promotes yolk precursor synthesis and follicle development in aging breeder hens via liver-blood-ovary signal axis. Animals 2021, 11, 1915. [Google Scholar] [CrossRef]
- Dai, B.; Zhang, Y.S.; Ma, Z.L.; Zheng, L.H.; Li, S.J.; Dou, X.H.; Gong, J.S.; Miao, J.F. Influence of dietary taurine and housing density on oviduct function in laying hens. J. Zhejiang Univ. Sci. B 2015, 16, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Bakst, M.R. Physiology and endocrinology symposium: Role of the oviduct in maintaining sustained fertility in hens. J. Anim. Sci. 2011, 89, 1323–1329. [Google Scholar] [CrossRef]
- Myhrstad, M.C.; Ottestad, I.; Günther, C.C.; Ryeng, E.; Holden, M.; Nilsson, A.; Brønner, K.W.; Kohler, A.; Borge, G.I.; Holven, K.B.; et al. The PBMC transcriptome profile after intake of oxidized versus high-quality fish oil: An explorative study in healthy subjects. Genes Nutr. 2016, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y. Molecular and cellular responses to oxidative stress and changes in oxidation-reduction imbalance in the intestine. Am. J. Clin. Nutr. 1999, 70, 557–565. [Google Scholar] [CrossRef]
- Vázquez-Añón, M.; Jenkins, T. Effects of feeding oxidized fat with or without dietary antioxidants on nutrient digestibility, microbial nitrogen, and fatty acid metabolism. J. Dairy Sci. 2007, 90, 4361–4367. [Google Scholar] [CrossRef]
- Feng, R.; Ma, L.J.; Wang, M.; Liu, C.; Yang, R.; Su, H.; Yang, Y.; Wan, J.B. Oxidation of fish oil exacerbates alcoholic liver disease by enhancing intestinal dysbiosis in mice. Commun. Biol. 2020, 3, 481. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Chen, D.; Tian, G.; Zheng, P.; Yu, J.; He, J.; Mao, X.; Luo, Y.; Luo, J.; Huang, Z.; et al. Effects of dietary aged maize with oxidized fish oil on growth performance, antioxidant capacity and intestinal health in weaned piglets. Animals 2019, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Cardoso Dal Pont, G.; Farnell, M.; Farnell, Y.; Kogut, M.H. Dietary factors as triggers of low-grade chronic intestinal inflammation in poultry. Microorganisms 2020, 8, 139. [Google Scholar] [CrossRef]
- Carlson, D.A.; Smith, A.R.; Fischer, S.J.; Young, K.L.; Packer, L. The plasma pharmacokinetics of R-(+)-lipoic acid administered as sodium R-(+)-lipoate to healthy human subjects. Altern. Med. Rev. 2007, 12, 343–351. [Google Scholar] [PubMed]
- Celebioglu, A.; Uyar, T. Encapsulation and stabilization of α-lipoic acid in cyclodextrin inclusion complex electrospun nanofibers: Antioxidant and fast-dissolving α-lipoic acid/cyclodextrin nanofibrous webs. J. Agric. Food Chem. 2019, 67, 13093–13107. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Ghibu, S.; Muresan, A.; Vergely, C. Alpha-lipoic acid: Molecular mechanisms and therapeutic potential in diabetes. Can. J. Physiol. Pharmacol. 2015, 93, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Papanas, N.; Ziegler, D. Efficacy of α-lipoic acid in diabetic neuropathy. Expert Opin. Pharmacother. 2014, 15, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Bao, Y.H.; Zhang, Y.; Ji, C.; Zhao, L.H.; Zhang, J.Y.; Gao, C.Q.; Ma, Q.G. Effects of dietary α-lipoic acid, acetyl-l-carnitine, and sex on antioxidative ability, energy, and lipid metabolism in broilers. Poult. Sci. 2014, 93, 2809–2817. [Google Scholar] [CrossRef]
- Theodosis-Nobelos, P.; Papagiouvannis, G.; Tziona, P.; Rekka, E.A. Lipoic acid. Kinetics and pluripotent biological properties and derivatives. Mol. Biol. Rep. 2021, 48, 6539–6550. [Google Scholar] [CrossRef]
- Queiroz, T.M.; Guimarães, D.D.; Mendes-Junior, L.G.; Braga, V.A. α-Lipoic acid reduces hypertension and increases baroreflex sensitivity in renovascular hypertensive rats. Molecules 2012, 17, 13357–13367. [Google Scholar] [CrossRef]
- Hermann, R.; Mungo, J.; Cnota, P.J.; Ziegler, D. Enantiomer-selective pharmacokinetics, oral bioavailability, and sex effects of various alpha-lipoic acid dosage forms. Clin. Pharmacol. 2014, 6, 195–204. [Google Scholar] [CrossRef]
- Zimmer, G.; Beikler, T.K.; Schneider, M.; Ibel, J.; Tritschler, H.; Ulrich, H. Dose/response curves of lipoic acid R-and S-forms in the working rat heart during reoxygenation: Superiority of the R-enantiomer in enhancement of aortic flow. J. Mol. Cell. Cardiol. 1995, 27, 1895–1903. [Google Scholar] [CrossRef]
- Tomassoni, D.; Amenta, F.; Di Cesare Mannelli, L.; Ghelardini, C.; Nwankwo, I.E.; Pacini, A.; Tayebati, S.K. Neuroprotective activity of thioctic acid in central nervous system lesions consequent to peripheral nerve injury. BioMed Res. Int. 2013, 2013, 985093. [Google Scholar] [CrossRef]
- Pacini, A.; Tomassoni, D.; Trallori, E.; Micheli, L.; Amenta, F.; Ghelardini, C.; Di Cesare Mannelli, L.; Traini, E. Comparative assessment of the activity of racemic and dextrorotatory forms of thioctic (Alpha-Lipoic) acid in low back pain: Preclinical results and clinical evidences from an open randomized trial. Front. Pharmacol. 2021, 12, 607572. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, W.; Zhang, J.; Zhao, L.; Ji, C.; Zhang, J.; Huang, S.; Ma, Q. Lipoamide alleviates oxidized fish oil-induced host inflammatory response and oxidative damage in the oviduct of laying hens. Front. Vet. Sci. 2022, 9, 875769. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.A.; He, H.; Pham-Huy, C. Chiral drugs: An overview. Int. J. Biomed. Sci. 2006, 2, 85–100. [Google Scholar] [PubMed]
- Lee, G.R.; Hyun, M.H. Liquid chromatographic resolution of fendiline and its analogues on a chiral stationary phase based on (+)-(18-crown-6)-2,3,11,12-tetracarboxylic acid. Molecules 2014, 19, 21386–21397. [Google Scholar] [CrossRef]
- Wilkinson, J.; Foretia, D.; Rossington, S.; Heagerty, A.; Leonard, J.; Hussain, N.; Austin, C. 2′-hydroxy-fendiline analogues as potent relaxers of isolated arteries. Eur. J. Pharmacol. 2007, 561, 160–163. [Google Scholar] [CrossRef]
- Bhatia, P.A.; Moaddel, R.; Wainer, I.W. The synthesis and characterization of cellular membrane affinity chromatography columns for the study of human multidrug resistant proteins MRP1, MRP2 and human breast cancer resistant protein BCRP using membranes obtained from Spodoptera frugiperda (Sf9) insect cells. Talanta 2010, 81, 1477–1481. [Google Scholar] [CrossRef][Green Version]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Et Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef]
- Flora, S.J. Structural, chemical and biological aspects of antioxidants for strategies against metal and metalloid exposure. Oxidative Med. Cell. Longev. 2009, 2, 191–206. [Google Scholar] [CrossRef]
- Hong, Y.S.; Jacobia, S.J.; Packer, L.; Patel, M.S. The inhibitory effects of lipoic compounds on mammalian pyruvate dehydrogenase complex and its catalytic components. Free Radic. Biol. Med. 1999, 26, 685–694. [Google Scholar] [CrossRef]
- Yamada, T.; Hashida, K.; Takarada-Iemata, M.; Matsugo, S.; Hori, O. α-Lipoic acid (LA) enantiomers protect SH-SY5Y cells against glutathione depletion. Neurochem. Int. 2011, 59, 1003–1009. [Google Scholar] [CrossRef]
- Tomassoni, D.; Amenta, F.; Amantini, C.; Farfariello, V.; Di Cesare Mannelli, L.; Nwankwo, I.E.; Marini, C.; Tayebati, S.K. Brain activity of thioctic Acid enantiomers: In vitro and in vivo studies in an animal model of cerebrovascular injury. Int. J. Mol. Sci. 2013, 14, 4580–4595. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Taylor, C.; Lapidus, J.; Ramsey, K.; Koop, D.; Spain, R. Gastrointestinal tolerability and absorption of R- Versus R,S-lipoic acid in progressive multiple sclerosis: A randomized crossover trial. J. Clin. Pharmacol. 2020, 60, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Nobari, H.; Saedmocheshi, S.; Chung, L.H.; Suzuki, K.; Maynar-Mariño, M.; Pérez-Gómez, J. An overview on how exercise with green tea consumption can prevent the production of reactive oxygen species and improve sports performance. Int. J. Environ. Res. Public Health 2021, 19, 218. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, P.; Yin, J.; Jin, S.; Su, W.; Tian, J.; Li, T.; Yao, K. Effects of ornithine α-ketoglutarate on growth performance and gut microbiota in a chronic oxidative stress pig model induced by d-galactose. Food Funct. 2020, 11, 472–482. [Google Scholar] [CrossRef]
- Chen, Z.; Xing, T.; Li, J.; Zhang, L.; Jiang, Y.; Gao, F. Oxidative stress induced by hydrogen peroxide promotes glycolysis by activating CaMKK/LKB1/AMPK pathway in broiler breast muscle. Poult. Sci. 2022, 101, 101681. [Google Scholar] [CrossRef]
- Nawab, A.; Li, G.; An, L.; Wu, J.; Chao, L.; Xiao, M.; Zhao, Y.; Birmani, M.W.; Ghani, M.W. Effect of curcumin supplementation on TLR4 mediated non-specific immune responses in liver of laying hens under high-temperature conditions. J. Therm. Biol. 2019, 84, 384–397. [Google Scholar] [CrossRef]
- Zhang, Y.; Mahmood, T.; Tang, Z.; Wu, Y.; Yuan, J. Effects of naturally oxidized corn oil on inflammatory reaction and intestinal health of broilers. Poult. Sci. 2022, 101, 101541. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Larroquet, L.; Dias, K.; Cluzeaud, M.; Heraud, C.; Corlay, D. Effects of dietary oxidized fish oil supplementation on oxidative stress and antioxidant defense system in juvenile rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2018, 74, 43–51. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, H.J.; Wu, S.G.; Qiu, K.; Fu, Y.; Qi, G.H.; Wang, J. Supplemental xylooligosaccharide modulates intestinal mucosal barrier and cecal microbiota in laying hens fed oxidized fish oil. Front. Microbiol. 2021, 12, 635333. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Xie, J.; Ge, X.; Zhou, Q.; Sun, C.; Zhang, H.; Shan, F.; Yang, Z. Oxidized fish oil injury stress in Megalobrama amblycephala: Evaluated by growth, intestinal physiology, and transcriptome-based PI3K-Akt/NF-κB/TCR inflammatory signaling. Fish Shellfish Immunol. 2018, 81, 446–455. [Google Scholar] [CrossRef]
- Wang, J.; Yue, H.; Wu, S.; Zhang, H.; Qi, G. Nutritional modulation of health, egg quality and environmental pollution of the layers. Anim. Nutr. 2017, 3, 91–96. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.; Celi, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Feng, B.; Xu, S.; et al. Alteration of the antioxidant capacity and gut microbiota under high levels of molybdenum and green tea polyphenols in laying hens. Antioxidants 2019, 8, 503. [Google Scholar] [CrossRef]
- Wang, J.P.; He, K.R.; Ding, X.M.; Luo, Y.H.; Bai, S.P.; Zeng, Q.F.; Su, Z.W.; Xuan, Y.; Zhang, K.Y. Effect of dietary vanadium and vitamin C on egg quality and antioxidant status in laying hens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Sohaib, M.; Anjum, F.M.; Nasir, M.; Saeed, F.; Arshad, M.S.; Hussain, S. Alpha-lipoic acid: An inimitable feed supplement for poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2018, 102, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Lv, Z.; Hu, C.; Shi, Z.; Wei, X.; Jin, S.; Yuan, Y.; Yu, D.; Shi, F. Alpha-lipoic acid improves the reproduction performance of breeder hens during the late egg-laying period. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1788–1797. [Google Scholar] [CrossRef] [PubMed]
- Stawiarska-Pięta, B.; Zalejska-Fiolka, J.; Wyszyńska, M.; Kleczka, A.; Janiga, B.; Grzegorzak, N.; Birkner, E. Influence of α-lipoic acid on morphology of organs of rabbits fed a high fat diet with the addition of oxidised rapeseed oil. J. Vet. Res. 2017, 61, 517–525. [Google Scholar] [CrossRef][Green Version]
- Li, M.; Wu, X.; Zou, J.; Lai, Y.; Niu, X.; Chen, X.; Kong, Y.; Wang, G. Dietary α-lipoic acid alleviates deltamethrin-induced immunosuppression and oxidative stress in northern snakehead (Channa argus) via Nrf2/NF-κB signaling pathway. Fish Shellfish Immunol. 2022, 127, 228–237. [Google Scholar] [CrossRef]
- Zalejska-Fiolka, J.; Wielkoszyński, T.; Rokicki, W., Jr.; Dąbrowska, N.; Strzelczyk, J.K.; Kasperczyk, A.; Owczarek, A.; Błaszczyk, U.; Kasperczyk, S.; Stawiarska-Pięta, B.; et al. The influence of α-lipoic acid and garlic administration on biomarkers of oxidative stress and inflammation in rabbits exposed to oxidized nutrition oils. BioMed Res. Int. 2015, 2015, 827879. [Google Scholar] [CrossRef]
- Skibska, B.; Goraca, A.; Skibska, A.; Stanczak, A. Effect of alpha-lipoic acid on rat ventricles and atria under LPS-induced oxidative stress. Antioxidants 2022, 11, 734. [Google Scholar] [CrossRef]
- Piechota-Polanczyk, A.; Fichna, J. Review article: The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef]
- Oh, W.S.; Jung, J.C.; Choi, Y.M.; Mun, J.Y.; Ku, S.K.; Song, C.H. Protective effects of fermented rice extract on ulcerative colitis induced by dextran sodium sulfate in mice. Food Sci. Nutr. 2020, 8, 1718–1728. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Guo, J.; Sun, H.; Huang, Y.; Zhao, R.; Yang, X. α-Lipoic acid attenuates LPS-induced liver injury by improving mitochondrial function in association with GR mitochondrial DNA occupancy. Biochimie 2015, 116, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Nielson, C.B.; Duethman, N.C.; Howard, J.M.; Moncure, M.; Wood, J.G. Burns: Pathophysiology of systemic complications and current management. J. Burn Care Res. 2017, 38, e469–e481. [Google Scholar] [CrossRef] [PubMed]
- Wéra, O.; Lancellotti, P.; Oury, C. The dual role of neutrophils in inflammatory bowel diseases. J. Clin. Med. 2016, 5, 118. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Gharib, D.M.; Reyad, H.R.; Mohamed, A.A.; Elroby, F.A.; Mahmoud, H.S. Antioxidant and anti-inflammatory properties of alpha-lipoic acid protect against valproic acid-induced liver injury. Can. J. Physiol. Pharmacol. 2021, 99, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Di Tucci, C.; Di Feliciantonio, M.; Vena, F.; Capone, C.; Schiavi, M.C.; Pietrangeli, D.; Muzii, L.; Benedetti Panici, P. Alpha lipoic acid in obstetrics and gynecology. Gynecol. Endocrinol. 2018, 34, 729–733. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.G.; Zhao, L.H.; Wei, H.; Duan, G.X.; Zhang, J.Y.; Ji, C. Effects of lipoic acid on immune function, the antioxidant defense system, and inflammation-related genes expression of broiler chickens fed aflatoxin contaminated diets. Int. J. Mol. Sci. 2014, 15, 5649–5662. [Google Scholar] [CrossRef]
- Bonenfant, M.; Blomquist, C.H.; Provost, P.R.; Drolet, R.; D’Ascoli, P.; Tremblay, Y. Tissue- and site-specific gene expression of type 2 17beta-hydroxysteroid dehydrogenase: In Situ hybridization and specificenzymatic activity studies in human placental endothelial cells of the arterial system. J. Clin. Endocrinol. Metab. 2000, 85, 4841–4850. [Google Scholar] [CrossRef][Green Version]
- Hazarika, J.; Ganguly, M.; Mahanta, R. A computational insight into the molecular interactions of chlorpyrifos and its degradation products with the human progesterone receptor leading to endocrine disruption. J. Appl. Toxicol. 2019, 40, 434–443. [Google Scholar] [CrossRef]
- Othman, A.I.; El-Missiry, M.A.; Koriem, K.M.; El-Sayed, A.A. Alfa-lipoic acid protects testosterone secretion pathway and sperm quality against 4-tert-octylphenol induced reproductive toxicity. Ecotoxicol. Environ. Saf. 2012, 81, 76–83. [Google Scholar] [CrossRef]
- Lebda, M.; Gad, S.; Gaafar, H. Effects of lipoic acid on acrylamide induced testicular damage. Mater. Socio-Med. 2014, 26, 208–212. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Egg Laying Rate/% | Egg Weight/g | Egg Production/g/d | Feed Intake/g | Weight of Laying Hen/kg | Feed/Egg | |
|---|---|---|---|---|---|---|
| CON | 73.51 a,b | 64.80 | 46.16 | 114.21 | 1.63 | 2.49 |
| OFO | 61.21 a | 66.42 | 46.95 | 115.88 | 1.66 | 2.52 |
| OFO + S-LA | 67.86 a,b | 66.71 | 47.59 | 117.32 | 1.75 | 2.62 |
| OFO + R-LA | 77.53 b | 69.70 | 47.98 | 118.60 | 1.82 | 2.71 |
| p value | 0.057 | 0.229 | 0.956 | 0.414 | 0.096 | 0.81 |
| SEM | 2.29 | 0.85 | 1.17 | 0.95 | 0.029 | 0.085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Li, W.; Huang, S.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. R- Is Superior to S-Form of α-Lipoic Acid in Anti-Inflammatory and Antioxidant Effects in Laying Hens. Antioxidants 2022, 11, 1530. https://doi.org/10.3390/antiox11081530
Liu Q, Li W, Huang S, Zhao L, Zhang J, Ji C, Ma Q. R- Is Superior to S-Form of α-Lipoic Acid in Anti-Inflammatory and Antioxidant Effects in Laying Hens. Antioxidants. 2022; 11(8):1530. https://doi.org/10.3390/antiox11081530
Chicago/Turabian StyleLiu, Qingxiu, Wenxiang Li, Shimeng Huang, Lihong Zhao, Jianyun Zhang, Cheng Ji, and Qiugang Ma. 2022. "R- Is Superior to S-Form of α-Lipoic Acid in Anti-Inflammatory and Antioxidant Effects in Laying Hens" Antioxidants 11, no. 8: 1530. https://doi.org/10.3390/antiox11081530
APA StyleLiu, Q., Li, W., Huang, S., Zhao, L., Zhang, J., Ji, C., & Ma, Q. (2022). R- Is Superior to S-Form of α-Lipoic Acid in Anti-Inflammatory and Antioxidant Effects in Laying Hens. Antioxidants, 11(8), 1530. https://doi.org/10.3390/antiox11081530