

Synergistic Antioxidant and Anti-Inflammatory Effects of Phenolic Acid-Conjugated Glutamine–Histidine–Glycine–Valine (QHGV) Peptides Derived from Oysters (Crassostrea talienwhanensis)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Cell Culture
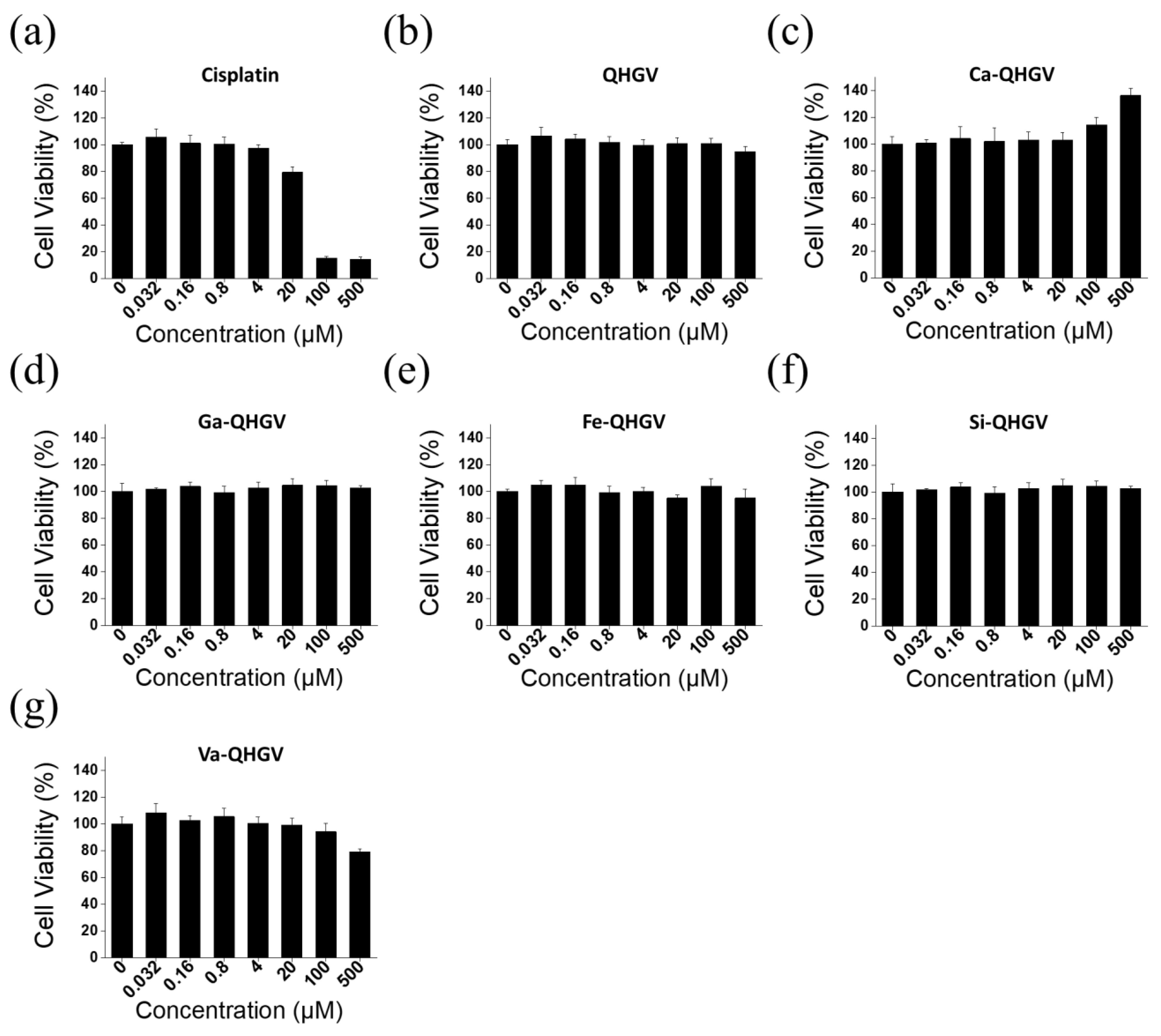
2.3. Cell Viability Assay
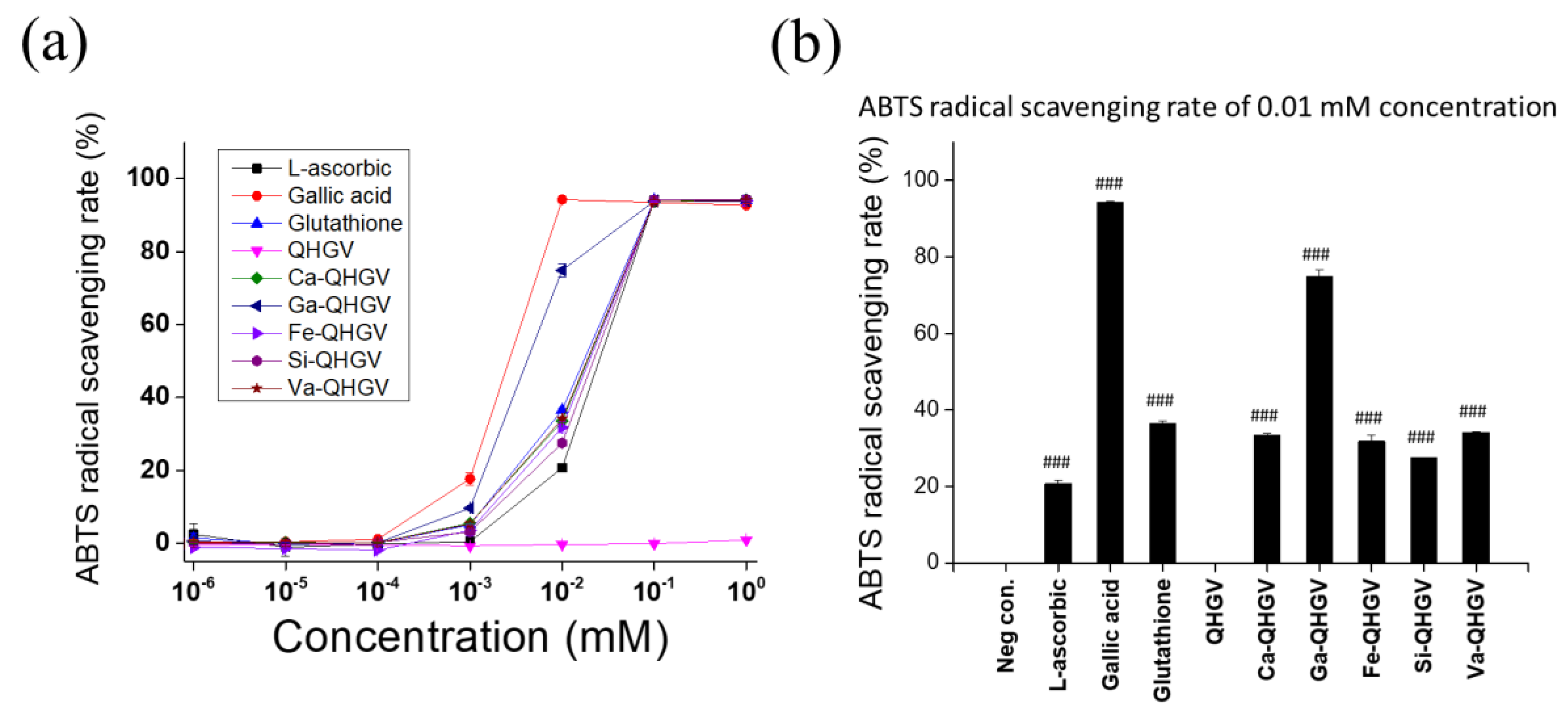
2.4. ABTS Radical Scavenging Activity Test
2.5. DPPH Radical Scavenging Activity Test
2.6. RNA Isolation and qRT-PCR
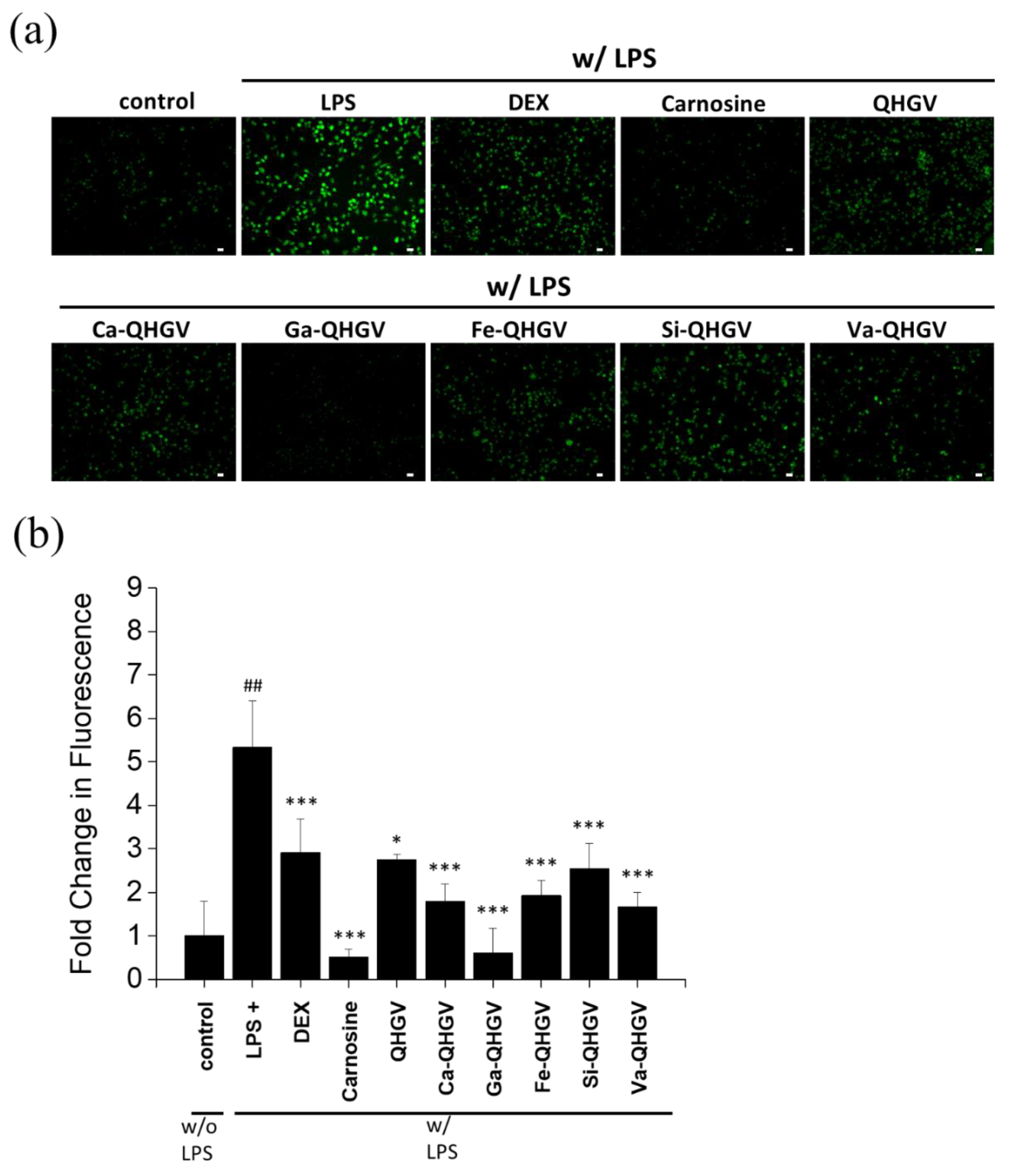
2.7. Quantification of ROS Using DCFH-DA Fluorescence
2.8. Western Blot Assay
2.9. Statistical Analysis
3. Results and Discussion
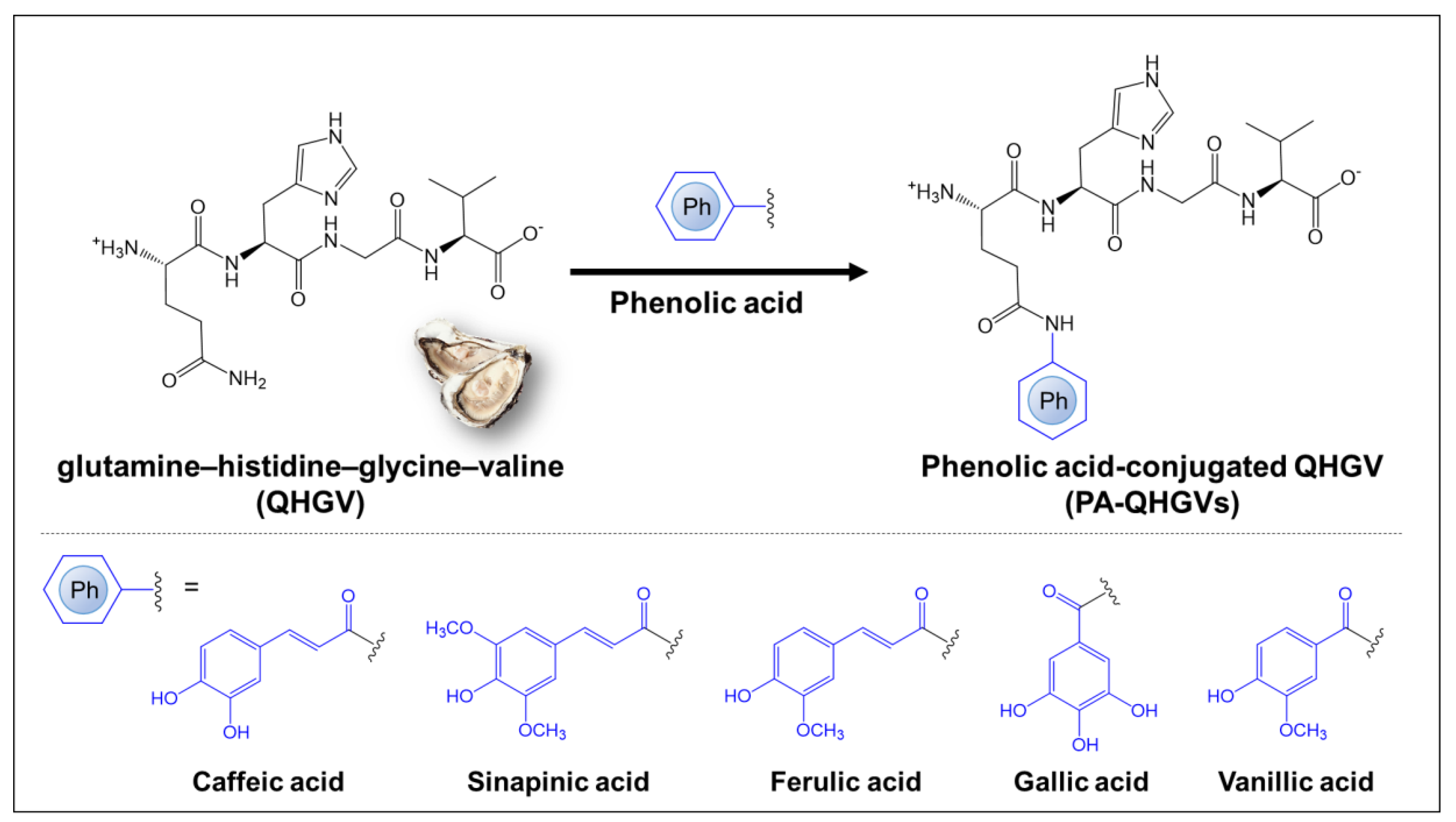
3.1. Chemical Structure of PA-QHGVs and Antioxidant Activity
3.2. Cytotoxicity of the PA-QHGVs in HaCaT Cells
3.3. PA-QHGVs Attenuates LPS-Induced ROS Generation in RAW 264.7 Cells
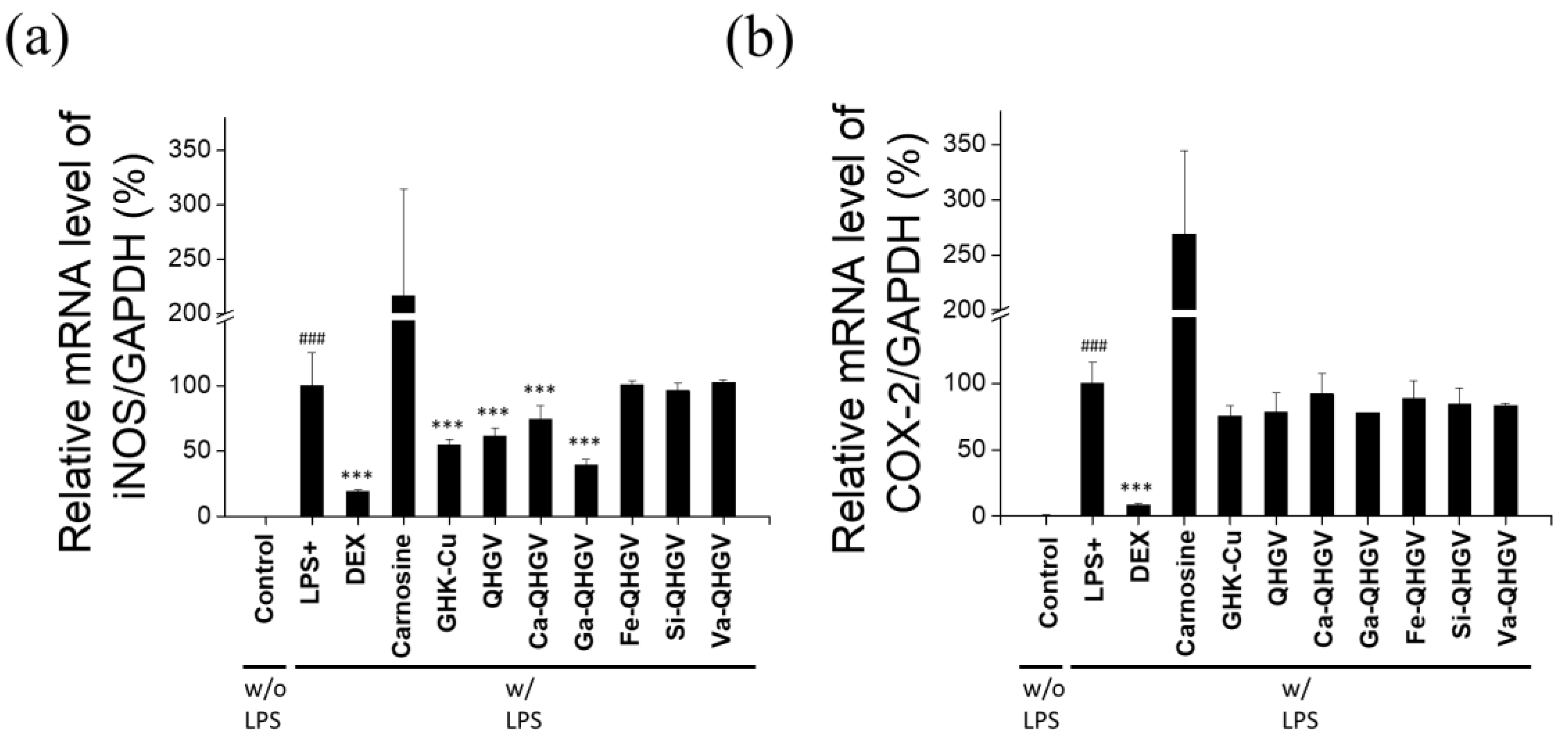
3.4. PA-QHGVs Inhibits iNos and Cox-2 mRNA Expression in LPS-Induced RAW 264.7 Cells
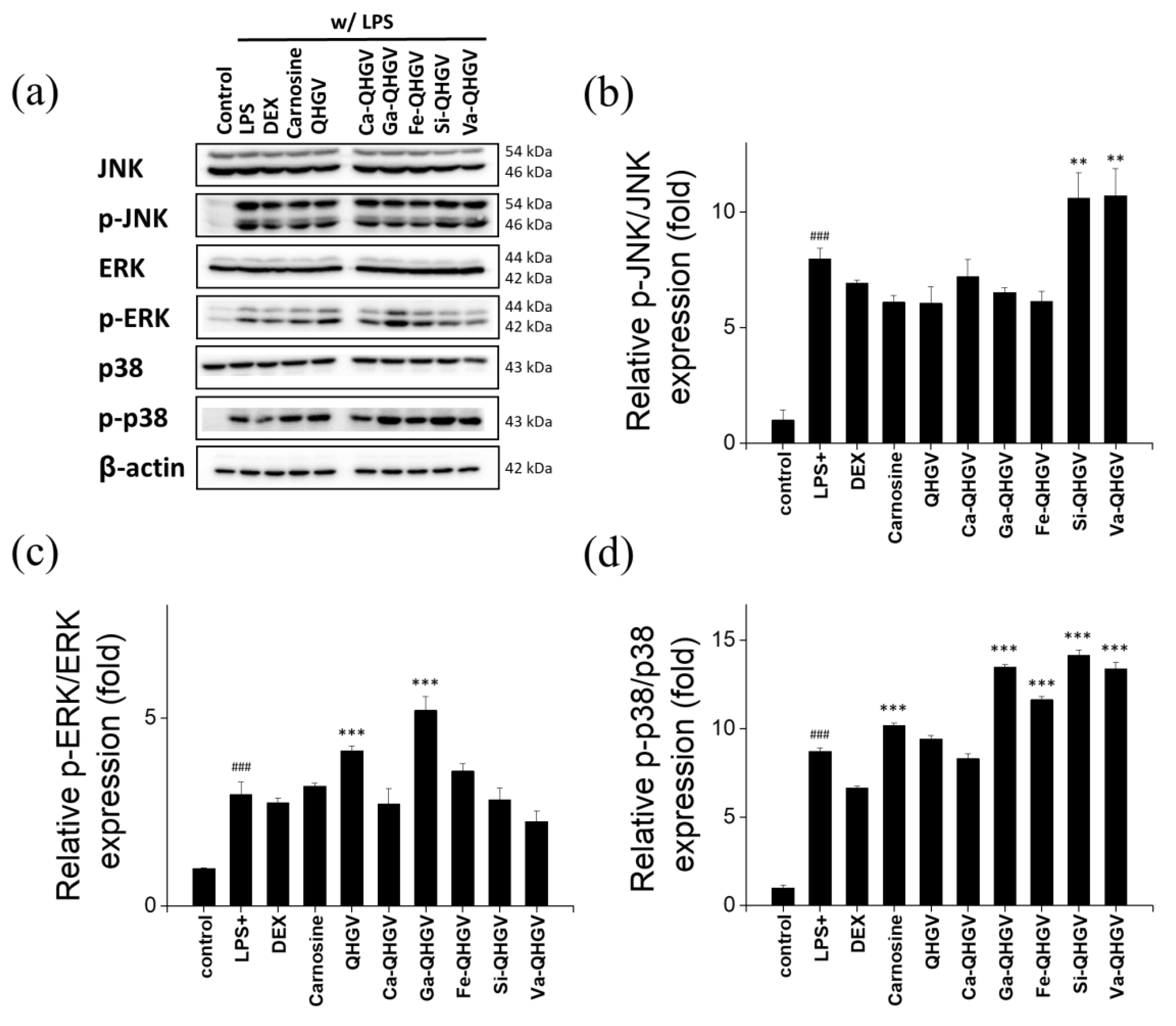
3.5. PA-QHGVs Antagonizes LPS-Induced Activation of the JNK/ERK/P38 Pathway
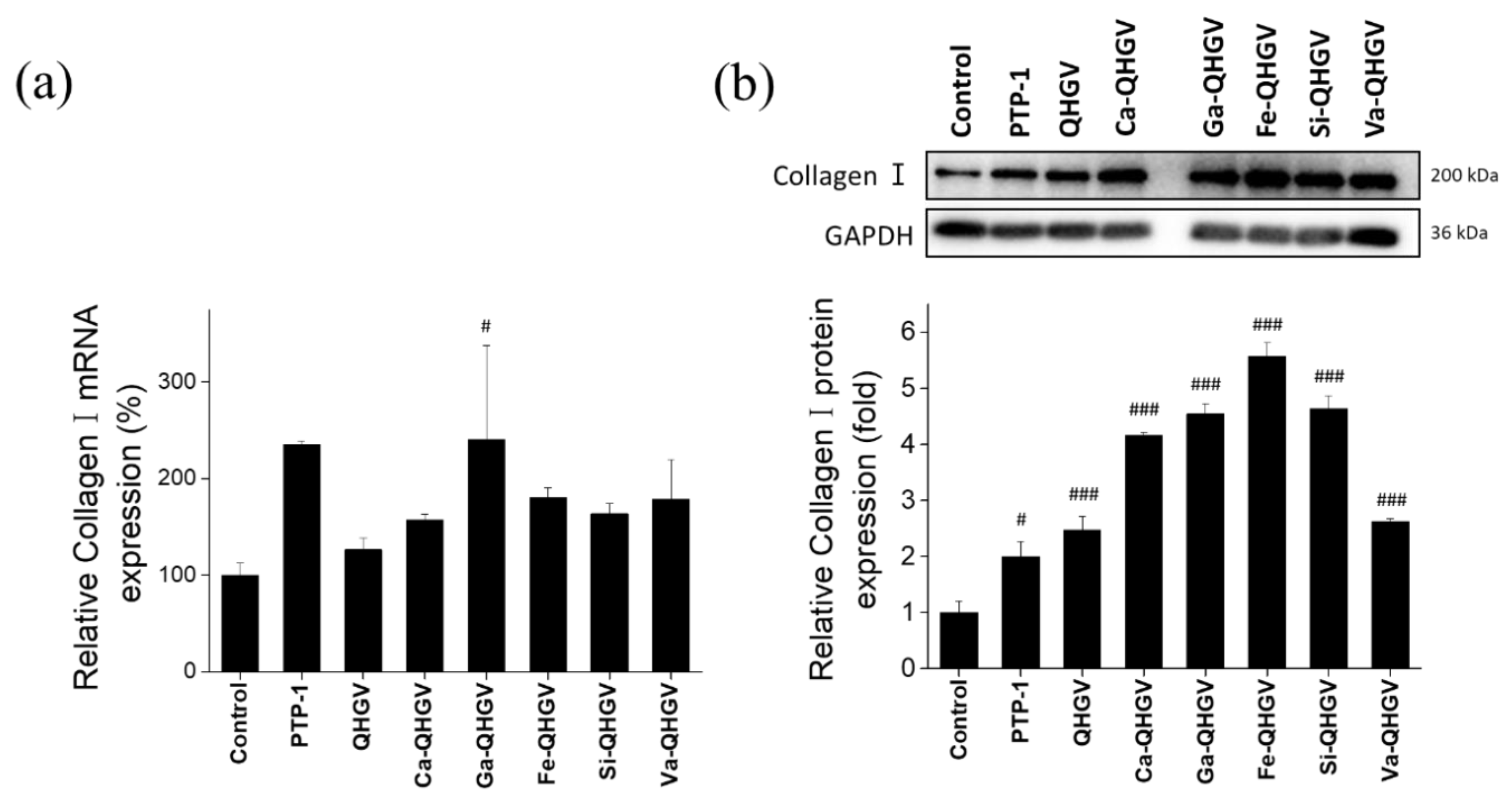
3.6. PA-QHGVs Upregulates the mRNA and Protein Levels of Collagen Type I in HaCaT Cells
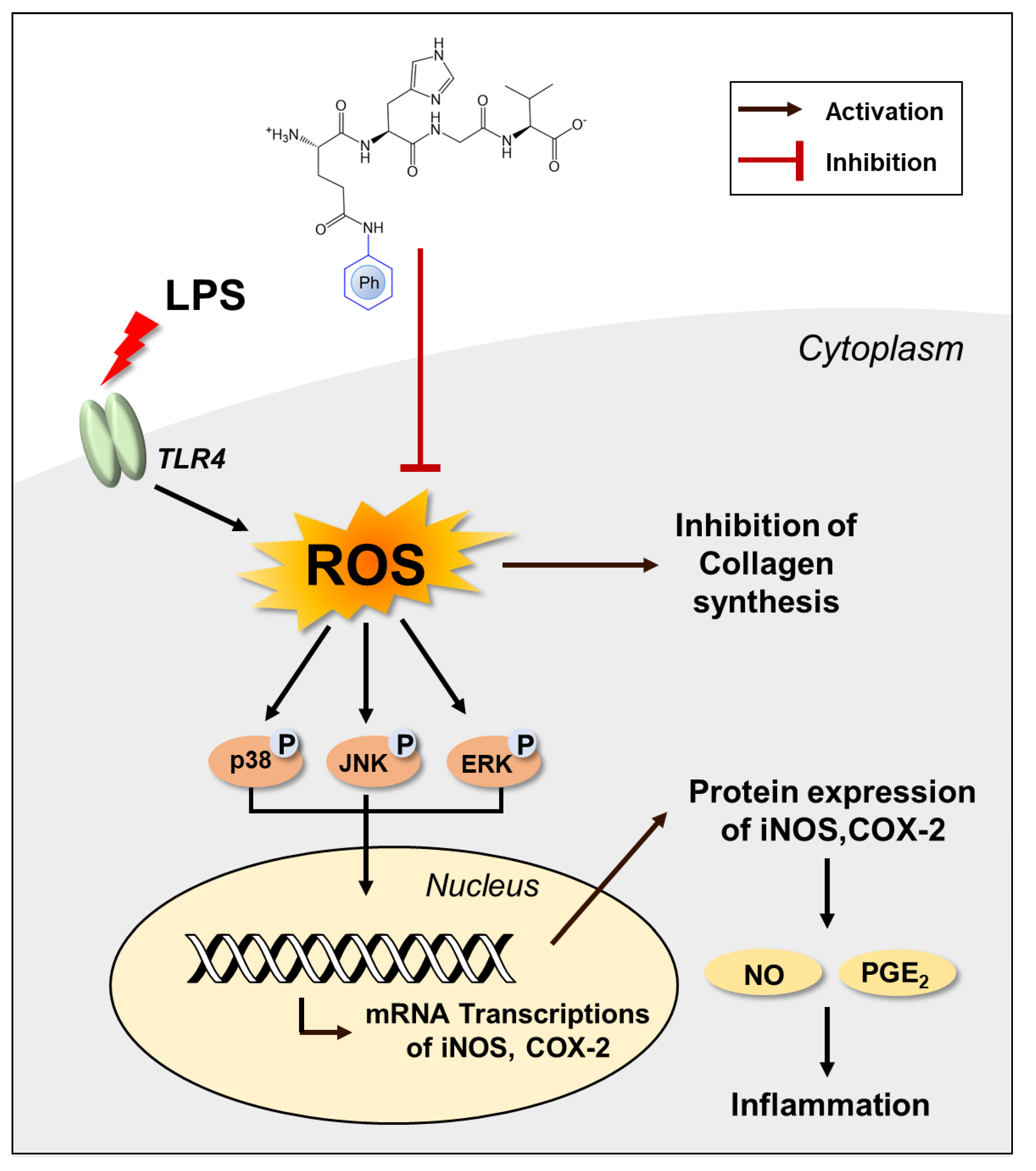
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakri, Z.F. Analyzing the Influence of Digital Marketing Strategies on Business Performance in the Beauty Industry: A Comprehensive Analysis of Social Media Engagement and Influencer Collaborations. J. Econ. Manag. Bus. Technol. 2023, 2, 37–48. [Google Scholar]
- Mortazavi, S.M.; Moghimi, H.R. Skin permeability, a dismissed necessity for anti-wrinkle peptide performance. Int. J. Cosmet. Sci. 2022, 44, 232–248. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, X.; Liu, D. Collagen peptides and the related synthetic peptides: A review on improving skin health. J. Funct. Foods 2021, 86, 104680. [Google Scholar] [CrossRef]
- Ngoc, L.T.N.; Moon, J.-Y.; Lee, Y.-C. Insights into Bioactive Peptides in Cosmetics. Cosmetics 2023, 10, 111. [Google Scholar] [CrossRef]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Yadav, A.R.; Mohite, S.K. Potential role of peptides for development of cosmeceutical skin products. Res. J. Top. Cosmet. Sci. 2020, 11, 77–82. [Google Scholar] [CrossRef]
- Rashid, J.; Sabar, M.F.; Gill, Z.; Mustafa, U.; Fatima, S.; Ashiq, S. Cosmeceuticals: The bioactive elements in new-age beauty products. Int. J. Pharm. Integr. Health Sci. 2023, 4, 70–82. [Google Scholar] [CrossRef]
- Yang, F.-J.; Xu, C.; Huang, M.-C.; Qian, Y.; Cai, X.-X.; Xuan, C.; Ming, D.; Huang, J.-I.; Wang, S.-Y. Molecular characteristics and structure–activity relationships of food-derived bioactive peptides. J. Integr. Agric. 2021, 20, 2313–2332. [Google Scholar] [CrossRef]
- Vogeler, S.; Wikfors, G.H.; Li, X.; Sauvage, J.; Joyce, A. Distribution of vitamin B12 in bivalve tissues: Investigations of larval and adult lifestages. Aquaculture 2022, 561, 738712. [Google Scholar] [CrossRef]
- Negara, B.F.S.P.; Mohibbullah, M.; Sohn, J.H.; Kim, J.S.; Choi, J.S. Nutritional value and potential bioactivities of Pacific oyster (Crassostrea gigas). Int. J. Food Sci. Technol. 2022, 57, 5732–5749. [Google Scholar] [CrossRef]
- Hao, L.; Wang, X.; Cao, Y.; Xu, J.; Xue, C. A comprehensive review of oyster peptides: Preparation, characterisation and bioactivities. Rev. Aquac. 2022, 14, 120–138. [Google Scholar] [CrossRef]
- Gao, J.-X.; Zhang, Y.-Y.; Huang, X.-H.; Liu, R.; Dong, X.-P.; Zhu, B.-W.; Qin, L. Comparison of amino acid, 5′-nucleotide and lipid metabolism of oysters (Crassostrea gigas Thunberg) captured in different seasons. Food Res. Int. 2021, 147, 110560. [Google Scholar] [CrossRef]
- Qian, B.; Zhao, X.; Yang, Y.; Tian, C. Antioxidant and anti-inflammatory peptide fraction from oyster soft tissue by enzymatic hydrolysis. Food Sci. Nutr. 2020, 8, 3947–3956. [Google Scholar] [CrossRef]
- Feng, C.; Tian, L.; Jiao, Y.; Tan, Y.; Liu, C.; Luo, Y.; Hong, H. The effect of steam cooking on the proteolysis of pacific oyster (Crassostrea gigas) proteins: Digestibility, allergenicity, and bioactivity. Food Chem. 2022, 379, 132160. [Google Scholar] [CrossRef]
- Peng, Z.; Gao, J.; Su, W.; Cao, W.; Zhu, G.; Qin, X.; Zhang, C.; Qi, Y. Purification and identification of peptides from oyster (Crassostrea hongkongensis) protein enzymatic hydrolysates and their anti-skin photoaging effects on UVB-irradiated HaCaT cells. Mar. Drugs 2022, 20, 749. [Google Scholar] [CrossRef]
- Wang, Q.; Li, W.; He, Y.; Ren, D.; Kow, F.; Song, L.; Yu, X. Novel antioxidative peptides from the protein hydrolysate of oysters (Crassostrea talienwhanensis). Food Chem. 2014, 145, 991–996. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Stuper-Szablewska, K. Phenolic acids used in the cosmetics industry as natural antioxidants. Eur. J. Med. Technol. 2019, 4, 24–32. [Google Scholar]
- Nakai, K.; Tsuruta, D. What are reactive oxygen species, free radicals, and oxidative stress in skin diseases? Int. J. Mol. Sci. 2021, 22, 10799. [Google Scholar] [CrossRef]
- Fang, Y.; Yang, L.; He, J. Plantanone C attenuates LPS-stimulated inflammation by inhibiting NF-κB/iNos/COX-2/MAPKs/Akt pathways in RAW 264.7 macrophages. Biomed. Pharmacother. 2021, 143, 112104. [Google Scholar] [CrossRef]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shahzad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef]
- Lu, Z.; Nie, G.; Belton, P.S.; Tang, H.; Zhao, B. Structure–activity relationship analysis of antioxidant ability and neuroprotective effect of gallic acid derivatives. Neurochem. Int. 2006, 48, 263–274. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A. Reactivity of phenolic compounds towards free radicals under in vitro conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Gil-Chávez, J.; Sotelo-Mundo, R.R.; Namiesnik, J.; Gorinstein, S.; González-Aguilar, G.A. Antioxidant interactions between major phenolic compounds found in ‘Ataulfo’mango pulp: Chlorogenic, gallic, protocatechuic and vanillic acids. Molecules 2012, 17, 12657–12664. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, M.; Xiao, C.; Zhao, Q.; Su, G. Practical problems when using ABTS assay to assess the radical-scavenging activity of peptides: Importance of controlling reaction pH and time. Food Chem. 2016, 192, 288–294. [Google Scholar] [CrossRef]
- Kostić, S.; Vilotić, A.; Pirković, A.; Dekanski, D.; Borozan, S.; Nacka-Aleksić, M.; Vrzić-Petronijević, S.; Krivokuća, M.J. Caffeic acid protects human trophoblast HTR-8/SVneo cells from H2O2-induced oxidative stress and genotoxicity. Food Chem. Toxicol. 2022, 163, 112993. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2022, 36, e22942. [Google Scholar] [CrossRef]
- Kim, Y.E.; Kim, J. ROS-scavenging therapeutic hydrogels for modulation of the inflammatory response. ACS Appl. Mater. Interfaces 2021, 14, 23002–23021. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, Z.; Zhou, X.; Zhao, Z.; Zhao, R.; Xu, X.; Kong, X.; Ren, J.; Yao, X.; Wen, Q. Microglia and macrophage exhibit attenuated inflammatory response and ferroptosis resistance after RSL3 stimulation via increasing Nrf2 expression. J. Neuroinflammation 2021, 18, 1–15. [Google Scholar] [CrossRef]
- Chen, Q.; Che, C.; Yang, S.; Ding, P.; Si, M.; Yang, G. Anti-inflammatory effects of extracellular vesicles from Morchella on LPS-stimulated RAW264. 7 cells via the ROS-mediated p38 MAPK signaling pathway. Mol. Cell. Biochem. 2023, 478, 317–327. [Google Scholar] [CrossRef]
- Saadati, S.; Kabthymer, R.H.; Aldini, G.; Mousa, A.; Feehan, J.; de Courten, B. Effects of carnosine and histidine-containing dipeptides on biomarkers of inflammation and oxidative stress: A systematic review and meta-analysis. Nutr. Rev. 2023, 2023, nuad150. [Google Scholar] [CrossRef]
- Anavi, S.; Tirosh, O. iNos as a metabolic enzyme under stress conditions. Free Radic. Biol. Med. 2020, 146, 16–35. [Google Scholar] [CrossRef]
- Ahmad, N.; Ansari, M.Y.; Haqqi, T.M. Role of iNos in osteoarthritis: Pathological and therapeutic aspects. J. Cell. Physiol. 2020, 235, 6366–6376. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Singh, P. Inflammation: Biochemistry, cellular targets, anti-inflammatory agents and challenges with special emphasis on cyclooxygenase-2. Bioorganic Chem. 2022, 121, 105663. [Google Scholar] [CrossRef]
- Zhang, Q.; Yan, L.; Lu, J.; Zhou, X. Glycyl-L-histidyl-L-lysine-Cu2+ attenuates cigarette smoke-induced pulmonary emphysema and inflammation by reducing oxidative stress pathway. Front. Mol. Biosci. 2022, 9, 925700. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.-h.; Li, M.; Ma, H.-f.; Li, W.; Liu, L.; Yin, Y.; Zhou, X.-m.; Hou, G. Protective effects of GHK-Cu in bleomycin-induced pulmonary fibrosis via anti-oxidative stress and anti-inflammation pathways. Life Sci. 2020, 241, 117139. [Google Scholar] [CrossRef] [PubMed]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G. Modulation of pro-oxidant and pro-inflammatory activities of m1 macrophages by the natural dipeptide carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef]
- Haftcheshmeh, S.M.; Abedi, M.; Mashayekhi, K.; Mousavi, M.J.; Navashenaq, J.G.; Mohammadi, A.; Momtazi-Borojeni, A.A. Berberine as a natural modulator of inflammatory signaling pathways in the immune system: Focus on NF-κB, JAK/STAT, and MAPK signaling pathways. Phytother. Res. 2022, 36, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Xiao, Z.; Tong, H.; Liu, Y.; Wu, Y.; Ge, C. Oral intake of chicken bone collagen peptides anti-skin aging in mice by regulating collagen degradation and synthesis, inhibiting inflammation and activating lysosomes. Nutrients 2022, 14, 1622. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qiu, H.; Xu, Y.; Gao, Y.; Tan, P.; Zhao, R.; Liu, Z.; Tang, Y.; Zhu, X.; Bao, C. The biological effect of recombinant humanized collagen on damaged skin induced by UV-photoaging: An in vivo study. Bioact. Mater. 2022, 11, 154–165. [Google Scholar] [CrossRef]
- Rizzi, V.; Gubitosa, J.; Fini, P.; Cosma, P. Neurocosmetics in skincare—The fascinating world of skin–brain connection: A review to explore ingredients, commercial products for skin aging, and cosmetic regulation. Cosmetics 2021, 8, 66. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Direction | Nucleotide Sequence | Gene Accession Number |
|---|---|---|---|
| iNos | Forward | 5′-GCG CCT TTG CTC ATG ACA TT-3′ | NM_000625.4 |
| Backward | 5′-ACA GGC TGC CTT GAA GGT TT-3′ | ||
| Cox-2 | Forward | 5′-AAG TTC ATC CCT GAT CCC CA-3′ | NM_000963.4 |
| Backward | 5′-GCC CTC GCT TAT GAT CTG TC-3′ | ||
| Col1a1 | Forward | 5′-CTG ACC TCC TGC GCC TGA TGT CC-3′ | NM_000088.4 |
| Backward | 5′-GTC TGG GGC ACC AAC GTC CAA GGG-3′ | ||
| Gapdh | Forward | 5′-GAA GGT CGG AGT CAA CGG AT-3′ | NM_001289745.3 |
| Backward | 5′-TGG AAT TTG CCA TGG GTG GA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.; Han, S.; Lee, S.; Kim, J.; Kim, J.; Kang, D.-K. Synergistic Antioxidant and Anti-Inflammatory Effects of Phenolic Acid-Conjugated Glutamine–Histidine–Glycine–Valine (QHGV) Peptides Derived from Oysters (Crassostrea talienwhanensis). Antioxidants 2024, 13, 447. https://doi.org/10.3390/antiox13040447
Choi S, Han S, Lee S, Kim J, Kim J, Kang D-K. Synergistic Antioxidant and Anti-Inflammatory Effects of Phenolic Acid-Conjugated Glutamine–Histidine–Glycine–Valine (QHGV) Peptides Derived from Oysters (Crassostrea talienwhanensis). Antioxidants. 2024; 13(4):447. https://doi.org/10.3390/antiox13040447
Chicago/Turabian StyleChoi, Soyun, Sohee Han, Seungmi Lee, Jongmin Kim, Jinho Kim, and Dong-Ku Kang. 2024. "Synergistic Antioxidant and Anti-Inflammatory Effects of Phenolic Acid-Conjugated Glutamine–Histidine–Glycine–Valine (QHGV) Peptides Derived from Oysters (Crassostrea talienwhanensis)" Antioxidants 13, no. 4: 447. https://doi.org/10.3390/antiox13040447