
Natural Astaxanthin Improves Testosterone Synthesis and Sperm Mitochondrial Function in Aging Roosters

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Reproductive Hormones Analysis
2.4. Antioxidant Enzymes and Free Oxygen Radical Analysis
2.5. Sperm Mitochondrial Preparation
2.6. Oxidative Phosphorylation (OXPHOS) Complexes Analysis
2.7. Sperm MMP, ROS Production, and Apoptosis Analysis
2.8. Caspase Activity Analysis
2.9. Testes Histology Analysis
2.10. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
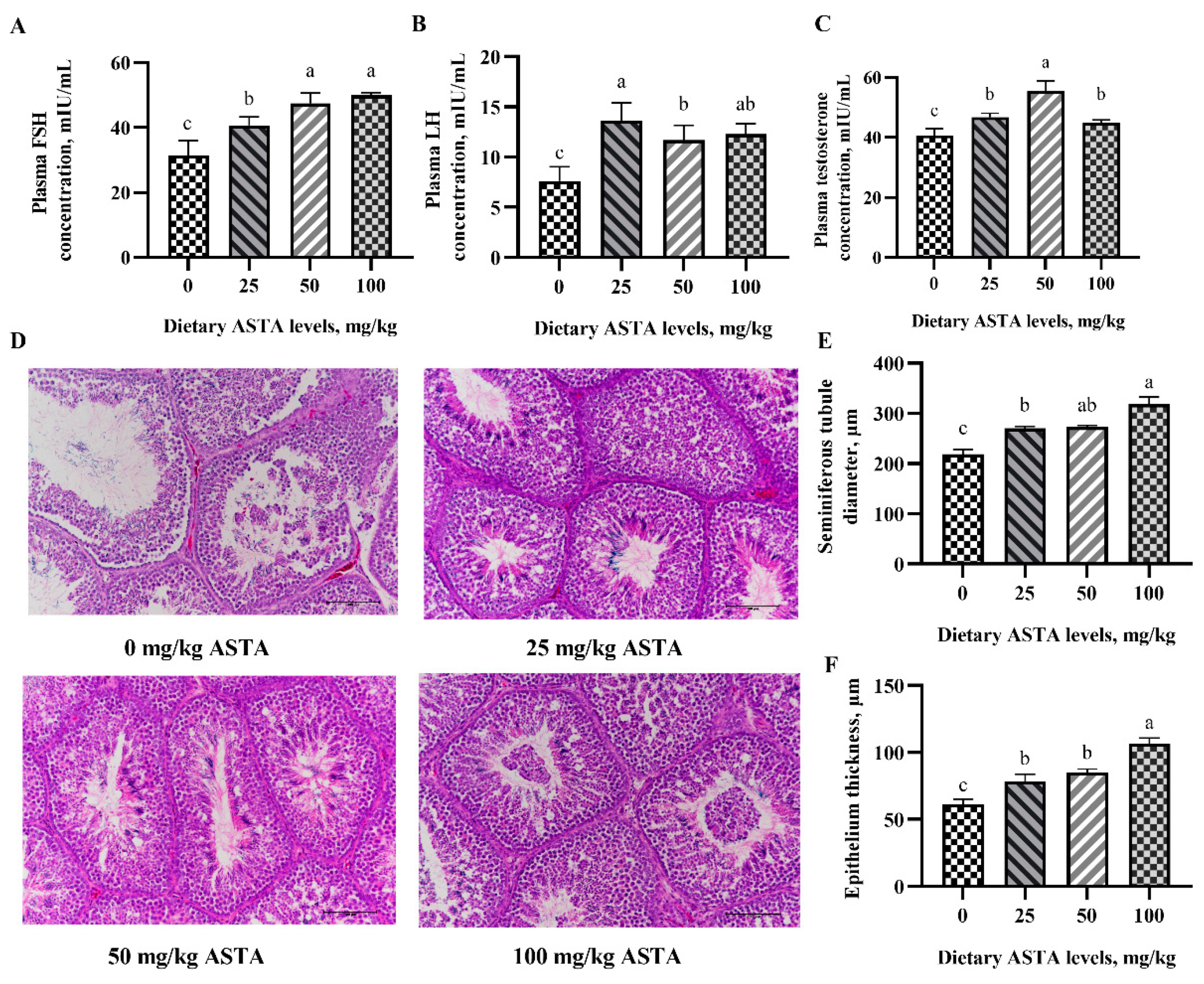
3.1. Reproductive Hormones Level and Testes Histology
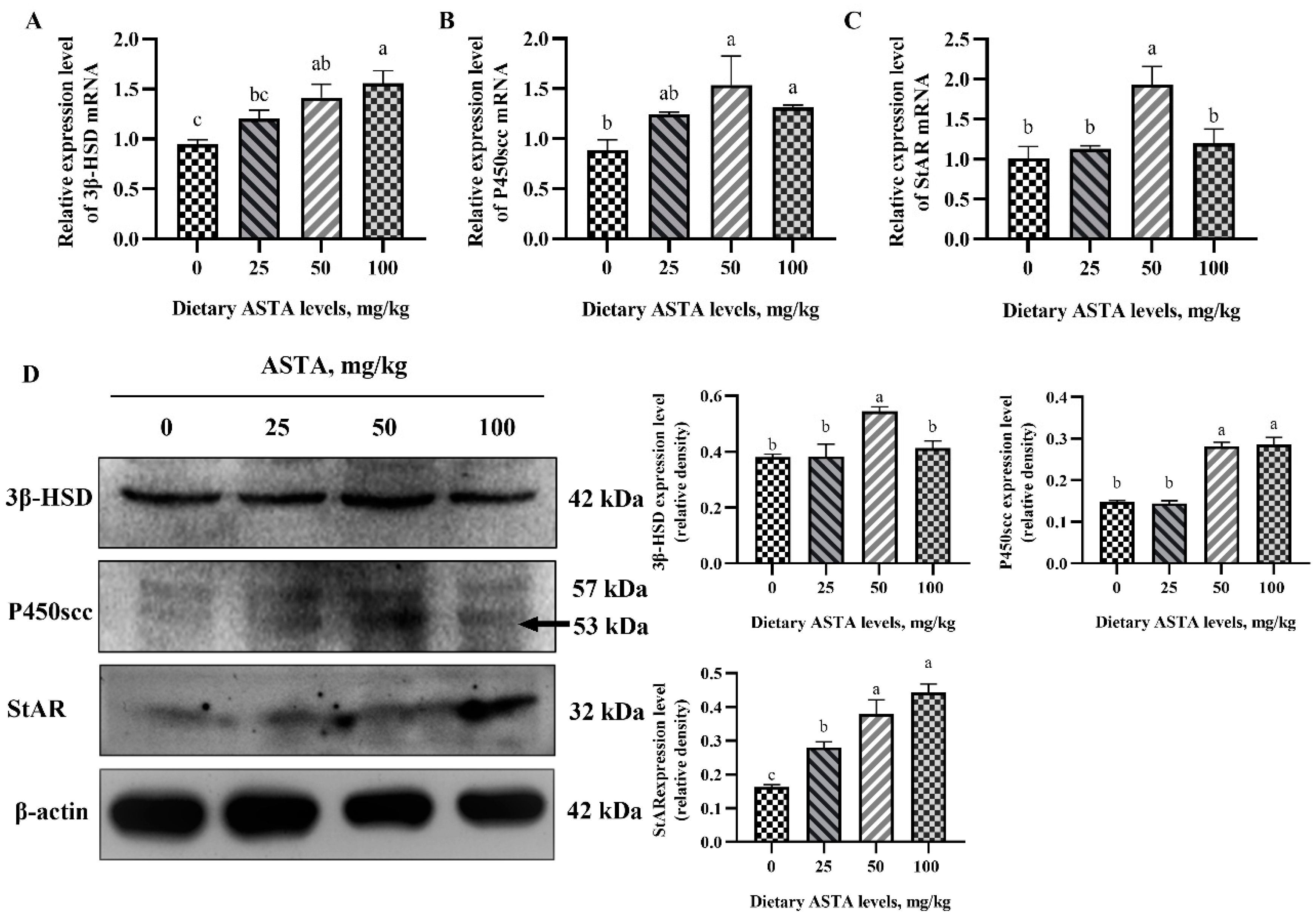
3.2. Expression of Genes Involved in the Testosterone Synthesis Pathway at the mRNA and Protein Levels
3.3. Antioxidant Ability of Sperm
3.4. OXPHOS Complex Activity
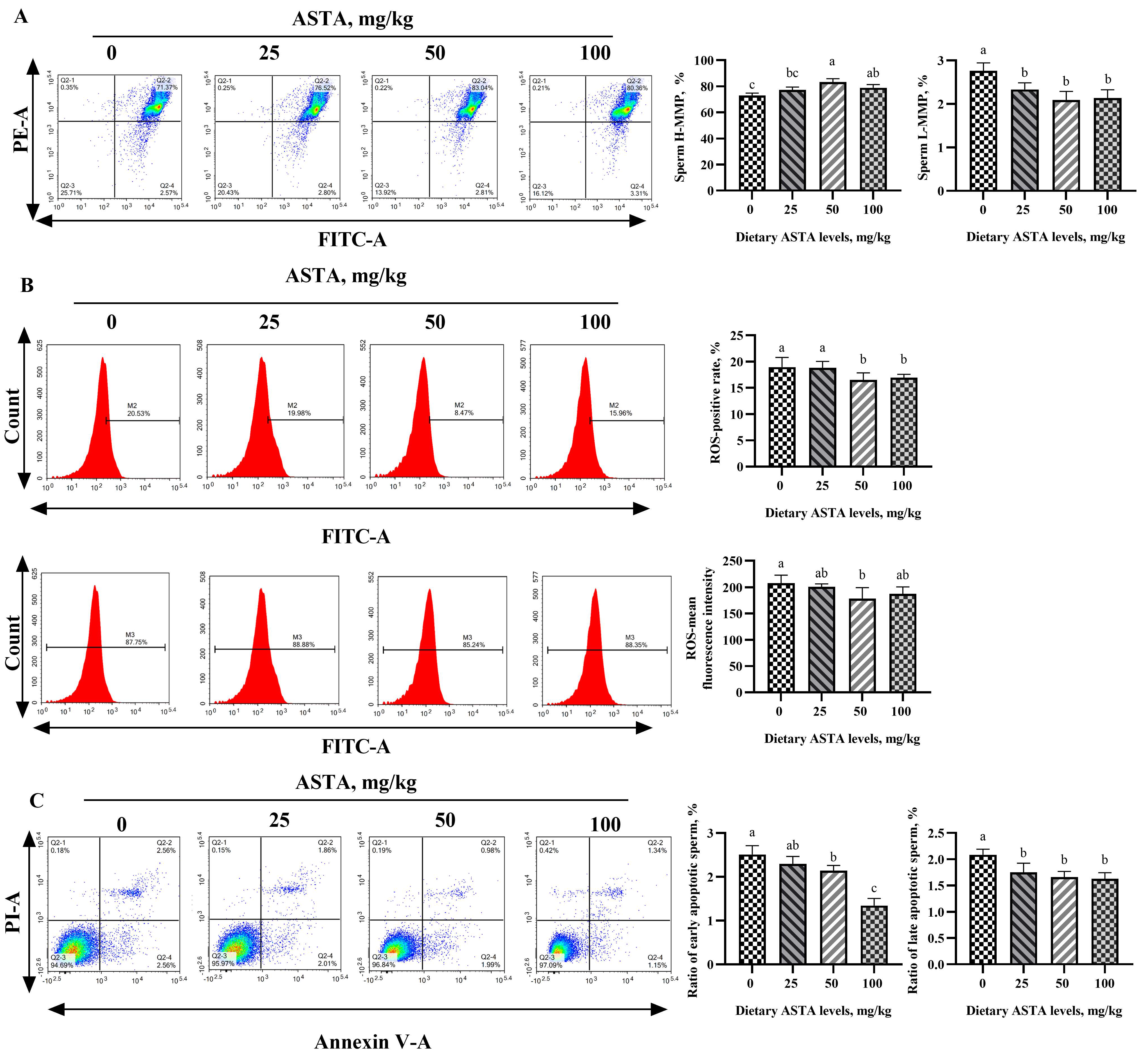
3.5. MMP, ROS, and Apoptosis Levels
3.6. Caspase-3 and Caspase-9 Activation and the mRNA Expression of Apoptosis Factors
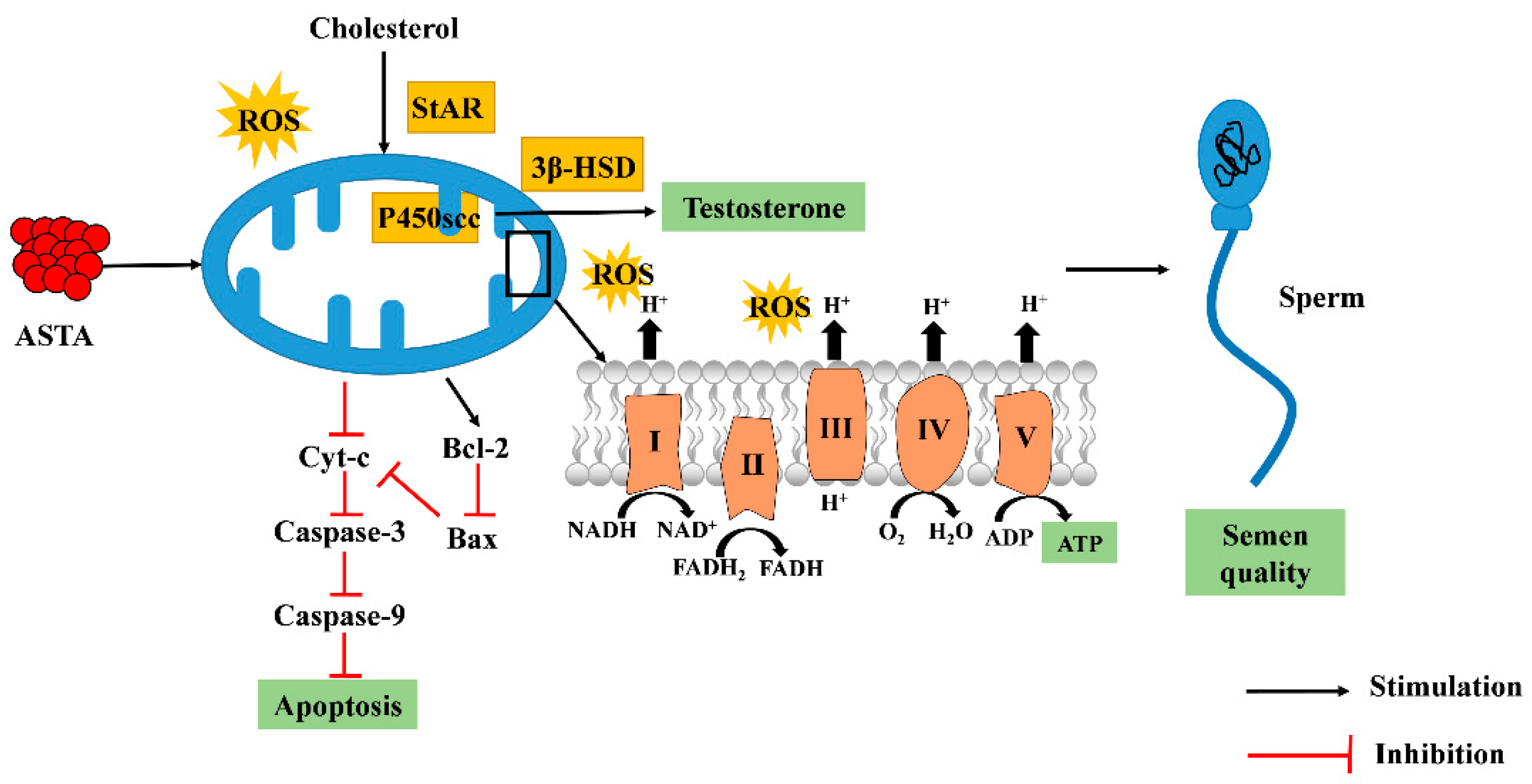
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
References
- Beattie, M.C.; Adekola, L.; Papadopoulos, V.; Chen, H.; Zirkin, B.R. Leydig cell aging and hypogonadism. Exp. Gerontol. 2015, 68, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xue, F.; Li, Y.; Fu, L.; Bai, H.; Ma, H.; Xu, S.; Chen, J. Differences in semen quality, testicular histomorphology, fertility, reproductive hormone levels, and expression of candidate genes according to sperm motility in Beijing-You chickens. Poult. Sci. 2019, 98, 4182–4189. [Google Scholar] [CrossRef] [PubMed]
- Rebourcet, D.; Mackay, R.; Darbey, A.; Curley, M.K.; Jørgensen, A.; Frederiksen, H.; Mitchell, R.T.; O’Shaughnessy, P.J.; Nef, S.; Smith, L.B. Ablation of the canonical testosterone production pathway via knockout of the steroidogenic enzyme HSD17B3, reveals a novel mechanism of testicular testosterone production. FASEB J. 2020, 34, 10373–10386. [Google Scholar] [CrossRef]
- Ansari, M.; Zhandi, M.; Kohram, H.; Zaghari, M.; Sadeghi, M.; Sharafi, M. Improvement of post-thawed sperm quality and fertility of Arian rooster by oral administration of d-aspartic acid. Theriogenology 2017, 92, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Yuan, Z.; Hou, J.; Zhao, J.; Liu, H.; Lu, W.; Wang, J. Prolonging photoperiod promotes testosterone synthesis of Leydig cells by directly targeting local melatonin system in rooster testes. Biol. Reprod. 2021, 105, 1317–1329. [Google Scholar] [CrossRef]
- Ansari, M.; Zhandi, M.; Kohram, H.; Zaghari, M.; Sadeghi, M.; Gholami, M.; Deldar, H.; Di Fiore, M.M.; Benson, A.P. D-Aspartate amends reproductive performance of aged roosters by changing gene expression and testicular histology. Reprod. Fertil. Dev. 2018, 30, 1038. [Google Scholar] [CrossRef]
- Jiang, H.; Zhu, W.; Li, J.; Chen, Q.; Liang, W.; Gu, Y. Quantitative histological analysis and ultrastructure of the aging human testis. Int. Urol. Nephrol. 2014, 46, 879–885. [Google Scholar] [CrossRef]
- Xia, M.F.; Zhang, Y.Z.; Jin, K.; Lu, Z.T.; Xiong, W. Communication between mitochondria and other organelles: A brand-new perspective on mitochondria in cancer. Cell Biosci. 2019, 9, 27. [Google Scholar] [CrossRef]
- Durairajanayagam, D.; Singh, D.; Agarwal, A.; Henkel, R. Causes and consequences of sperm mitochondrial dysfunction. Andrologia 2021, 53, e13666. [Google Scholar] [CrossRef]
- Losano, J.O.D.A.; Fernando, P.J.; Iago, M.; Angrimani, D.S.R.; García, G.A.; Barnabe, V.H.; Marcilio, N. The Stimulated Glycolytic Pathway Is Able to Maintain ATP Levels and Kinetic Patterns of Bovine Epididymal Sperm Subjected to Mitochondrial Uncoupling. Oxidative Med. Cell. Longev. 2017, 2017, 1682393. [Google Scholar] [CrossRef] [Green Version]
- Moraes, C.R.; Meyers, S. The sperm mitochondrion: Organelle of many functions. Anim. Reprod. Sci. 2018, 194, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Sharbatoghli, M.; Valojerdi, M.R.; Amanlou, M.; Khosravi, F.; Jafar-Abadi, M.A. Relationship of sperm DNA fragmentation, apoptosis and dysfunction of mitochondrial membrane potential with semen parameters and ART outcome after intracytoplasmic sperm injection. Arch. Gynecol. Obstet. 2012, 286, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Zhang, L.; Zhang, D.; Jin, H. L-Cystathionine Protects against Homocysteine-Induced Mitochondria-Dependent Apoptosis of Vascular Endothelial Cells. Oxidative Med. Cell. Longev. 2019, 2019, 1253289. [Google Scholar] [CrossRef]
- Gao, J.; Shuang, L.; Fan, X.; Liu, Y.; Lv, C.; Yan, D.; Shi, J.; Gong, Q. Trilobatin Protects Against Oxidative Injury in Neuronal PC12 Cells Through Regulating Mitochondrial ROS Homeostasis Mediated by AMPK/Nrf2/Sirt3 Signaling Pathway. Front. Mol. Neurosci. 2018, 11, 267. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Najafi, A.; Taheri, R.A.; Mehdipour, M.; Farnoosh, G.; Martínez-Pastor, F. Lycopene-loaded nanoliposomes improve the performance of a modified Beltsville extender broiler breeder roosters. Anim. Reprod. Sci. 2018, 195, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Kanno, C.; Oshima, E.; Kuzuma, Y.; Kim, S.W.; Bai, H.; Takahashi, M.; Yanagawa, Y.; Nagano, M.; Wakamatsu, J.; et al. Enhancement of sperm motility and viability by turmeric by-product dietary supplementation in roosters. Anim. Reprod. Sci. 2017, 185, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M.A. Antioxidant Activities of Astaxanthin and Related Carotenoids. J. Agr. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Sajad, F.; Fatemeh, A.; Leila, D.; Masoumeh, J. Astaxanthin: A Mechanistic Review on its Biological Activities and Health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar]
- Janina, D.; Seiichi, M.; Haruka, Y.; Yutaro, K.; Shigetoshi, O.; Ulrike, S.; Manfred, E.; Gerald, R.; Tuba, E. Free Radical Scavenging and Cellular Antioxidant Properties of Astaxanthin. Int. J. Mol. Sci. 2016, 17, 103. [Google Scholar]
- Iskender, H.; Yenice, G.; Dokumacioglu, E.; Hayirli, A.; Sevim, C.; Dokumacioglu, A.; Terim, K.K. Astaxanthin alleviates renal damage of rats on high fructose diet through modulating NFkappaB/SIRT1 pathway and mitigating oxidative stress. Arch. Physiol. Biochem. 2020, 126, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H. Multiple Mechanisms of Anti-Cancer Effects Exerted by Astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Heng, N.; Liu, F.; Guo, Y.; Chen, Y.; Wang, L.; Ni, H.; Sheng, X.; Wang, X.; Xing, K.; et al. Natural astaxanthin enhanced antioxidant capacity and improved semen quality through the MAPK/Nrf2 pathway in aging layer breeder roosters. J. Anim. Sci. Biotechnol. 2021, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Xiaolong Qi, S.W.H.Z. Effects of dietary conjugated linoleic acids on lipid metabolism and antioxidant capacity in laying hens. Arch. Anim. Nutr. 2011, 65, 354–365. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ni, C.; Li, Y.; Li, Z.; Tian, L.; Fu, J.; Wu, K.; Wang, Y.; Yao, M.; Ge, R.S. Cisatracurium stimulates testosterone synthesis in rat and mouse Leydig cells via nicotinic acetylcholine receptor. J. Cell. Mol. Med. 2020, 24, 14184–14194. [Google Scholar] [CrossRef]
- Chen, P.; Zirkin, B.R.; Chen, H. Leydig Cells in the Adult Testis: Characterization, Regulation and Potential Applications. Endocr. Rev. 2020, 41, 22–32. [Google Scholar] [CrossRef]
- Heng, D.; Zhang, T.; Tian, Y.; Yu, S.; Liu, W.; Xu, K.; Liu, J.; Ding, Y.; Zhu, B.; Yang, Y.; et al. Effects of dietary soybean isoflavones (SI) on reproduction in the young breeder rooster. Anim. Reprod. Sci. 2017, 177, 124–131. [Google Scholar] [CrossRef]
- Elokil, A.A.; Bhuiyan, A.A.; Liu, H.Z.; Hussein, M.N.; Ahmed, H.I.; Azmal, S.A.; Yang, L.; Li, S. The capability of L-carnitine-mediated antioxidant on cock during aging: Evidence for the improved semen quality and enhanced testicular expressions of GnRH1, GnRHR, and melatonin receptors MT 1/2. Poult. Sci. 2019, 98, 4172–4181. [Google Scholar] [CrossRef]
- Qi, X.; Shang, M.; Chen, C.; Chen, Y.; Hua, J.; Sheng, X.; Wang, X.; Xing, K.; Ni, H.; Guo, Y. Dietary supplementation with linseed oil improves semen quality, reproductive hormone, gene and protein expression related to testosterone synthesis in aging layer breeder roosters. Theriogenology 2019, 131, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.; Lu, C.; Lin, C.; Wang, P.S. Antisteroidogenic actions of hydrogen peroxide on rat leydig cells. J. Cell. Biochem. 2003, 90, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Diemer, T.; Allen, J.A.; Hales, K.H.; Hales, D.B. Reactive Oxygen Disrupts Mitochondria in MA-10 Tumor Leydig Cells and Inhibits Steroidogenic Acute Regulatory (StAR) Protein and Steroidogenesis. Endocrinology 2003, 144, 2882–2891. [Google Scholar] [CrossRef] [PubMed]
- Dale, B.H.; John, A.A.; Tristan, S.; Paul, J.; Steve, B.; Thorsten, D.; Hales, K.H. Mitochondrial Function in Leydig Cell Steroidogenesis. Testic. Cell Dyn. Endocr. Signal. 2005, 1061, 120–134. [Google Scholar]
- Wang, J.; Lee, Y.; Chou, M.; Chang, R.; Chiu, C.; Liang, Y.; Wu, L. Astaxanthin Protects Steroidogenesis from Hydrogen Peroxide-Induced Oxidative Stress in Mouse Leydig Cells. Mar. Drugs 2015, 13, 1375–1388. [Google Scholar] [CrossRef] [PubMed]
- Seifried, H.E.; Anderson, D.E.; Fisher, E.I.; Milner, J.A. A review of the interaction among dietary antioxidants and reactive oxygen species. J. Nutr. Biochem. 2007, 18, 567–579. [Google Scholar] [CrossRef]
- Heng, N.; Gao, S.; Chen, Y.; Wang, L.; Li, Z.; Guo, Y.; Sheng, X.; Wang, X.; Xing, K.; Xiao, L.; et al. Dietary supplementation with natural astaxanthin from Haematococcus pluvialis improves antioxidant enzyme activity, free radical scavenging ability, and gene expression of antioxidant enzymes in laying hens. Poult. Sci. 2021, 100, 101045. [Google Scholar] [CrossRef]
- Hama, S.; Uenishi, S.; Yamada, A.; Ohgita, T.; Tsuchiya, H.; Yamashita, E.; Kogure, K.; Department, O.B.C.; Kyoto, P.U.; Fuji, C.I.C.; et al. Scavenging of Hydroxyl Radicals in Aqueous Solution by Astaxanthin Encapsulated in Liposomes. Biol. Pharm. Bull. 2012, 35, 2238–2242. [Google Scholar] [CrossRef]
- Ferrusola, C.O.; Fernández, L.G.; Sandoval, C.S.; García, B.M.; Martínez, H.R.; Tapia, J.A.; Peña, F.J. Inhibition of the mitochondrial permeability transition pore reduces “apoptosis like” changes during cryopreservation of stallion spermatozoa. Theriogenology 2010, 74, 458–465. [Google Scholar] [CrossRef]
- Frey, T.G.; Mannella, C.A. The internal structure of mitochondria. Trends Biochem. Sci. 2000, 25, 319–324. [Google Scholar] [CrossRef]
- Werner, K. Structure and function of mitochondrial membrane protein complexes. BMC Biol. 2015, 13, 89. [Google Scholar]
- Lenaz, G.; Genova, M.L. Structure and organization of mitochondrial respiratory complexes: A new understanding of an old subject. Antioxid. Redox Signal. 2010, 12, 961–1008. [Google Scholar] [CrossRef]
- Boyer, P.D. Toward an Adequate Scheme for the ATP Synthase Catalysis. Biochemistry 2001, 66, 1058–1066. [Google Scholar] [PubMed]
- Klingenberg, M. The ADP and ATP transport in mitochondria and its carrier. Biochim. Biophys. Acta 2008, 1778, 1978–2021. [Google Scholar] [CrossRef] [Green Version]
- Krämer, R. Structural and functional aspects of the phosphate carrier from mitochondria. Kidney Int. 1996, 49, 947–952. [Google Scholar]
- Kamali Sangani, A.; Masoudi, A.A.; Vaez Torshizi, R. Association of mitochondrial function and sperm progressivity in slow- and fast-growing roosters. Poult. Sci. 2017, 96, 211–219. [Google Scholar] [CrossRef]
- Ruiz-Pesini, E.; Lapeña, A.; Díez-Sánchez, C.; Pérez-Martos, A.; Montoya, J.; Alvarez, E.; Díaz, M.; Urriés, A.; Montoro, L.; López-Pérez, M.J.; et al. Human mtDNA Haplogroups Associated with High or Reduced Spermatozoa Motility. Am. J. Hum. Genet. 2000, 67, 682–696. [Google Scholar] [CrossRef]
- Quinlan, C.L.; Perevoshchikova, I.V.; Hey-Mogensen, M.; Orr, A.L.; Brand, M.D. Sites of reactive oxygen species generation by mitochondria oxidizing different substrates. Redox Biol. 2013, 1, 304–312. [Google Scholar] [CrossRef]
- Kuroki, T.; Ikeda, S.; Okada, T.; Maoka, T.; Kitamura, A.; Sugimoto, M.; Kume, S. Astaxanthin ameliorates heat stress-induced impairment of blastocyst development In Vitro: –astaxanthin colocalization with and action on mitochondria. J. Assist. Reprod. Gen. 2013, 30, 623–631. [Google Scholar] [CrossRef]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.R.; Chen, G.W.; Shi, H.J.; Wai-Sum, O.; Martin- De Leon, P.A.; Chen, H. Prohibitin involvement in the generation of mitochondrial superoxide at complex I in human sperm. J. Cell. Mol. Med. 2017, 21, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Iio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Brasacchio, D.; Noori, T.; House, C.; Brennan, A.J.; Simpson, K.J.; Susanto, O.; Bird, P.I.; Johnstone, R.W.; Trapani, J.A. A functional genomics screen identifies PCAF and ADA3 as regulators of human granzyme B-mediated apoptosis and Bid cleavage. Cell Death Differ. 2014, 21, 748–760. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wang, Y.; Xiong, Y.; Gao, W.; Teng, L. Salidroside alleviated hypoxia-induced liver injury by inhibiting endoplasmic reticulum stress-mediated apoptosis via IRE1α/JNK pathway. Biochem. Biophys. Res. Commun. 2020, 529, 335–340. [Google Scholar] [CrossRef]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Singh, K.K.; Rajender, S. Mitochondria, spermatogenesis, and male infertility—An update. Mitochondrion 2020, 54, 26–40. [Google Scholar]
- Yiğit, M.; Güneş, A.; Uğuz, C.; Yalçın, T.Ö.; Tök, L.; Öz, A.; Mustafa, N. Effects of astaxanthin on antioxidant parameters in ARPE-19 cells on oxidative stress model. Int. J. Ophthalmol. 2019, 12, 930–935. [Google Scholar]
- Paasch, U.; Grunewald, S.; Agarwal, A.; Glandera, H.J. Activation pattern of caspases in human spermatozoa. Fertil. Steril. 2004, 81, 802–809. [Google Scholar] [CrossRef]
- Rajender, S.; Rahul, P.; Mahdi, A.A. Mitochondria, spermatogenesis and male infertility. Mitochondrion 2010, 10, 419–428. [Google Scholar] [CrossRef]
- Wang, C.C.; Shi, H.H.; Xu, J.; Yanagita, T.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Docosahexaenoic acid-acylated astaxanthin ester exhibits superior performance over non-esterified astaxanthin in preventing behavioral deficits coupled with apoptosis in MPTP-induced mice with Parkinson’s disease. Food Funct. 2020, 11, 8038–8050. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.M.; El-Senousey, H.K.; Ruan, D.; Xia, W.; Chen, W.; Wang, S.; Zheng, C. Nutritional modulation of fertility in male poultry. Poult. Sci. 2020, 99, 5637–5646. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content, % | Nutrition Level | |
|---|---|---|---|
| Corn | 69.93 | Metabolizable energy, MJ·kg−1 | 12.02 |
| Soybean meal | 18.60 | Crude protein, % | 14.00 |
| Wheat bran | 3.80 | Methionine, % | 0.23 |
| Soybean oil | 1.40 | Lysine, % | 0.64 |
| Limestone | 2.60 | Calcium, % | 1.37 |
| Dicalcium phosphate | 1.80 | Total phosphorus, % | 0.63 |
| Salt | 0.25 | Available phosphorus, % | 0.44 |
| Choline chloride | 0.20 | ||
| Premix 1 | 1.42 | ||
| Total | 100.00 | ||
| Genes | Primer Sequence (5′-3′) | Product Size, bp | Accession Number |
|---|---|---|---|
| Caspase-3 | Forward: TGCTCCAGGCTACTACTCCT | 91 | NM_20472.51 |
| Reverse: TTTCCTGGCGTGTTCCTTCA | |||
| Caspase-9 | Forward: GTCCATCCCAGTCCAACCTG | 98 | XM_424580.6 |
| Reverse: GGTACACCAGTCTGTGGTCG | |||
| Bax | Forward: ACAGGGATCGTCACAGTCAT | 120 | XM_015290060.2 |
| Reverse: CACCAACTGTGTGTCGTAGG | |||
| Bcl-2 | Forward: TCGTCGCCTTCTTCGAGTTC | 150 | NM_205339.2 |
| Reverse: CATCCCATCCTCCGTTGTCC | |||
| Cyt-c | Forward: CCTGTCCTGGTGCATGATG | 77 | NM_205147.1 |
| Reverse: TACTCTGATCCAGCTCTGCCTGAA | |||
| 3β-HSD | Forward: GCAAGAGGCTGGCAGAGGAATG | 90 | NM_205118.2 |
| Reverse: GGTGACGGCGTCGATGAA | |||
| P450scc | Forward: GCTTTGCCTTGGAGTCTGTG | 104 | NM_001001756.2 |
| Reverse: GGTGACGGCGTCGATGAA | |||
| StAR | Forward: TCAGCCGGCGGATTTAAGG | 64 | NM_204686.2 |
| Reverse: TGGTGGCTGCTACAAACACT | |||
| ACTB | Forward: GCCAACAGAGAGAAGATGACAC | 118 | NM_205518 |
| Reverse: GTAACACCATCACCAGAGTCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Zhao, B.-X.; Long, C.; Heng, N.; Guo, Y.; Sheng, X.-H.; Wang, X.-G.; Xing, K.; Xiao, L.-F.; Ni, H.-M.; et al. Natural Astaxanthin Improves Testosterone Synthesis and Sperm Mitochondrial Function in Aging Roosters. Antioxidants 2022, 11, 1684. https://doi.org/10.3390/antiox11091684
Gao S, Zhao B-X, Long C, Heng N, Guo Y, Sheng X-H, Wang X-G, Xing K, Xiao L-F, Ni H-M, et al. Natural Astaxanthin Improves Testosterone Synthesis and Sperm Mitochondrial Function in Aging Roosters. Antioxidants. 2022; 11(9):1684. https://doi.org/10.3390/antiox11091684
Chicago/Turabian StyleGao, Shan, Bang-Xin Zhao, Cheng Long, Nuo Heng, Yong Guo, Xi-Hui Sheng, Xiang-Guo Wang, Kai Xing, Long-Fei Xiao, He-Min Ni, and et al. 2022. "Natural Astaxanthin Improves Testosterone Synthesis and Sperm Mitochondrial Function in Aging Roosters" Antioxidants 11, no. 9: 1684. https://doi.org/10.3390/antiox11091684