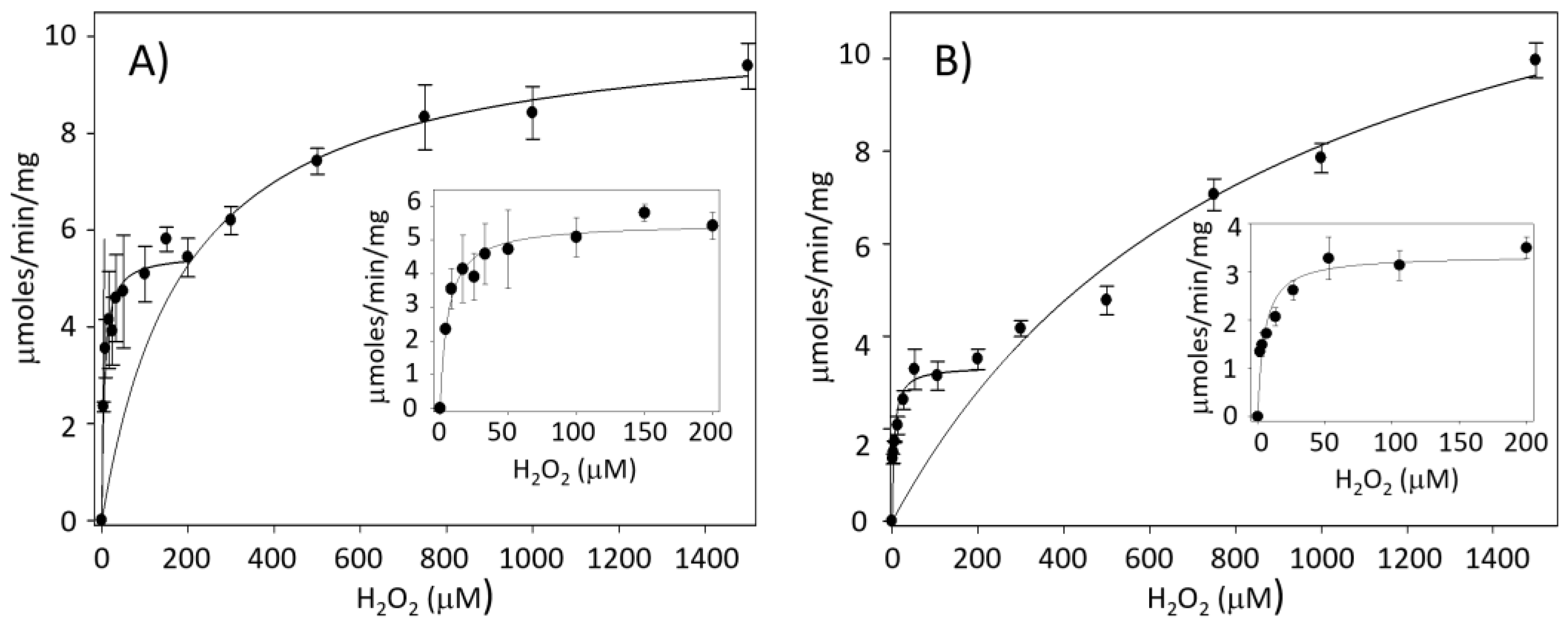
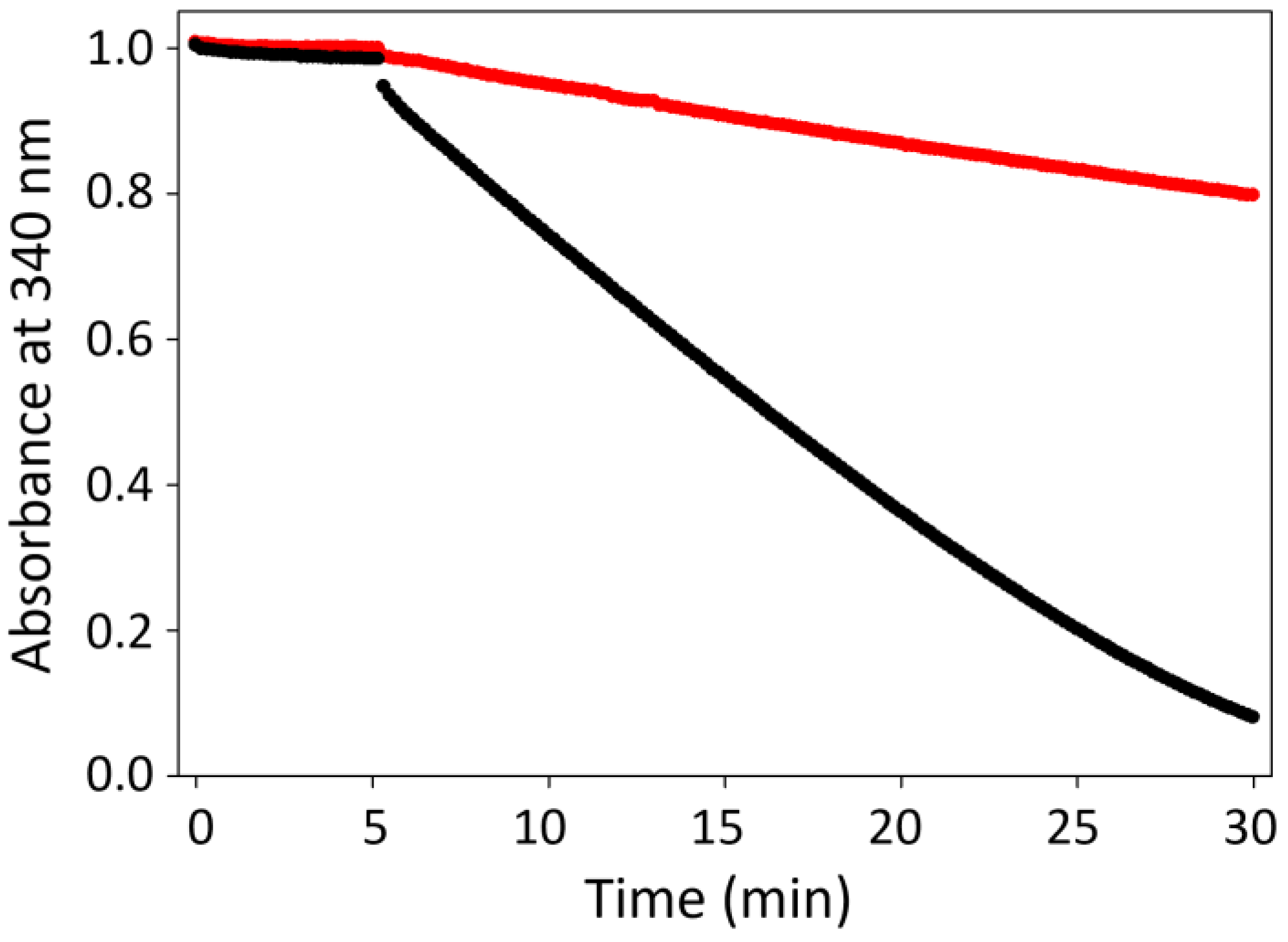
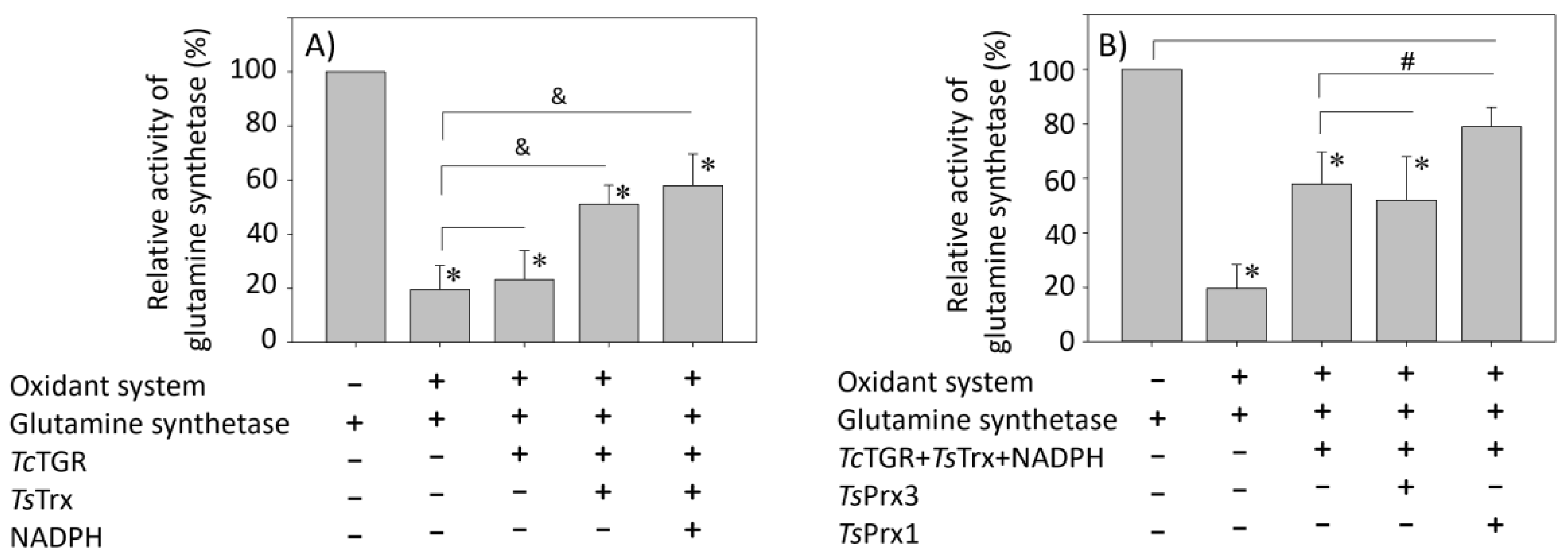
A Physiological Approach to Explore How Thioredoxin–Glutathione Reductase (TGR) and Peroxiredoxin (Prx) Eliminate H2O2 in Cysticerci of Taenia

Abstract
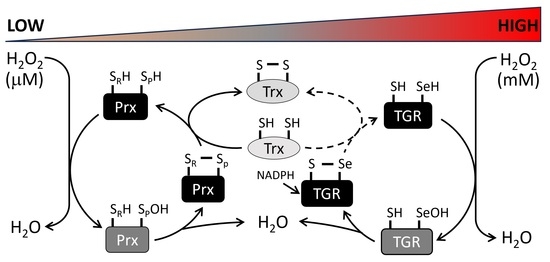
{kind=link}
{kind=link}
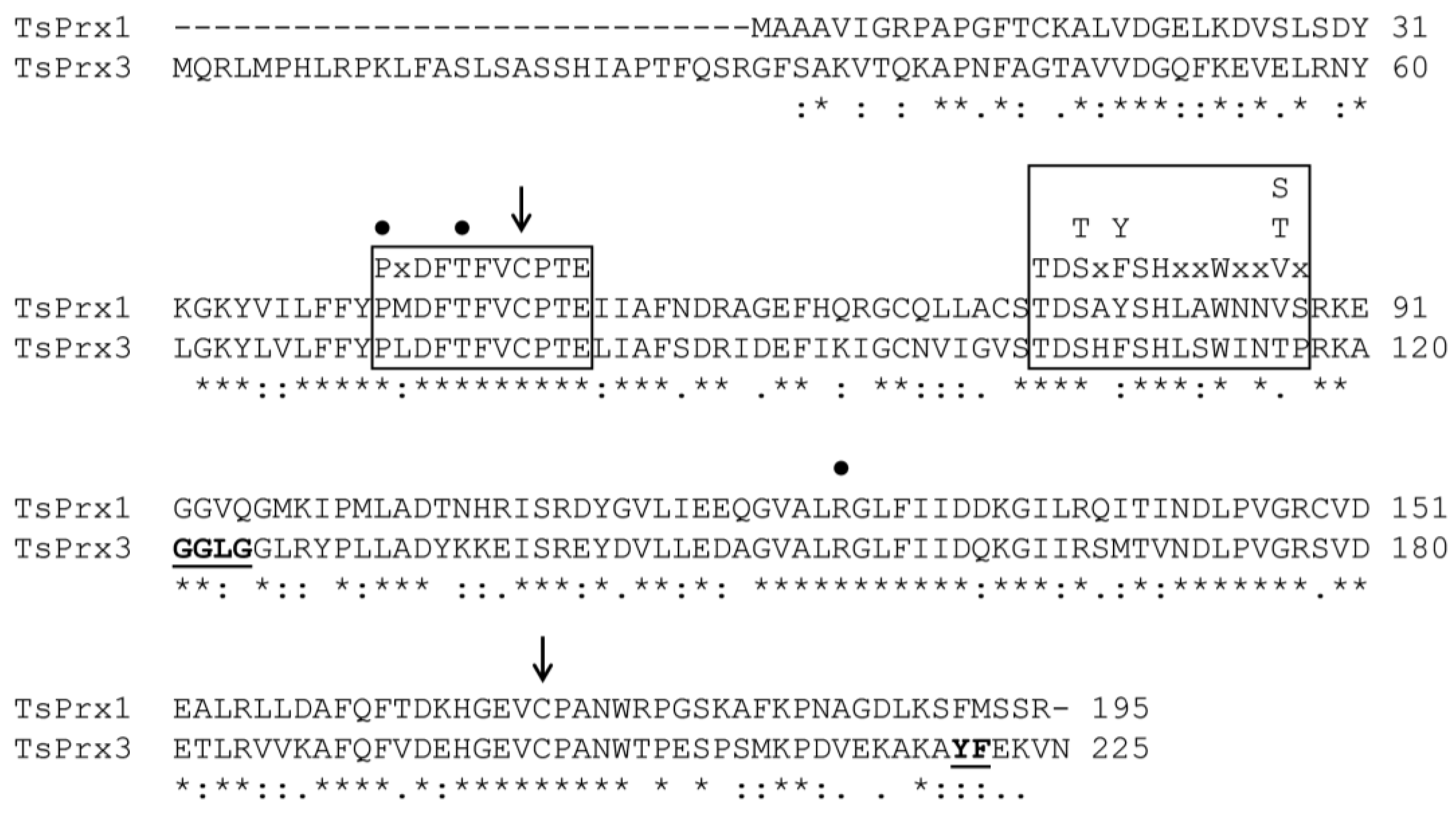
{kind=link}
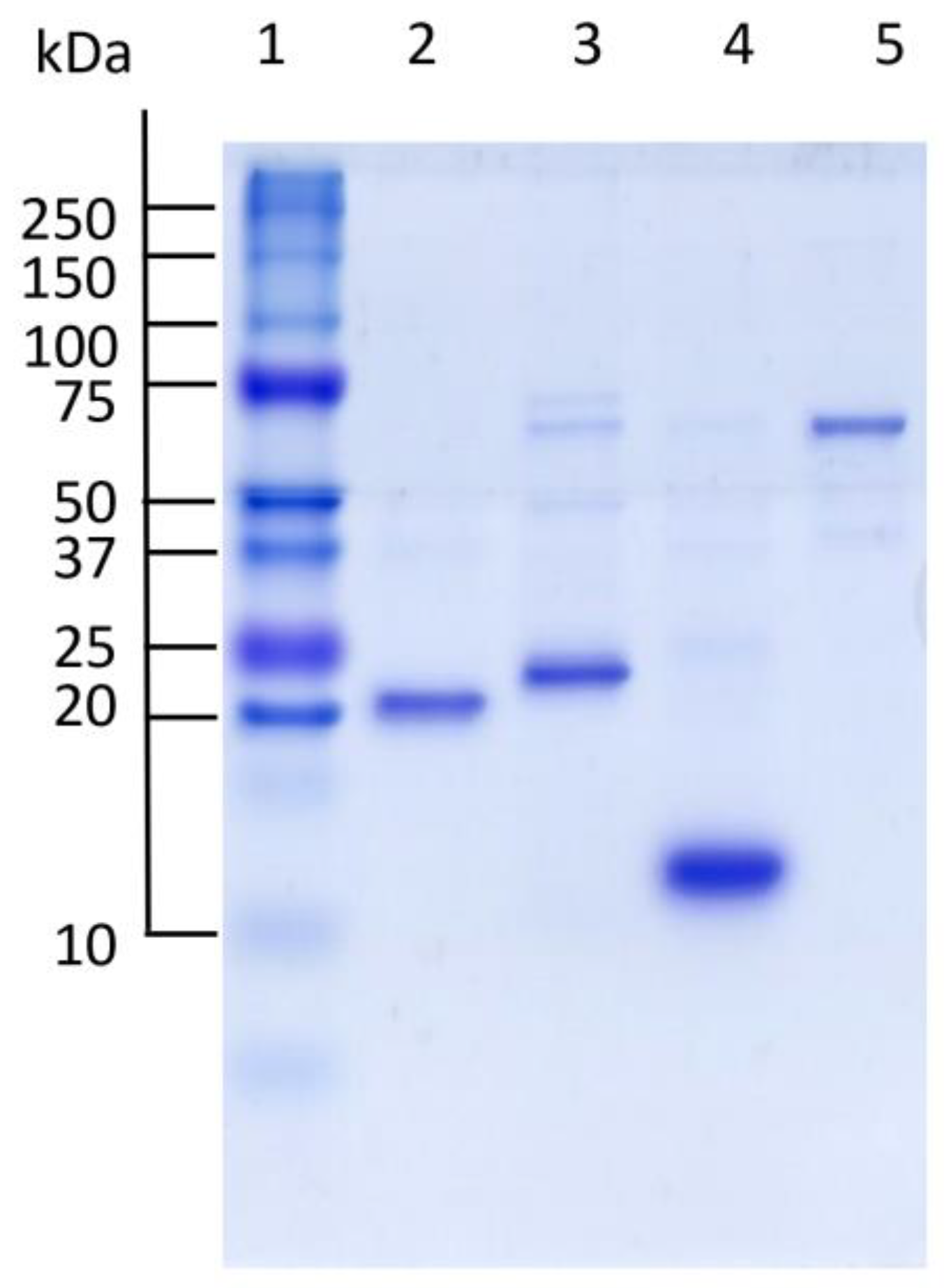
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Guevara-Flores, A.; Nava-Balderas, G.; de Jesús Martínez-González, J.; Vásquez-Lima, C.; Rendón, J.L.; del Arenal Mena, I.P. A Physiological Approach to Explore How Thioredoxin–Glutathione Reductase (TGR) and Peroxiredoxin (Prx) Eliminate H2O2 in Cysticerci of Taenia. Antioxidants 2024, 13, 444. https://doi.org/10.3390/antiox13040444
Guevara-Flores A, Nava-Balderas G, de Jesús Martínez-González J, Vásquez-Lima C, Rendón JL, del Arenal Mena IP. A Physiological Approach to Explore How Thioredoxin–Glutathione Reductase (TGR) and Peroxiredoxin (Prx) Eliminate H2O2 in Cysticerci of Taenia. Antioxidants. 2024; 13(4):444. https://doi.org/10.3390/antiox13040444
Chicago/Turabian StyleGuevara-Flores, Alberto, Gabriela Nava-Balderas, José de Jesús Martínez-González, César Vásquez-Lima, Juan Luis Rendón, and Irene Patricia del Arenal Mena. 2024. "A Physiological Approach to Explore How Thioredoxin–Glutathione Reductase (TGR) and Peroxiredoxin (Prx) Eliminate H2O2 in Cysticerci of Taenia" Antioxidants 13, no. 4: 444. https://doi.org/10.3390/antiox13040444
APA StyleGuevara-Flores, A., Nava-Balderas, G., de Jesús Martínez-González, J., Vásquez-Lima, C., Rendón, J. L., & del Arenal Mena, I. P. (2024). A Physiological Approach to Explore How Thioredoxin–Glutathione Reductase (TGR) and Peroxiredoxin (Prx) Eliminate H2O2 in Cysticerci of Taenia. Antioxidants, 13(4), 444. https://doi.org/10.3390/antiox13040444